


Abstract
Background: This pilot study is the first to focus on the potential role for death-associated protein 3 (DAP3) in human breast cancer. Materials and Methods: A total of 153 samples were studied. The levels of transcription of DAP3 were determined using quantitative polymerase chain reaction (qPCR). Transcript levels within breast cancer specimens were compared to those of normal background tissues and correlated with clinicopathological data accumulated by over a 10-year follow-up period. Results: The expression of DAP3 mRNA was demonstrated to decrease with increasing Nottingham Prognostic Index (NPI2 vs. 3, p=0.036), TNM stage (TNM1 vs. 3, p=0.07), and tumour grade (grade 1 vs. 3, p=0.08). Lower DAP3 expression levels were significantly associated with local recurrence (p=0.013), distant metastasis (p=0.0057) and mortality (p=0.019). Conclusion: This study demonstrates an inverse association between DAP3 mRNA levels and tumour stage and clinical outcome in breast cancer, consistent with the pro-apoptosis function of DAP3. Further research is required in order to confirm our findings and clarify the mechanisms that regulate DAP3 expression in human breast cancer.
The role of apoptosis genes in oncogenesis has been the subject of much research in the past. The earliest and most well documented gene is the so-called ‘guardian of the genome’; p53. Since then, a number of other genes have been studied for their role in programmed cell death, including, but not limited to, BAX, BCL2, and those encoding caspases 3, 6, 8 and 9. Silencing of these genes has been postulated as a possible route to oncogenesis, and this has been demonstrated in various studies (1, 2).
The death-associated proteins (DAP) are a relatively novel group of proteins found to have a role in programmed cell death. Four proteins were initially identified; DAP kinase, and DAP1, -3 and -5 (3).
DAP3 has been studied in vitro and in murine models, and has been found to be a highly conserved protein, with a role in apoptosis and in mitochondrial function. More recent studies have also identified two possible associated regulatory proteins; death ligand signal enhancer (DELE, also referred to as KIAA0141) and interferon-beta promoter stimulator 1 (IPS1, also referred to as PPIP5K1) (4-6).
The role of DAP3 in cancer has been overlooked in the recent literature. In this pilot study, we investigated the relationship between the expression of DAP3 and breast cancer. Potentially this would open a potentially significant avenue of research to further our understanding of the mechanisms behind oncogenesis.
Materials and Methods
Samples. Institutional guidelines including ethical approval and informed consent were adhered to. Immediately after surgical excision, a tumour sample was obtained from the tumour area while another was obtained from the associated non-cancerous tissue (ANCT) within 2 cm from the tumour area, without affecting the assessment of tumour margins. Breast cancer tissues (n=127) and normal background tissues (n=33) were collected and stored at −140°C in liquid nitrogen until the commencement of this study.
All the patients were treated according to local guidelines, following discussions in multidisciplinary meetings. Patients undergoing breast conserving surgery also underwent radiotherapy. Hormone-sensitive patients were aministered tamoxifen. Hormone-insensitive cases, high-grade cancer, and node-positive cases were treated with adjuvant therapy. Clinicopathological data (Table I) was collected from the patient charts, and was collated in an encrypted database.
RNA extraction kits and reverse transcription kits were obtained from AbGene Ltd. (Surrey, UK). PCR primers were designed using Beacon Designer (Palo Alto, CA, USA) and synthesised in-house. Custom made hot-start Master Mix for quantitative PCR was purchased from AbGene (7).
Clinical data showing number of patients in each category.
Tissue processing, RNA extraction and cDNA synthesis. Approximately 10 mg of cancerous tissue were homogenised. A larger amount of ANCT (20-50 mg) was used, as its high fat content made it difficult to obtain a sufficient RNA concentration for analysis. The concentration of RNA was determined using a UV spectrophotometer (Wolf Laboratories, York, UK) to ensure adequate amounts of RNA for analysis. Reverse transcription was carried out using a reverse transcription kit (AbGene) with an anchored olig (dT) primer using 1 mg of total RNA in a 96-well plate to produce cDNA. The quality of cDNA was verified using β-actin primers (primers 5’-ATGATATCGCCGCGCTCGTC-3’ and 5’-CGCTCGGTGAGGATCTTCA-3’) (8).
