


Abstract
Eleven derivatives from Antrodia camphorata were isolated in order to evaluate their selective cytotoxicity toward 14 types of human cancer cell and two non-transformed cell types. Among these triterpenoids, methyl antcinate A (MAA) exhibited the most potent spectrum of anticancer effects in KB cells, four different oral cancer cell lines (TSCCa, GNM, OC-2, and OEC-M1), Panc-1, BT474, PC-3, OVCAR-3, HeLa, and U2OS cells with high selectivity indices (CC50/IC50). The expression of B-cell lymphoma 2 (Bcl-2), Bcl-2–associated X protein (Bax), and poly(ADP-ribose) polymerase (PARP) of PC-3 cells tested by western blotting suggested that MAA exerts cell death through the caspase-dependent cascade and the Bax-mediated mitochondrial apoptotic pathway, not only on liver and oral cancer cells but on other types as well, including prostate cancer, in a dose-dependent manner. In addition to MAA, methyl antcinate B, dehydroeburicoic acid, and 15α-acetyl-dehydrosulfurenic acid also exhibited significant selective cytotoxic effects to respective cancer cells. Modifications of these triterpenoids may lead to the development of more potent anticancer drugs.
- Antrodia camphorata
- anticancer effects
- apoptosis
- caspase
- cytotoxicity
- triterpenoid
- PC-3
- TsCCA
- AsPC
- BT474
- T47D
- HeLa
- U2OS cells
- Bcl-2
- Bax
- PARP
Antrodia camphorata (A. camphorata), also called A. cinnamomea, is a parasitic fungus that only grows on the inner heartwood wall of Cinnamomum kanehirai Hay (Lauraceae). A. camphorata has been used in traditional Chinese medicine over centuries for food, wine, and drug intoxication, as well as hypertension, itching, and the improvement of liver and stomach immunity (1). Crude extracts of A. camphorata exhibited a broad range of biological activities, and recent research has focused on elucidating the pharmacological potential including anticancer, anti-inflammatory/immunomodulatory, and hepatoprotective effects (2-7). Our previous study also reported that the chloroform extract of fruiting bodies of A. camphorata exhibited cytotoxic activity towards human cancer cell lines Jurkat, HepG2, Colo 205, and MCF-7 (8). However, previous reports on the pharmacological activities of A. camphorata mainly utilize the crude extracts, raising the concern as to which compounds within the crude extract are responsible for generating the observed anticancer effects. Moreover, the variation in the purity of extract preparation may cause a discrepancy in the observed effects.
The characteristic constituents of A. camphorata are various benzenoids, terpenoids, benzoquinones, lignans, and maleic/succinic acid derivatives aside from polysaccharides. Among these constituents, triterpenoids are the major representative of phytoconstituents in the fruiting bodies of A. camphorata (9). Previously, we have isolated five lanostane- and three ergostane-type triterpenes from the fruiting bodies, and three of these compounds, antcin A, antcin C, and methyl antcinate A (MAA), can induce apoptosis on human colon cancer cells HT-29 and SW-480 (10). Furthermore, we demonstrated that MAA induces apoptosis in human liver and oral cancer cells through the mitochondrial-dependent pathway (11, 12).
Herein we report the extensive screening results of the anticancer effects of 11 triterpenoids isolated from the fruiting bodies of A. camphorata. Four series of 11 derivatives were isolated and used to evaluate selective cytotoxicity towards 14 human cancer cell lines and two human non-transformed cell types in the present study.
