


Abstract
Background: We previously reported that epigenetic silencing of microRNA-34b/c (miR-34b/c) plays an important role in the pathogenesis of malignant pleural mesothelioma (MPM). We examined the impact of miR-34b/c restoration on the radiosensitivity of MPM cells. Materials and Methods: We established stable miR-34b/c and scramble transfectants of two MPM cell lines, H2052 and H28. We examined these transfectants by clonogenic survival assay, phosphorylated histone H2AX (γH2AX) foci assay, cell-cycle analysis, and western blotting. Results: The clonogenic survival assay revealed that miR-34b/c radiosensitized MPM cells. γH2AX foci assay showed that DNA double-strand break repair was delayed in miR-34b/c transfectants. The proportion of sub-G1 phase cells was increased in miR-34b/c transfectants after irradiation. miR-34b/c inhibited expression of cyclin-D1, cyclin-dependent kinase 4/6, B-cell lymphoma-2 (BCL-2) and increased cleaved poly (ADP-ribose) polymerase (cPARP) and cleaved caspase-3 after irradiation. Conclusion: Our results indicate that miR-34b/c enhances radiosensitivity by promoting radiation-induced apoptosis and suggested that miR-34b/c might be a useful therapeutic molecule to enhance radiotherapy in MPM.
Malignant pleural mesothelioma (MPM) is an aggressive neoplasm arising from the pleura. The prognosis of MPM is dismal, as neither chemotherapy nor radiotherapy remarkably improve its prognosis (1). Thus, an urgent need to understand the molecular biology of MPM exists so that new therapeutic strategies for MPM can be developed (2).
MicroRNAs (miRs) are a class of 17-22 nucleotide small non-coding RNAs (3). Among them, miR-34b/c has an upstream p53-binding site and is directly induced by p53 in response to oncogenic stress and DNA damage (4, 5). The known targets for miR-34b/c include mesenchymal epithelial transition factor (c-MET), cyclin-dependent kinase (CDK) 4/6, cyclin D1 (CCND1), cyclin E2 (CCNE2) (4), and BCL2 (6). Since miR-34b/c suppresses these targets, miR-34b/c is thought to play an important role in the p53 tumor suppressor network.
We previously reported that miR-34b/c was frequently methylated in MPM, compared with miR-34a (7). Aberrant methylation in the promoter region of miR-34b/c was present in 6 out of 6 MPM cell lines and in 40 (85.1%) out of 47 primary MPMs, whereas methylation of miR-34a was present in 2 (33.3%) out of 6 MPM cell lines and in 13 (27.7%) out of 47 primary MPMs. Interestingly, the introduction of miR-34b/c, but not p53, induced significant antitumor effects such as G1-arrest, apoptosis, and the inhibition of migration, invasion and cell motility of MPM cells (7); these effects arose through the suppression of multiple targets of miR-34b/c, indicating the importance of miR-34b/c in MPM.
Although genetic alteration of p53 is rare in MPM (8), MPMs biologically exhibit an apparent p53-deficiency, resulting in anti-apoptosis effects and cell-cycle alterations. CCND1, one of the targets of miR-34b/c, is an important component of the core cell-cycle machinery. Recently, CCND1 was reported to be a therapeutic target for MPM (9). One study showed that the reduction of CCND1 enhances radiosensitivity of human cancer by suppressing DNA double-strand break (DSB) repair (10). Another report described that flavopiridol, a pan-CDK inhibitor, also enhances radiosensitivity and induces apoptosis in non-small cell lung cancer (11), as well as gastric and colon cancer (12).
After considering these points, we hypothesized that the introduction of miR-34b/c might restore p53-mediated tumor suppression in response to ionizing radiation (IR) in MPM cells. In this study, we examined the impact of miR-34b/c on the cellular radiosensitivity of MPM cells.