Quantitative analysis. Transcripts of cDNA library were determined using real-time quantitative PCR based on the Amplifluor technology. The PCR primers were designed using Beacon Designer software, but an additional sequence, known as the Z sequence (5’-ACTGAACCTGACCGTACA-3’) which is complementary to the universal Z probe (Intergen Inc., Oxford, UK) was added to the primer. The primers used are detailed in Table II.
The reaction was carried out under the following conditions: 94°C for 12 min and 50 cycles of 94°C for 15 s, 55°C for 40 s, and 72°C for 20 s. The levels of each transcript were generated from a standard plasmid which contained the specific DNA sequence that was simultaneously amplified within the samples.
With every run of the PCR, a negative and positive control was employed, using a known cDNA sequence (7) .
Statistical analysis. Analysis of the data was performed using the Minitab 14.1 statistical software package (Minitab Ltd. Coventry, UK.) using a custom-written macro (Stat 2005.mtw). Independent variables were compared using the Mann-Whitney U-test while paired variables were compared using the two-sample t-test. The transcript levels within the breast cancer specimens were compared to those of the ANCT and correlated with clincopathological data collected over a 10-year follow-up period.
Primers used in RT-PCR analysis.
Mean DAP3 mRNA expression levels (copy number) in a cohort of 127 breast cancer patients; a comparison between subgroups with different tumour grade, Nottingham prognostic index (NPI), and TNM stage.
P-values less than 0.05 were considered significant whereas p-values between 0.05 and 0.10 were considered marginally significant.
For purposes of the Kaplan-Meier survival analysis, the samples were divided arbitrarily into high and low transcription groups, with the value of the moderate prognostic group as defined by NPI serving as the dividing line. Survival analyses were performed using SPSS version 12.0.1 (SPSS Inc., Chicago, IL, USA).