Materials and Methods
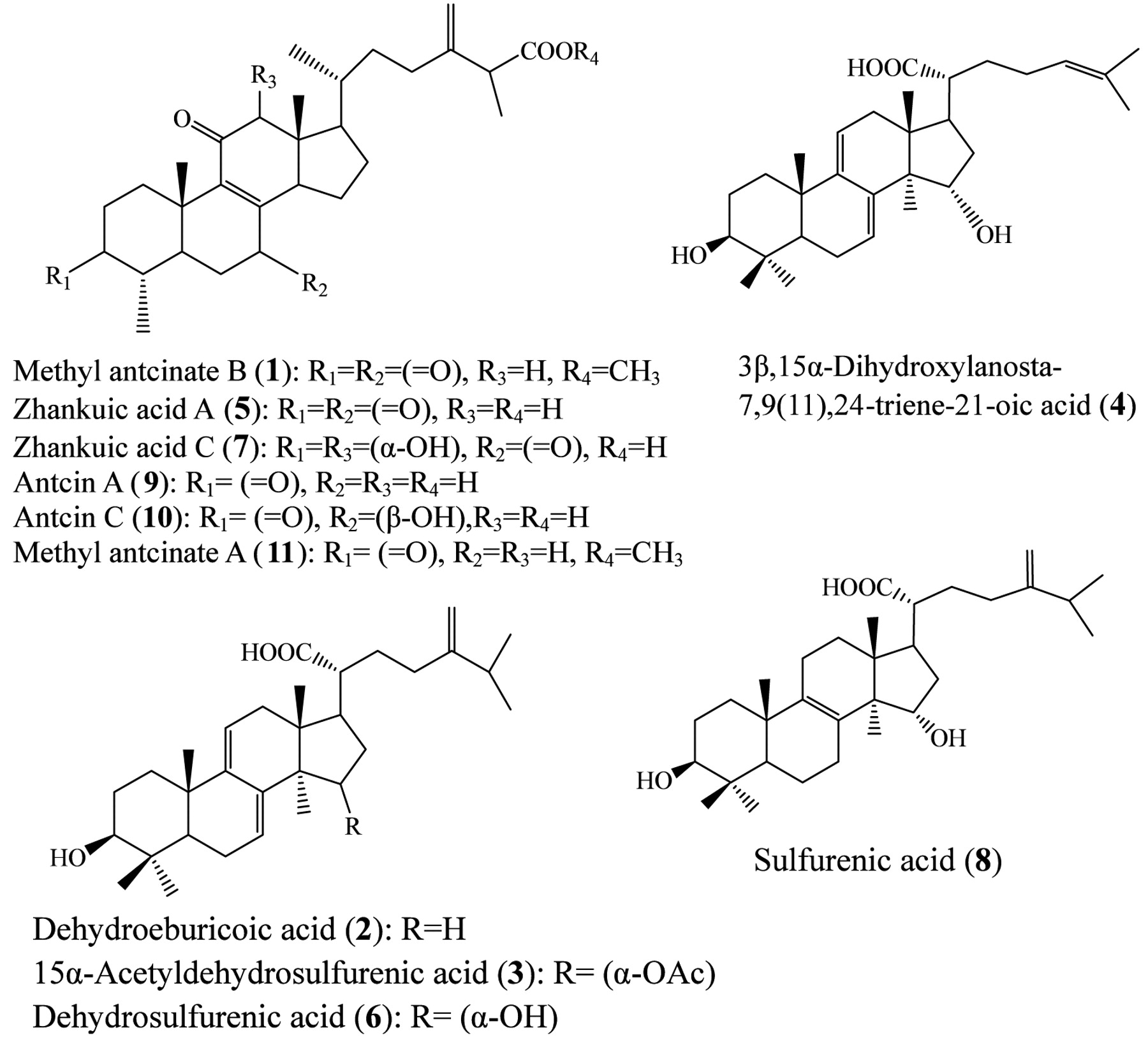
Extraction and isolation of compounds from fruiting bodies of A. camphorate. Fruiting bodies of A. camphorata were collected from Yuli, Hualien County, Taiwan, in December 2007. The species was identified by Professor Yew-Min Tzeng at the Natural Products and Bioprocess Laboratory, Chaoyang University of Technology. A voucher specimen (YMT 7002) was deposited in the Herbarium of the Institute of Biochemical Sciences and Technology, Chaoyang University of Technology, Taiwan, ROC. Air-dried powder of the fruiting bodies (101.9 g) of A. camphorata was successively extracted (3×1000 ml) with n-hexane, chloroform and methanol under reflux. The resulting extracts were concentrated individually under reduced pressure to yield 3.02 g (2.96% based on dry weight), 46.3 g (45.43%) and 2.7 g (2.64 %) of residues from n-hexane, chloroform and methanol (MeOH) extracts, respectively. Repeated silica-gel column chromatography (5×90 cm) of the chloroform extract residue was carried out with increasing polarity of elutant using mixtures of hexane/ethyl acetate. Following thin-layer chromatographic analysis, eluates with similar profiles were combined to produce six fractions (F1-F6). Fraction F2 was rechromatographed over a silica-gel column eluted with a gradient of hexane/EtOAc to afford methyl antcinate B (MAB, compound 1), dehydroeburicoic acid (DeEA, compound 2) and 15α-acetyl dehydrosulfurenic acid (ADSA, compound 3), respectively. Fraction F3 was purified by silica-gel chromatography eluted with a gradient of hexane/ethyl acetate, resulting in 3β,15α-dihydroxylanosta-7,9,24-triene-21-oic acid (compound 4), and zhankuic acid A (compound 5). Fractions F4 and F5 were purified by column chromatography and eluted with a gradient of n-hexane/ethyl acetate, to afford four subfractions (D1 to D4). Dehydrosulfurenic acid (compound 6) and zhankuic acid C (compound 7), and sulfurenic acid (SA, compound 8) were obtained from subfractions D2, D3 and D4, respectively. Fraction F6 was purified by silica-gel column chromatography using chloroform/methanol from 100% chloroform to 20% methanol to yield antcin A (Ant A, compound 9) and a mixture of compounds that was further separated by column chromatography using chloroform/methanol elution to give antcin C (Ant C, compound 10) and methyl antcinate A (MAA, compound 11). The structures of compounds 1-11 were determined by 1H- and 13C-nuclear magnetic resonance (NMR) spectroscopy and by comparison of the spectral data with published values (13, 14).