Materials and Methods
Cell culture and establishment of stable miR-34b/c transfectants. Two MPM cell lines (H2052 and H28) in which the expressions of miR-34b and miR-34c were epigenetically suppressed were used in this study. The two MPM cell lines were kindly provided by Dr. Adi F. Gazdar (Department of Pathology, University of Texas Southwestern Medical Center, Dallas, TX, USA) and were maintained in RPMI-1640 (Sigma Chemical Co., Saint Louis, MO, USA) supplemented with 10% fetal bovine serum in 5% CO2. Stable miR-34b/c and scramble RNA transfectants were established, as previously described in our report (7). Briefly, a fragment of genomic DNA encoding miR-34b/c or scramble RNA fragment as a negative control was subcloned into a pSilencer 4.1-CMV neo plasmid vector (Ambion, Austin, TX, USA). The constructed plasmids were then transfected into MPM cells using Lipofectamine 2000 Reagent (Invitrogen, Carlsbad, CA, USA). Resistant clones were isolated by G418 selection for two weeks and were maintained in G418-containing medium.
Evaluation of miR-34b/c expression using quantitative real-time-polymerase chain reaction (PCR). microRNA was extracted from miR-34b/c-transfected and scramble RNA-transfected MPM cells using TaqMan MicroRNA Cells-to-CT™ Kit (Ambion). Real-time PCR for miR-34b and 34c was performed using TaqMan MicroRNA Assays (P/N4427975, Assay ID 000427 for has-miR-34b, Assay ID 000428 for has-miR-34c; Applied Biosystems, Foster City, CA) using the StepOnePlus™ Real-Time PCR system (Applied Biosystems). The expression of miR-374 was used as an endogenous control following the manufacturer's recommendation (www.appliedbiosystems.com). The relative expression ratio for miR-34b and miR-34c was calculated from triplicate PCR.
Clonogenic cell survival assay. miR-34b/c-transfected and scramble RNA-transfected H2052 and H28 cells were trypsinized to generate a single-cell suspension, and a specified number of cells were seeded into each well in six-well tissue culture plates. After allowing the cells time to attach (6 h), the plates were irradiated at 2, 4, 6 or 8 Gy. Fourteen days after IR, colonies were stained with crystal violet. The number of colonies containing at least 50 cells was determined, and survival curves were generated. The dose enhancement factor (DEF) was calculated at a surviving fraction of 0.1 to estimate the increase in radiosensitivity. Data presented are the mean±standard deviation (SD) from at least two independent experiments.
Immunofluorescent staining for phosphorylated histone H2AX (γH2AX). γH2AX has been established as a sensitive indicator of DNA DSB with the resolution of foci corresponding to DNA DSB repair (13). miR-34b/c-transfected and scramble RNA-transfected H2052 cells were grown in Lab-Tek chamber slides (Nalge Nunc International, Naperville, IL, USA). The cells were fixed in 4% paraformaldehyde for 10 min at 1 h, 6 h, and 24 h after IR (4 Gy) and permeabilized in phosphate-buffered saline (PBS) containing 0.2% NP40 for 15 min. The cells were then incubated overnight at 4°C with 1:200 diluted anti-γH2AX antibody (Millipore, Billerica, MA,USA). The cells were incubated with 1:50 diluted fluorescein isothiocyanate (FITC)-labeled secondary antibody (Jackson Immuno Research Labs, West Grove, PA, USA) for 1 h and were then incubated in PBS, containing 4’,6-diamidino-2-phenylindole (1 μg/ml) for 30 min. Coverslips were mounted using an anti-fade solution (DAKO Corp., Carpinteria, CA, USA). The slides were examined under a fluorescent microscope (Keyence BZ-8000; Keyence, Osaka, Japan). The number of γH2AX foci was counted in each nucleus, of at least 50 cells in each sample.
Cell-cycle analysis. The cell-cycle distribution was evaluated using flow cytometry in miR-34b/c-transfected and scramble RNA-transfected H2052 cells. Non-irradiated transfectants and 4 Gy-irradiated transfectants at 24 h after IR were harvested and resuspended in PBS containing 0.2% Triton X-100 and 1 mg/ml RNase, then stained with propidium iodide and analyzed using a FACScan instrument, as previously described (7). Doublets, cell debris, and fixation artifacts were gated out, and cell-cycle analysis was performed using the CellQuest version 3.1 software.
