


Abstract
Background: Adenovirus vectors have been utilized for a variety of cancer gene therapy. The present study examined the oncolytic effect of adenovirus type 5 (Ad5) and fiber-substituted conditionally replicating adenovirus (CRAD) Ad5/F35 vectors on human lung cancer A549 cells (an epithelial adenocarcinoma cell line), SBC-3 cells (a small-cell cancer cell line), and Lu-65 cells (a giant-cell lung cancer cell line). Materials and Methods: For adenovirus, the first mRNA/protein to be made (~1 h after infection) is early region 1A (E1A). Ad5F35 and Ad5 CRAD vectors containing the E1 gene, controlled by the human midkine promoter (Ad5F35/MKp-E1 and Ad5/MKp-E1, respectively) were constructed. Reverse transcription-polymerase chain reaction (RT-PCR), western blotting and cell viability assays were carried out in cells transfected with Ad5/MKp-E1 and Ad5F35/MKp-E1. Results: Less expression of mRNA and protein for coxsackie and adenovirus receptor (CAR), a cell surface target of Ad5, was found with lung cancer cells as compared with the expression in human embryonic kidney 293 (HEK293) cells, but otherwise mRNA and CD46 protein, a cell surface target of Ad35, was expressed in lung cancer cells as much as in HEK293 cells. Both Ad5/MKp-E1 and Ad5F35/MKp-E1 induced oncolysis of lung cancer cells in a viral particle-dependent manner, with more efficient advantage for Ad5F35/MKp-E1. Conclusion: The results of the present study suggest that Ad5F35/MKp-E1 is more useful for the gene therapy of human lung cancer than Ad5/MKp-E1 is.
Adenovirus vectors have been utilized for cancer gene therapy (1). Conditionally replicating adenovirus (CRAD) represents a promising class of biological agents that selectively replicate in and lyse cancer cells, i.e. CRAD enables induction of tumor-specific cell death and amplifies oncolysis by intratumor replications (2-4). In addition, CRAD enables oncolysis of cancer cells neighboring primarily infected cells by secondary infection, to an extent greater than that of non-replicating adenoviruses (5-7).
The infection efficacy of adenovirus is the most critical factor determining whether beneficial results are obtained or not (8, 9). The adenovirus infects into cells through individual target receptors on the membrane surface. Adenovirus type 5 (Ad5) has been widely used for gene therapy. Ad5 is transfected into cells through coxsackie and adenovirus receptor (CAR) (10). In contrast, adenovirus type 35 (Ad35) is transfected into cells through cluster of differentiation 46 (CD46) (11). Generally, cancer cells exhibit much lower expression of CAR as compared with CD46, suggesting that more efficient oncolytic effect on cancer cells would be obtained with Ad35 than Ad5.
The present study examined the oncolytic effect of CRAD vector encoding the early region 1 (E1) gene under the control of 0.6 kb midkine promoter without (Ad5/MKp-E1) and with replacement of the fiber knob for Ad5 compared to that of Ad35 (Ad5F35/MKp-E1) on human lung cancer A549 cells (an epithelial adenocarcinoma cell line), SBC-3 cells (a small-cell cancer cell line), and Lu-65 cells (a giant-cell lung cancer cell line). We showed that Ad5F35/MKp-E1 is useful for gene therapy of human lung cancer, presenting with more efficient advantage than Ad5/MKp-E1.
Materials and Methods
Construction of CRAD vectors. Replication-competent adenovirus vector containing the E1 gene controlled by the midkine promoter was prepared as follows: The cytomegalovirus (CMV) promoter of pShuttle2 vector (Clontech Laboratories, Mountain View, CA, USA) was replaced with the 0.6 kb 5’-upstream regulatory region of the midkine gene and the pS-MK/E1 vector was produced by inserting the E1 gene downstream the midkine region. To construct pAd5F35, the Ad5 fiber region on the pAdeno-X vector (Clontech Laboratories) was substituted with the Ad35 fiber region on RHSP vector (Avior Therapeutics, Seattle, WA, USA). Ad5/MKp-E1 and Ad5F35/MKp-E1 were prepared by linking the pS-MK/E1 vector with pAdeno-X and pAd5F35, respectively.
Primers used for reverse transcription-polymerase chain reaction.
Cell culture. A549, SBC-3, and Lu-65 cells were purchased from the Health Science Research Resources Bank (Osaka, Japan), and human embryonic kidney 293 (HEK293) cells from RIKEN Bioresource Center (Tsukuba, Japan). A549 and SBC-3 cells were grown in minimum essential medium with 0.1 mM non-essential amino acids; Lu-65 cells were cultured in RPMI-1640, and HEK293 cells were cultured in Dulbecco's modified Eagle's medium, each supplemented with 10% (v/v) heat-inactivated fetal bovine serum, penicillin (final concentration, 100 U/ml), and streptomycin (final concentration, 0.1 mg/ml), in a humidified atmosphere of 5% CO2 and 95% air at 37°C.