Cell culture. Cells used in this study were purchased from Bioresource Collection and Research Center, Food Industry Research and Development Institute (Hsinchu, Taiwan, ROC) or provided by the Cell Bank of Chung Shan Medical University. TsCCA, AsPC, BT474, T47D, HeLa, U2OS and gingival fibroblast (GF) cells were grown in Dulbecco's modified Eagle's medium (DMEM; Life Technologies, Carlsbad, CA, USA) supplemented with 10% fetal bovine serum (FBS; Life Technologies). GNM, OC-2, OEC-M1, Panc-1, PC-3 and AGS cells were cultured in RPMI-1640 with 10% FBS. KB cells were maintained in Eagle's minimal essential medium (MEM) with 10% FBS. OVCAE-3 cells were cultured in DMEM with 20% FBS and 0.01 mg/ml bovine insulin. The maintenance growth medium for the MCF-12A cells consisted of a 1:1 mixture of DMEM and Ham's F12 medium, 20 ng/ml epidermal growth factor, 100 ng/ml cholera toxin, 10 mg/ml insulin and 500 ng/ml hydrocortisone. All media included penicillin (100 IU/ml) and streptomycin (100 μg/ml). Briefly, the cells were maintained in the appropriate growth medium at 37°C in a humidified atmosphere of 5% CO2 and 95% air and were used over a restricted culture period of 10 passages.
Cell viability analysis. The effect of test agents on cell viability was assessed by using the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazolium bromide (MTT) assay in 4 to 6 replicates as previously described (15-19). All tested cell lines were grown in 5% FBS-supplemented culture medium in 96-well plates overnight, and then exposed to the different compounds for 24 hours at different concentrations, including 10, 50 and 200 μM. Controls received 0.1% (V/V) dimethyl sulfoxide (DMSO) vehicle at a concentration equal to the one used for drug-treated cells. At the end of the treatment, the medium was removed, replaced by 200 μl of 0.5 mg/ml of MTT in the same medium, and cells were re-incubated for 2 h. Supernatants were removed from the wells, and the reduced MTT dye was solubilized in 200 μl/well DMSO. Absorbance was determined at 595 nm on a plate reader.
Caspase-3 activity. Caspase-3 activity was determined using the BD ApoAlert Caspase-3 assay according to the manufacturer's instructions (Becton, Dickinson and Company, Palo Alto, CA, USA) as previously described (16, 19, 20). Briefly, 2×106 cells were centrifuged at 400 ×g for 5 min, resuspended in 50 μl of chilled cell lysis buffer and incubated on ice for 10 min, then 50 μl of 2× reaction buffer/ DTT mix and 1 μl of caspase-3 inhibitor (DEVD-fmk, 1 mM) were added to some samples which were then incubated on ice for 30 min as negative controls; 50 μl of 2× reaction buffer/DTT mixture and 1 μl DMSO per 50 μl of 2× reaction buffer were added to the samples without inhibitor to ensure that all samples were tested under similar conditions. Five microliters of 1 mmol/l caspase-3 substrate DEVD-pNA were added and the mixture was incubated at 37°C for 1 h. Cells were analyzed by a FACScan flow cytometer (Becton, Dickinson and Company). All samples were run in triplicate with appropriate controls. Camptothecin (Sigma-Aldrich, St. Louis, MO, USA) served as the positive control.