Western blot analysis. Cells were harvested and western blot analysis was carried out as previously described (7). We examined four molecules (CCND1, CDK4, CDK6, and BCL2) reported to be primary targets of miR-34b/c. We also examined BCL2 Associated X-protein (BAX), cPARP, pro-caspase-3, and cleaved caspase-3 to detect apoptosis. Actin was used as the loading control. The primary antibodies used for western blotting were as follows: anti-CCND1 (sc718, diluted 1000:1; Santa Cruz, Santa Cruz, CA, USA), anti-CDK4 (sc260, diluted 200:1; Santa Cruz), anti-CDK6 (#3136, diluted 2000:1; Cell Signaling), anti-BCL2 (sc7382, diluted 500:1; Santa Cruz), anti-BAX (#2774, diluted 1000:1; Cell Signaling Technology, Beverly, MA, USA), anti-cPARP (#9546, diluted 2000:1; Cell Signaling), anti-pro-caspase-3 (sc7272, diluted 2000:1; Santa Cruz), and anti-actin (MAB1501, diluted 20000:1; Millipore).
Statistical analysis. Data are represented as the mean±SD. An unpaired Student's t-test was used to compare data between the two groups. Probability values less than 0.05 were considered statistically significant. All the data were analyzed using JMP9.0 for Windows (SAS Institute, Cary, NC, USA).
Results
Evaluation of miR-34b/c expression using quantitative real-time PCR. The expressions of miR-34b and 34c were examined in miR-34b/c and scramble RNA transfectants. Compared with the scramble RNA transfectants, the expression of miR-34b was increased by 26.6-fold and 10.5-fold in miR-34b/c-transfected H2052 and H28 cells, respectively. The expression of miR-34c was increased by 68.7-fold and 31-fold in miR-34b/c-transfected H2052 and H28 cells, respectively. Compared with the increase of miR-34b and miR-34c in miR-34b/c transfectants, the expression of miR-34b and miR-34c in scramble RNA-transfected H2052 cells at 24 hours after 4 Gy IR was only increased by 1.2-fold and 1.4-fold, respectively. Thus, we confirmed that the expression of mature miR-34b and miR-34c were increased in stable miR-34b/c transfectants.