Reverse transcription-polymerase chain reaction (RT-PCR). RT-PCR was carried out using primers as shown in Table I by the method described previously (12). The expression levels for each mRNA were normalized by the levels for glyceraldehyde-3-phosphate dehydrogenase (GAPDH) mRNA.
Western blotting. Western blotting was carried out by the method described previously (13). Briefly, cells were lysed with 1% (w/v) sodium dodecyl sulfate (SDS). Proteins were separated by SDS-polyacrylamide gel electrophoresis (SDS-PAGE) using a TGX gel (BioRad, Hercules, CA, USA) and then transferred to polyvinylidene difluoride membranes. Membranes were blocked with TBS-T [150 mM NaCl, 0.1% (v/v) Tween20 and 20 mM Tris, pH 7.5] containing 5% (w/v) bovine serum albumin and subsequently incubated with antibodies against CAR (Santa Cruz Biotechnology, Santa Cruz, CA, USA), CD46 (Santa Cruz Biotechnology), and β-actin (Sigma, St Louis, MO, USA)
Oncolysis assay. Cells (5×103) in 96-well plates were infected with Ad5/MKp-E1 or Ad5F35/MKp-E1 at 0.01-10000 viral particles (VP)/cell. Five days after infection, cells were incubated with Alamar Blue (Life Technologies, Gaithersburg, MD, USA) and the number of viable cells counted by detecting absorbance at 560 nm with a high-throughput microplate spectrophotometer (SpectraMax Plus 384; Molecular Devices, Sunnyvale, CA, USA).
Results
Expression of CAR and CD46. All lung cancer cell lines examined here expressed the midkine mRNA, with the lowest expression for SBC-3 cells among HEK293 cells and lung cancer cells (Figure 1A and B). HEK293 is recognized to abundantly express CAR and CD46. In the present study, lower expression of CAR mRNA and protein was found in lung cancer cells as compared with the expression in HEK293 cells (Figure 1A and C, and 2A). In contrast, the expression level of CD46 protein for lung cancer cells was similar to that for HEK293 cells, although the expression level of CD46 mRNA for SBC-3 and Lu-65 cells, but not A549 cells, was lower than that for HEK293 cells (Figure 1A and D, and 2B).
Oncolytic effects of Ad5/MKp-E1 and Ad5F35/MKp-E1 on lung cancer cells. For HEK293 cells, Ad5/MKp-E1 and Ad5F35/MKp-E1 reduced cell viability in a VP-dependent manner, with the Ad5F35/MKp-E1 efficacy being higher than that of Ad5/MKp-E1 (Figure 3A). Ad5/MKp-E1 and Ad5F35/MKp-E1 also reduced cell viability in a VP-dependent manner for all the lung cancer cell lines, with beneficial advantage for Ad5F35/MKp-E1 (Figure 3B-D).
Discussion
Expression of midkine, a heparin-binding growth factor, is induced by retinoic acid in embryonal carcinoma cells (14). Mounting evidence has pointed to the role of midkine in mitogenesis, angiogenesis, anti-apoptosis, fibrinolysis, and transformation (15-19). Midkine is abundantly expressed in a variety of cancer cells originating from the esophagus, stomach, colon, liver, breast, and pancreas, while its expression in non-malignant cells is quite limited, with moderate expression in the kidney and weak expression in the lung, colon, and thyroid gland (20-24). The midkine promoter, therefore, is widely utilized for suicide gene therapy.
In the present study, we constructed Ad5 (Ad5/MKp-E1) and Ad5F35 CRAD vectors (Ad5F35/MKp-E1) encoding the E1 gene under the control of 0.6 kb midkine promoter. Ad5 and Ad35 are infected into cells through CAR and CD46, respectively (10, 11). CD46 is more commonly expressed in cancer cells as compared to CAR (25). The expression level of CD46 for lung cancer cell lines here was similar to that for HEK293 cells, although expression of CAR in lung cancer cells was lower than that in HEK293 cells.
Ad5F35/MKp-E1 reduced cell viability in a VP-dependent manner for all the lung cancer cell lines more efficiently than Ad5/MKp-E1 did. This suggests that the oncolytic effect of Ad5F35/MKp-E1 depends on the expression level of CD46.