Western blotting. Western blotting was performed as previously described (12, 19). Cells were lysed in boiled lysis buffer containing 1% SDS and 10 mM/l Tris-HCl (pH 7.4). Following centrifugation at 400 ×g for 10 min, the protein concentration in each lysate was measured (Bradford assay). Equal amounts of total protein from each sample were fractionated by SDS-PAGE, blotted onto a polyvinylidene difluoride membrane, and probed with the primary antibody, mouse monoclonal anti-caspases-3 (1:1000, v/v), or rabbit monoclonal antibodies to poly (ADP-ribose) polymerase (PARP) (1:4000, v/v), Bcl-2-associated × protein (Bax) (1:8000, v/v) and β-actin (1:2000, v/v), followed by 1 h incubation with a 1:10000 (v/v) dilution of the appropriate horseradish peroxidase (HRP)-conjugated secondary antibody. Primary antibodies (β-actin, PARP, Bax) were purchased from Cell Signaling Technology Inc. (Danvers, MA, USA), whereas secondary antibodies (anti-rabbit IgG and anti-mouse IgG HRP-labeled) were purchased from PerkinElmer (Boston, MA, USA). The hybridized immunocomplex was detected by Renaissance Chemiluminescence Reagent Plus (NEN Life Science Products, Boston, MA, USA).

Chemical structures of triterpenoid compounds 1-11 isolated from the fruiting bodies of Antrodia camphorata.
Statistical analysis. Values are the mean±standard deviation (SD) of experiments carried out in triplicate. Statistical significance was tested by Student's t-test for either paired or unpaired data, where appropriate. Statistical significance was defined as p<0.01.
Results
Anticancer cytotoxicity assays of compounds. Figure 1 shows the chemical structures of triterpenoid compounds 1-11 isolated from the fruiting bodies of A. camphorata. Fourteen cancer cell lines of head and neck, pancreas, breast, prostate, ovary, cervix, stomach and bone cancer were utilized to examine the 50% inhibitory concentration (IC50) by the MTT assay. Two non-transformed cells, MCF-12A and GF were also included as normal controls for testing 50% cytotoxic concentration (CC50). Table I summarizes the IC50 and CC50 data. Although all the triterpenoid compounds were isolated from fruiting bodies of A. camphorata, the antiproliferative effects were remarkably different. Among the eight cancer types tested, head and neck cancer and prostate cancer cell lines were especially sensitive to the isolated compounds. However, zhankuic acid A (compound 5), zhankuic acid C (compound 7), Ant A (compound 9), and Ant C (compound 10) hardly suppressed cancer cell growth, neither did 3β,15α-dihydroxylanosta-7,9(11),24-triene-21-oic acid (compound 4) nor SA (compound 8). On the contrary, methyl antcinate and DeEA derivatives showed significant inhibitory effects on most cancer types investigated, especially the head and neck cancer cells. MAA (compound 11) and MAB (compound 1) had significant cytotoxicity, with IC50 values for GNM cells of 12.6 and 35.8 μM, respectively (Table I). Figure 2 shows the morphological change of PC-3 cells after treating each compound with 50 μM for 24 hours. Fifty micromolar camptothecin served as the positive control and 0.1% (V/V) DMSO as the vehicle control. MAA (compound 11) and MAB (compound 1) caused a significant decrease in cell numbers, as well as a change in cell morphology. Among the eight types of cancers screened, MAA (compound 11), MAB (compound 1), DeEA (compound 2) and ADSA (compound 3) exhibited the most potent cytotoxic effects.

Morphological change in PC-3 cells induced by treatment with triterpenoid compounds 1-11. PC-3 cells were treated with different compounds for 24 hours. Cell morphology was observed under optical microscopy at ×100 (A) and ×200 (B) magnification. Ctrl: DMSO vehicle control; Camp.: camptothecin, positive control. For compound names, see legend to Figure 1.