Results
In the course of our study, we recorded a strong inverse correlation between mean copy number of DAP3 and survival. The mean copy number in patients with no disease recurrence was significantly higher than that in patients with local recurrence (122 vs. 12.9, p=0.013), distant recurrence (122 vs. 1.67, p=0.0057), and with mortalities (122 vs. 19.1, p=0.019).
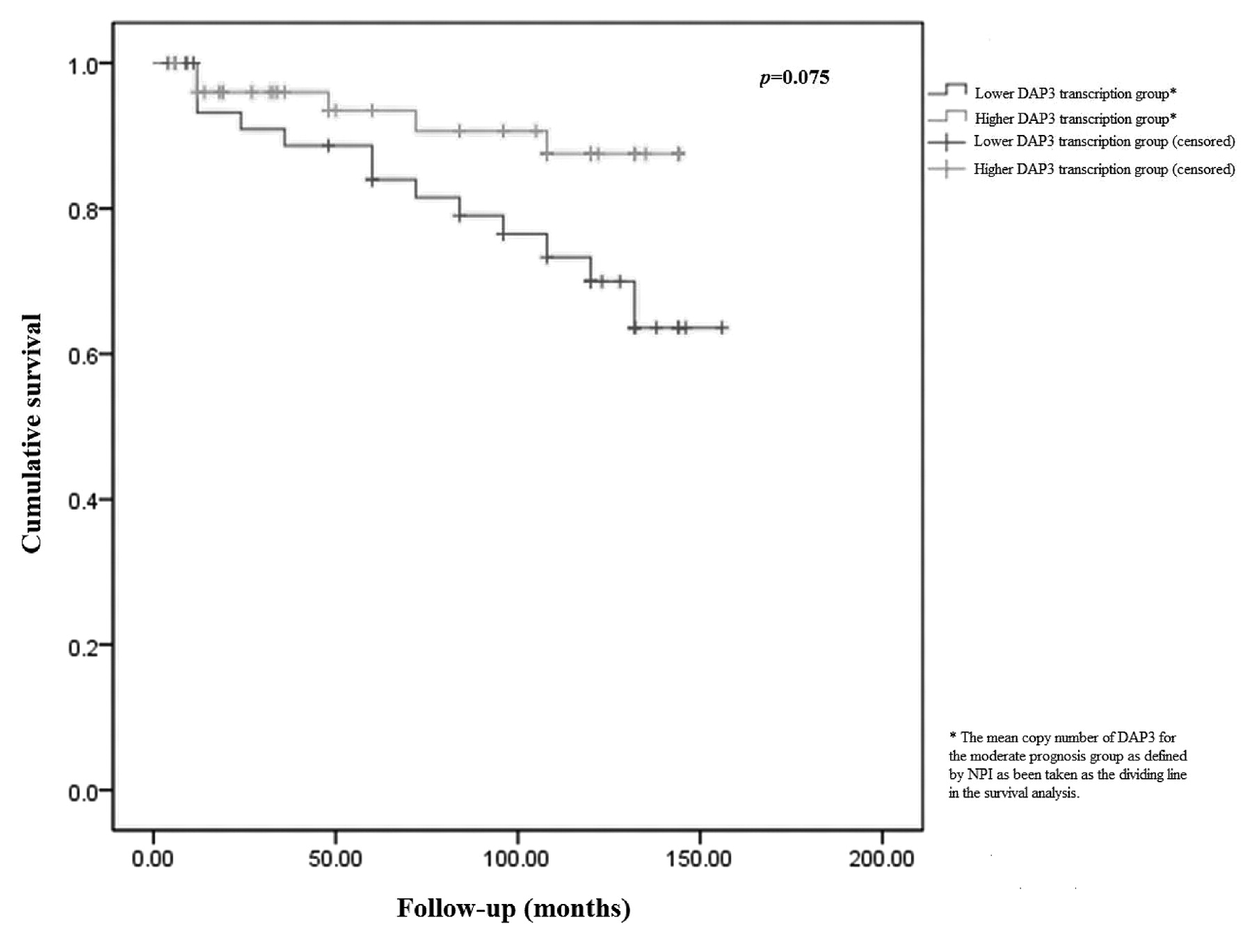
Overall survival curve.
The expression of DAP3 has been shown to decrease in relation to tumour grade (a mean copy number of 305 for grade 1 vs. 50 for grade 3, p=0.082). Similarly, a decreasing trend in DAP3 expression is seen in relation to the NPI (a mean copy number of 134 for NPI1 vs. 15.9 for NPI3, p=0.036).
There was an inverse correlation between DAP3 and TNM staging. A marginally significant decrease in mean DAP3 copy number was seen between TNM1 and TNM3 (104 vs. 15.9, p=0.07).
The two-sample test showed that mean DAP3 mRNA levels were significantly lower in tumours associated with local recurrence (122 for disease-free vs. 12.9 for local recurrence, p=0.013), distant recurrence (122 for disease-free vs. 1.67 for distant recurrence, p=0.0057), and breast cancer-specific mortality (122 for disease-free vs. 19.1 for breast cancer-specific mortality, p=0.019). The Kaplan- Meier plot analysis has demonstrated that better overall survival in patients expressing higher levels of DAP3 approached statistical significance (p =0.075) (Figure 1) .
Discussion
This is the first study in the literature to examine the direct relationship between DAP3 mRNA expression and human breast cancer. Our findings demonstrate that this gene may have a tumour suppressor function through promoting apoptosis and could act as a useful prognostic indicator in human breast cancer.