Effect of miR-34b/c on cellular radiosensitivity. A clonogenic survival assay was performed to examine the effect of miR-34b/c on radiosensitivity in miR-34b/c transfectants, compared with the scramble control. As shown in Figure 1, miR-34b/c enhanced radiosensitivity of both miR-34b/c-transfected H2052 and H28 cells, with a DEF of 1.3 and 1.2, respectively. Because the radiosensitizing effect of miR-34b/c was stronger in H2052 cells than that in H28 cells, subsequent experiments were performed using H2052-derived transfectants.
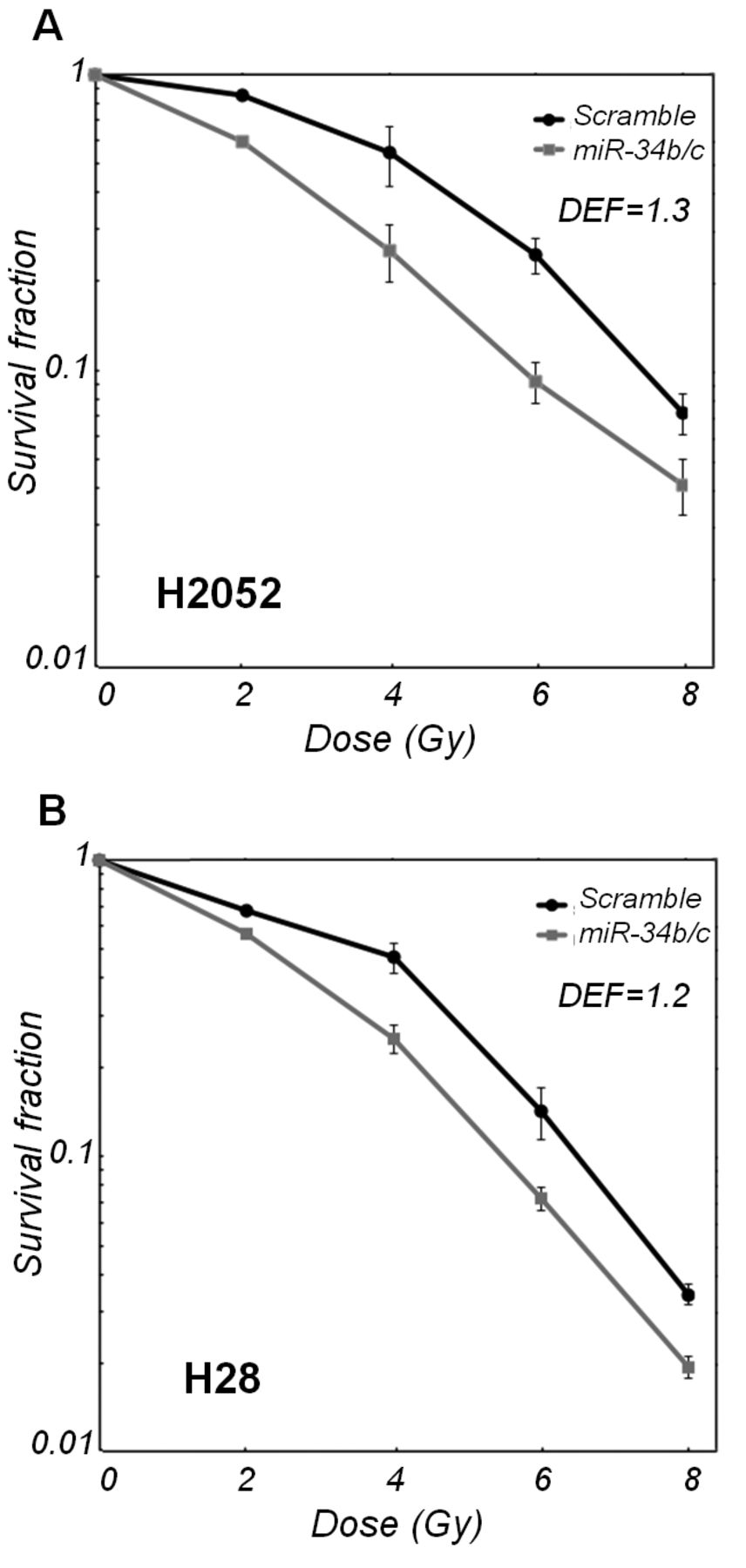
Effect of miR-34b/c on cellular radiosensitivity in malignant pleural mesothelioma (MPM) cells. miR-34b/c-transfected and scramble RNA-transfected MPM cells, H2052 (A) and H28 (B), were irradiated with graded doses of X-rays. The colony-forming efficiency was determined at 14 days after irradiation, and survival curves were generated. Data are the mean±SD of two independent experiments. DEF, Dose-enhancement factor.
Effect of miR-34b/c on radiation-induced γH2AX foci. We examined the DNA damage repair potential using a γH2AX foci assay. Representative micrographs are shown in Figure 2A. Compared with the scramble RNA transfectant, the number of γH2AX foci in miR-34b/c-transfected H2052 was significantly lower at 1 and 6 hours (considered to be early-phase) after IR but was higher at 24 hours (considered to be late-phase) (Figure 2B). We also calculated the rate of reduced γH2AX foci at 6 hours and 24 hours compared with 1 hour, i.e. the named reduced DSB rate (NDR). The formula to calculate the NDR is as followed: (γH2AX foci per cell at 1 hour-γH2AX foci per cell at 24 hours)/γH2AX foci per cell at 1 hour. NDR was lower in miR-34b/c transfectants (24.0%) than that in scramble transfectants (73.8%) at 24 h after IR (Figure 2C). This result suggests that DNA DSB repair potential was impaired in miR-34b/c transfectants compared with the scramble transfectants.