Caspase-3 activity assays. The CC50 of MAA on MCF-12A and GF was 133.6 and 194.8 μM, respectively, while the IC50 of MAA on cancer cells was less than 50 μM (Table I). Among the 11 compounds, MAA demonstrated the most profound anticancer effects. MAA was thus used as the representative compound for a further apoptosis study. In order to examine the apoptotic death mode of cancer cells treated with the 11 compounds, caspase-3 activity assays were performed by flow cytometry. As GNM, OEC-M1, and OC-2 oral cancer cell lines have been investigated in our previous study (12), which suggested that MAA induces its growth inhibition via Bax-mediated mitochondrial apoptotic pathway, another cell line, PC-3 (prostate cancer), was used in this study. Figure 3 shows the results of the caspase-3 assay. Taken together with the results of the cytotoxic assays, the mode of cancer cell growth inhibition induced by these compounds treatment appeared to be caspase-3-mediated apoptosis (p<0.01).

Effects of triterpenoid compounds 1-11 on caspase-3 activity in PC-3 cells. PC-3 cells were treated with 50 μM of each compound or camptothecin (Camp.), which served as the positive control. Flow cytometric analysis was carried out after 24-h treatment in order to determine the caspase-3 activity. DMSO served as the vehicle control. For compound names, see legend to Figure 1.
Investigation of apoptotic pathway. To clarify whether the apoptotic pathway induced by MAA is the same in other types of cancer as it is in oral cancer, the protein expression of Bcl-2, Bax, and PARP in PC-3 prostate cancer cells treated with MAA were tested by western blotting. Treatment of PC-3 cells with different concentrations of MAA for 6 h promoted dose-dependent cleavage of PARP from the full-length 116-kDa to an inactive 85-kDa form by activated caspases (Figure 4), which is another indicator of apoptosis. Moreover, the results of the western blot analysis showed that the expression of pro-apoptotic Bax protein increased and that of anti-apoptotic Bcl-2 protein decreased in a dose-dependent manner (p<0.01) (Figure 4). This finding suggests that the effect of the Bax gene product via mitochondria might be responsible for MAA-induced apoptosis in PC-3 cells. The results suggest that MAA triggers the Bax-mediated mitochondrial apoptotic pathway in PC-3 prostate cancer cells in a dose-dependent manner, as it does in oral cancer cells as reported in our previous study (12).
Discussion
These results suggest that the anticancer capacities of the 11 triterpenoids are variable for the eight types of cancer cells screened. MAB, DeEA, and ADSA, all exhibited significant anticancer effects. MAB has been shown to induce apoptosis in HT-29 and SW-480 cells, especially in combination with zhanhuic acids A (compound 5) and C (compound 7) (10). However, the latter used alone, were not as potent as in colon and breast cancer according to our previous study (10). The inhibition of reactive oxygen species (ROS) production in leukocytes by eburicoic acid, DeEA, SA, and Ant C has been suggested to be responsible for the immunomodulating effects of A. camphorata (21). DeEA exhibited non-cytotoxic effects towards human stomach cancer NUGC-3 cells (22, 23); here it had significant cytotoxic effects on various cancer types, especially on head and neck cancer. Moreover, it has been reported that DeEA induced calcium- and calpain-dependent necrosis but not apoptosis in human U87MG glioblastomas (24); this might explain its slight cytotoxicity toward non-transformed cells, MCF-12A and GF, and the broad spectrum of anticancer effects apparent in the present study.
The cell growth inhibitory effects of the 11 compounds towards 14 human cancer cell lines and two non-transformed cells. The concentrations of each tested compound required to inhibit cancer cell proliferation by 50% (IC50) and non-transformed cell proliferation by 50% (CC50) after 24 h of treatment are shown.