The DAP3 gene has been localised to chromosome 1 band q21, and encodes a 46 kDa protein (9) which is a highly conserved a GTP-binding protein. DAP3 is suppressed when phosphorylated by AKT (10), and has been demonstrated to be necessary for mitochondrial function. This role is distinct from that it has as an apoptotic protein. Kim et al. reported a deficiency of cytochrome c oxidase 1 in the mitochondria of Dap3 −/− mouse embryos. As this is a protein encoded in the mitochondrial genome, it is inferred that DAP3 has an integral role in intra-mitochondrial protein synthesis (11) .
Li and colleagues described a role for DAP3 in anoikis (the induction of apoptosis in detached epithelial cells from extracellular matrix). This function is mediated via IPS-1, which upon binding with DAP3 stimulates caspases 3, 8 and 9(4) .
Harada et al. described a further regulator protein for DAP3, namely DELE. It is suggested that DELE responds to apoptosis induction by tumour necrosis factor alpha (TNF-α) and TNF-related apoptosis-inducing ligand (TRAIL) by binding with DAP3, and stimulating caspases 3, 8 and 9 (5). Furthermore, Takeda et al. observed that DAP3 expression induced apoptosis in osteosarcoma cells (12) .
Such studies support and explain our finding of a compelling trend for DAP3 mRNA levels to be lower in cancerous tissues and in patients who developed progressive disease (distal metastasis, local recurrence, or died of breast cancer). These observations are consistent with a possible tumour suppressor function of this gene through the activation of the apoptosis pathway in breast cancer.
Counterintuitively, Mariani et al. reported increased expression levels of DAP3 in invasive glioblastoma tumour cells, and in glioma cell lines with induced migratory phenotype (13). These observations can be explained by the fact that P-loop mutant DAP3 is less effective in inducing apoptosis, and the COOH-terminal deleted protein (230 amino acids) acts in a dominant negative fashion, protecting cells from induced apoptosis (14). Furthermore, such findings could reflect a different balance between the mitochondrial maintenance and the pro-apoptotic functions of the DAP3 levels in glioma compared with human breast cancer. However, any inferences one may derive from the aforesaid in relation to the findings in glioma cell lines, or indeed with regards to the migration of neoplastic cells in general, whilst tempting, would be speculative without further investigation.
The strength of our report lies in the use of robust RT-PCR methodology to analyze DAP3 mRNA expression in a cohort of breast cancer patients with a long-term follow up. However there are several inherent limitations to our study related to the relatively small sample size and lack of data regarding protein expression of the studied gene.
Further research is required to confirm the role of DAP3 gene in the pathogenesis of breast cancer including immunohistochemistry studies, in vitro experiments and the preparation of animal models with suppressed DAP3 gene. In addition to its potential role as a prognostic indicator, DAP3 could represent a potential therapeutic target in human breast cancer.
- Received November 26, 2011.
- Revision received December 16, 2011.
- Accepted December 21, 2011.
- Copyright© 2012 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
Jump to section
Related Articles
Cited By...
- Expression of Death Associated Proteins DAP1 and DAP3 in Human Pancreatic Cancer
- Stratification Using hTERT and Stem Cell Markers Confers a Good Prognosis in Invasive Breast Cancer
- Correlation of TERT and Stem Cell Markers in the Context of Human Breast Cancer
- Lactate-mediated mitoribosomal defects impair mitochondrial oxidative phosphorylation and promote hepatoma cell invasiveness
- The Interaction Between DAP1 and Autophagy in the Context of Human Carcinogenesis
- P14ARF Is Down-regulated During Tumour Progression and Predicts the Clinical Outcome in Human Breast Cancer
- The mRNA Expression of Inhibitors of DNA Binding-1 and -2 Is Associated with Advanced Tumour Stage and Adverse Clinical Outcome in Human Breast Cancer
- Guanine Nucleotide Binding Protein {beta} 1: A Novel Transduction Protein with a Possible Role in Human Breast Cancer
- The mRNA Expression of DAP1 in Human Breast Cancer: Correlation with Clinicopathological Parameters