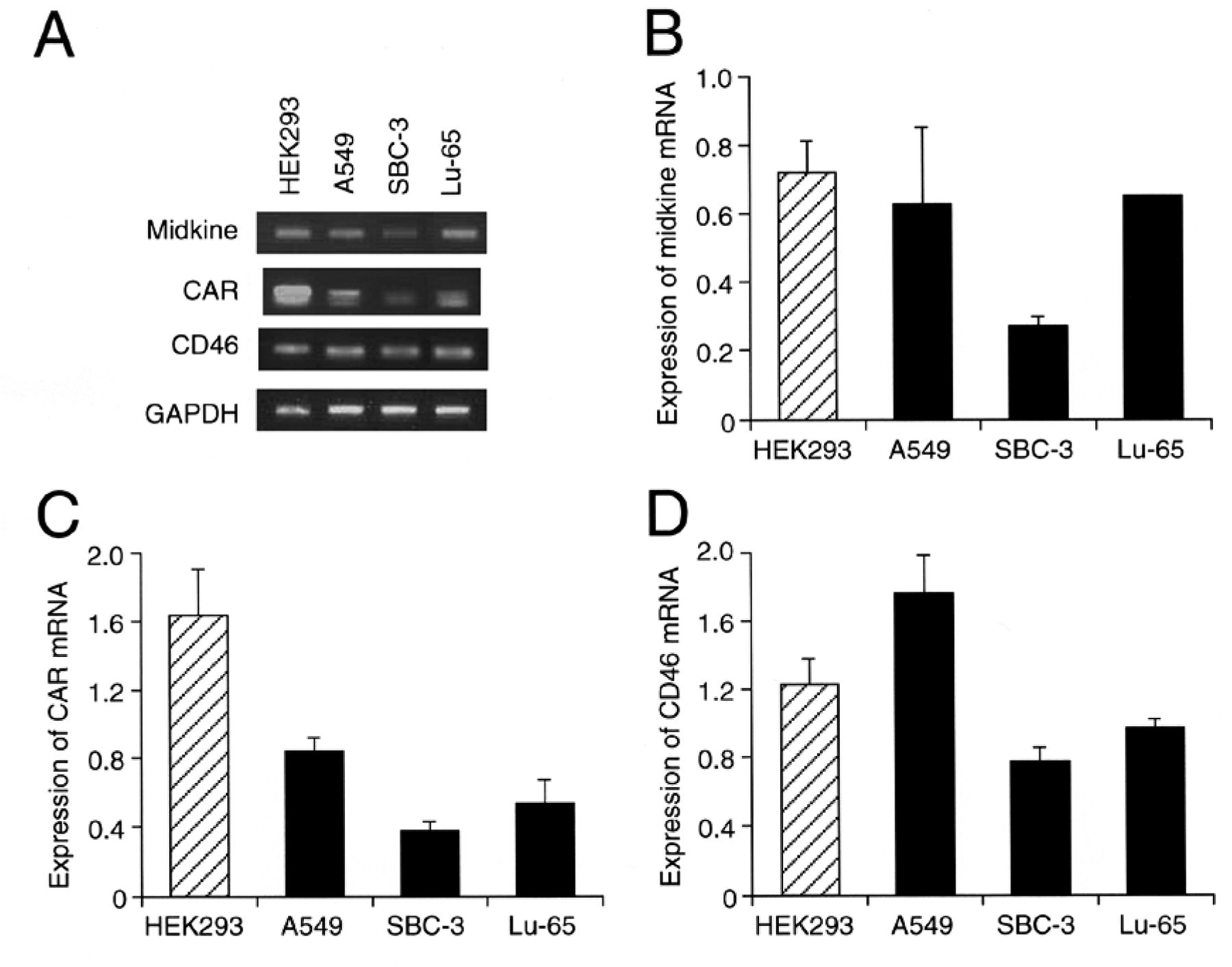
Expression of the mRNA for midkine, CAR and CD46. RT-PCR of cells was carried out as indicated. RT-PCR products are shown in (A). In the graphs, each column represents the mean (±SEM) expression of mRNA for midkine (B), CAR (C), and CD46 (D) (n=3 independent experiments).

Expression of CAR and CD46 protein. Western blotting for CAR (A) and CD46 (B) were carried out in HEK293 (HEK), A549, SBC-3, and Lu-65 cells. The expression levels of each protein were normalized by those for β-actin. In the graphs, each column represents the mean (±SEM) expression of each protein (n=4 independent experiments).