Zhanhuic acids A and C were first isolated as new steroid acids from A. cunnamomea in 1995 (25). Zhankuic acid A also exhibited significant cytotoxicity towards leukemia HL 60 cells (IC50=5.45 μg/ml). It also demonstrated potential in combined therapy with histone deacetylase inhibitor trichostatin A (TSA) by increasing NFκB activation and death receptor 5 expression, leading to an increased cytotoxic sensitivity towards TSA (7). However, it had no significant growth inhibition effect on various cancer cells in the present study. This result is consistent with the finding observed by Yu et al, that zhanhuic acid A and C showed no significant cytotoxicity at 50 μM in bone marrow-derived dendritic cells (26). Ant A exerted its anti-inflammatory action by inhibiting superoxide generation, since it could act against fMLP-induced production of superoxide anion by neutrophils, with an IC50 value of 8.55±1.04 μM (23). In addition to zhanhuic acid A and C, DeEA, 15α-acetyl-dehydrosulfurenic acid, SA, dehydrosulfurenic acid, Ant A, and Ant C have been examined. After 16 hours' treatment, none of these compounds had any significant cytotoxicity at 50 μM (26). Although zhanhuic acids A and C had no appreciable anticancer effects, pretreatment with these compounds dose-dependently diminished fMLP- and PMA-induced ROS production (27). Zhankuic acids A and C also effectively inhibited fMLP- or PMA-induced firm adhesion without interfering with the expression of surface Mac-1 (CD11b/CD18), a β2 integrin mediating the firm adhesion of neutrophils to endothelium (25). Moreover, these anti-inflammatory actions of zhankuic acids A and C were not due to cytotoxic effects because there was no significant difference in cell viability compared to the vehicle control (27). The results are consistent with the non-significant cytotoxicity observed on the eight cancer cell lines tested in the present study.
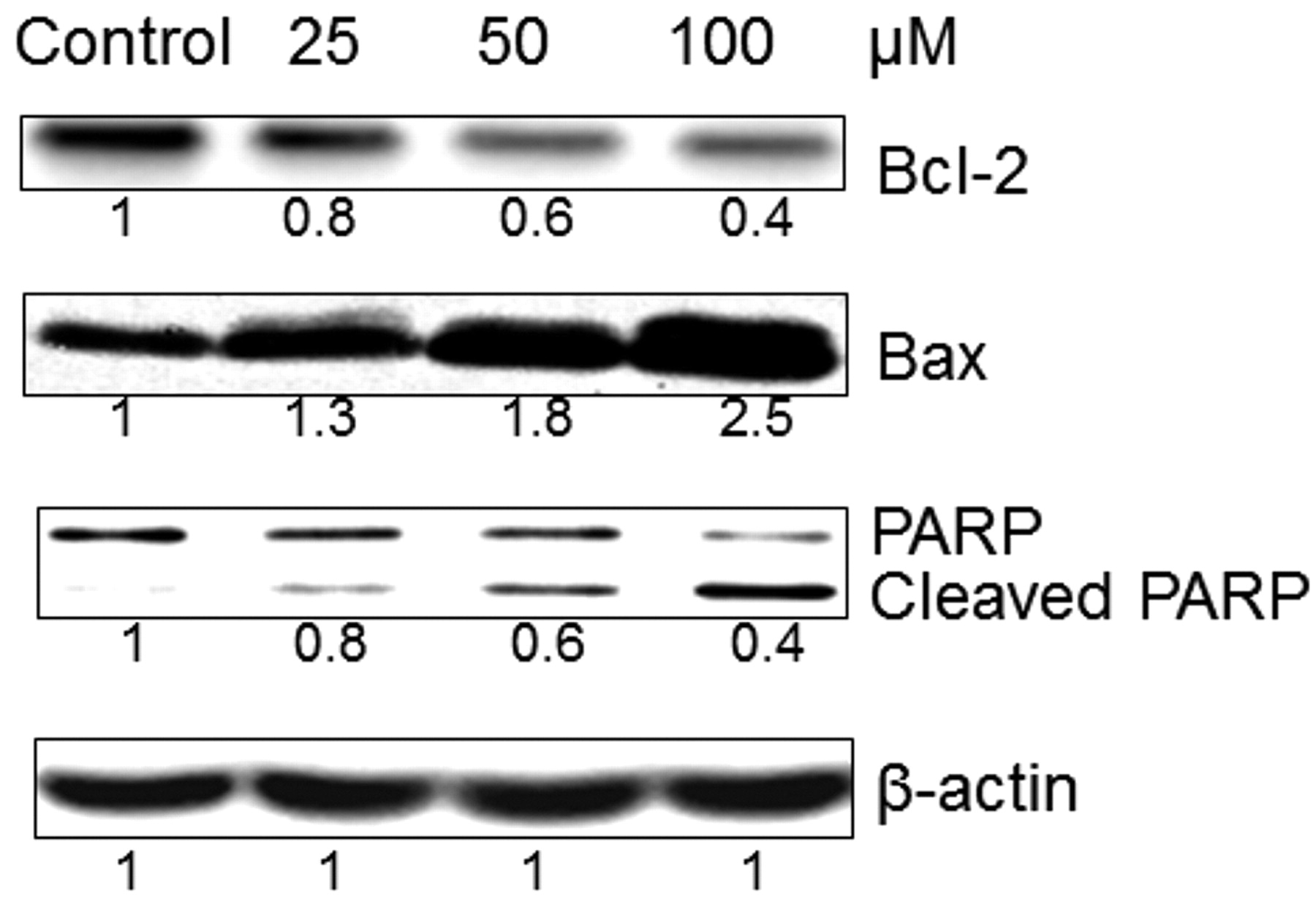
Effect of methyl antcinate A on apoptosis regulatory proteins. PC-3 cells were treated with 25 μM, 50 μM, and 100 μM of MAA or vehicle control (DMSO) for 6 h. Equal amounts of cell lysates were analyzed for Bcl-2, Bax, and PARP protein expression by western blotting with corresponding antibodies. β-Actin served as the loading control. The values are fold intensity of protein bands of MAA-treated PC-3 cells compared to the respective DMSO vehicle-treated control after being normalized to the respective internal reference (β-actin). The values in the third row denote the relative expression of full-length PARP but not cleaved PARP.