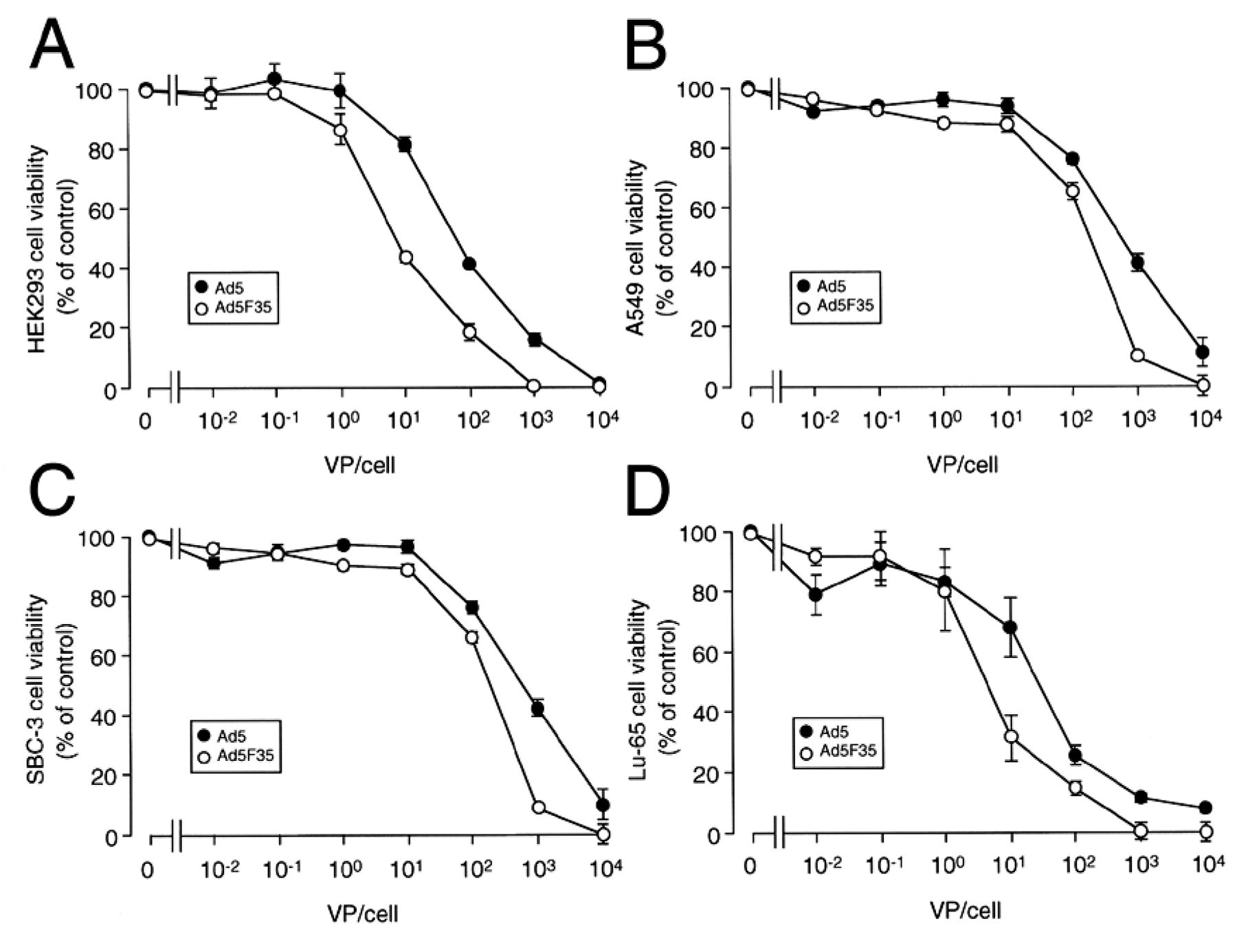
Oncolytic effects of Ad5/MKp-E1 and Ad5F35/MKp-E1. Cell viability was assayed 5 days after infection using Alamar Blue in HEK293 (A), A549 (B), SBC-3 (C), and Lu-65 cells (D). In the graphs, each point represents the mean (±SEM) percentage viability relative to the basal level (viabilities of cells uninfected) (n=6 independent experiments).
The Ad5 vector containing the Arg-Gly-Asp (RGD) motif in the Ad fiber knob (Ad5RGD) induces oncolysis of renal cancer cells by targeting the RGD receptor integrin (26). The transduction efficacy of the Ad5RGD vector for renal cancer cells, however, is much lower than the efficacy of Ad5F35/MKp-E1 (26). Overall, the results of the present study indicate that Ad5F35/MKp-E1 may be more useful for the gene therapy of lung cancer than the Ad5RGD vector or Ad5/MKp-E1.
Conclusion
The results of the present study show that Ad5/MKp-E1 and Ad5F35/MKp-E1 induced oncolysis of lung cancer cells in a VP-dependent manner, with an efficient advantage for Ad5F35/MKp-E1. This may offer new hope for development of lung cancer gene therapy.
Footnotes
-
Conflicts of Interest
None of the Authors has any potential conflicts of interest.
- Received August 27, 2012.
- Revision received October 9, 2012.
- Accepted October 10, 2012.
- Copyright© 2012 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved





{kind=link}
{kind=link}
{kind=link}