Our previous study in a liver cancer model showed that MAA, compound 11, which has more potent cytotoxic effects than Ant A and Ant C, may exert its anticancer capability via inducing ROS generation and NADPH oxidase activation (11). MAA can also up-regulate Bax, Bak, PUMA, and mitochondrial translocation of cofilin, activate caspase-3 and lead to cell death (11). We have also reported that MAA caused significant tumor growth inhibition via Bax-mediated mitochondrial apoptosis pathway in oral cancer cells (12). In the present study, MAA was further shown to have appreciable growth-inhibitory effects in KB cells, four different oral cancer cell lines (TSCCa, GNM, OC-2, and OEC-M1), Panc-1, BT474, PC-3, OVCAR-3, HeLa, and U2-OS cells. Moreover, the CC50 in MCF-12A and GF cells of MAA was 133.6 and 194.8 μM, respectively, while the IC50 of MAA in most cancer cells tested was less than 50 μM (Table I). Consequently, however, cytotoxicity towards two non-transformed cell lines was observed and MAA was found to be the most promising anticancer candidate compound among the 11 triterpenoids according to its high selectivity index (CC50/IC50). Furthermore, the expression of Bcl-2, Bax, and PARP proteins of PC-3 cells tested by western blotting suggested that MAA exerted cell death through caspase-dependent cascade and Bax-mediated mitochondrial apoptotic pathway not only on liver and oral cancer (11, 12) but also on other types of cancer cells, including prostate cancer, in a dose-dependent manner.
In summary, among the 11 triterpenoids, MAA exhibited the most potent anticancer effects. MAA is a novel compound isolated and characterized in our laboratory two years ago (10). In addition to MAA, MAB, DeEA, and 15α-acetyl-dehydrosulfurenic acid also exhibited significant selective cytotoxic effects toward cancer cells, including TsCCa, GNM, KB, OC-2, PC-3, HeLa, AGS, and U2-OS cells. Modifications of these triterpenoids may lead to the development of more potent anticancer drugs; moreover, our results highlight the necessity of examining their effects on a wide spectrum of cancer types in vivo.
Acknowledgements
This study was funded by grants from the Chung Shan Medical University (98-CCH-CSMU-09) and the National Science Council Taiwan (NSC100-2113-M-324-001-MY3). Flow cytometric analysis was performed at the Instrument Centre of Chung Shan Medical University, which is supported by the National Science Council, Ministry of Education, and Chung Shan Medical University.
Footnotes
-
↵* These Authors contributed equally to this study.
-
Conflict of Interest
None to declare.
- Received January 27, 2012.
- Revision received February 23, 2012.
- Accepted February 28, 2012.
- Copyright© 2012 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved





{kind=link}
{kind=link}
{kind=link}
{kind=link}