Effect of miR-34b/c on radiation-induced phosphorylated histone H2AX (γH2AX) foci in H2052 cells. A: Representative micrographs obtained from scramble RNA-transfected H2052 cells (left) and miR-34b/c-transfected H2052 cells (right) at 1 h after irradiation (4 Gy). B: miR-34b/c-transfected and scramble RNA-transfected H2052 cells on growing chamber slides were exposed to irradiation (4 Gy) and fixed at a specific time for use in immunocytochemical analyses of the nuclear γH2AX foci. Foci were evaluated in 50 nuclei per treatment for each sample. Data are the mean±SEM. *p<0.05. C: Reduced double-strand break (DSB) rate in miR-34b/c-transfected and scramble RNA-transfected H2052 cells at 1, 6 and 24 h after 4 Gy irradiation. Data are the mean+SEM of two independent experiments.

Influence of miR-34b/c on the cell-cycle distribution. miR-34b/c-transfected and scramble RNA-transfected H2052 cells were irradiated at 4 Gy. The cells were collected at 24 h after irradiation. The cell-cycle distribution was determined using flow cytometry according to an analysis of propidium iodide-stained cells. Data shown are representative of two independent experiments.
miR-34b/c induced G1 arrest and apoptosis. To further investigate the mechanism responsible for the radiosensitization effect of miR-34b/c, we performed a cell-cycle analysis on H2052 cells. The percentage of sub-G1 and G1 phase miR-34b/c transfectants was increased by 7.8% in non-irradiated H2052 cells and by 5.5% in 4-Gy-irradiated H2052 cells at 24 hours after IR (Figure 3). Focusing on the apoptotic fraction, the sub-G1 phase was increased by 9.6% in miR-34b/c-transfected H2052 cells at 24 hours after IR, suggesting that miR-34b/c enhanced radiation-induced apoptosis.
Impact of miR-34b/c on primary target proteins and apoptosis-related proteins. To examine the effect of miR-34b/c on its target proteins and apoptosis-related proteins, we focused on CCND1, CDK4/6, and BCL2 as targets of miR-34b/c and BAX, cPARP, pro-caspase 3, and cleaved caspase-3 for detecting apoptosis. The expressions of CCND1, CDK4/6 and BCL2 in miR-34b/c transfected H2052 cells were down-regulated at almost all the time points after IR that were examined (Figure 4). The expression of pro-caspase-3 was decreased in miR-34b/c transfectants. In contrast, the expressions of cleaved Caspase-3 and cPARP was increased in miR-34b/c transfectants and gradually increased after IR. These results suggested that miR-34b/c suppressed its target proteins and that IR enhanced apoptosis of miR-34b/c transfectants.

Effect of miR-34b/c on primary target proteins and apoptosis-related proteins. miR-34b/c-transfected and scramble RNA-transfected H2052 cells were irradiated (4 Gy) and harvested at the specified times. The putative target proteins of miR-34b/c, namely (cyclin-D1, CCND1), cyclin dependent kinase-4 (CDK4), cyclin dependent kinase-6 (CDK6), and B-cell lymphoma-2 (BCL2) and apoptosis-related proteins BCL2 Associated X-protein (BAX), cleaved poly (ADP-ribose) polymerase (cPARP), procaspase-3, and cleaved caspase-3 were examined. Each blot is representative of two independent experiments. IR: Irradiation.
Discussion
In this study, a clonogenic survival assay revealed that the restoration of miR-34b/c enhanced the radiosensitivity of MPM cells. This result is the essential finding of the present study. To understand the mechanism of radiosensitization, we focused on DNA DSB repair, cell-cycle distribution, and expression of the target proteins of miR-34b/c and apoptosis-related proteins.
Firstly, we estimated the DNA DSB status by counting γH2AX foci. The number of γH2AX foci after IR was lower at the early-phase but higher at the late-phase and the NDR was low in miR-34b/c transfectants compared with the scramble transfectants. As translation of this result, two possibilities should be considered: i) DNA DSB repair was impaired in miR-34b/c transfectants in the early-phase and DSBs accumulated at the late-phase, ii) miR-34b/c transfectants contained a low amount of DNA because miR-34b/c itself influenced the cell cycle, resulting in a low rate of cells entering the S phase. To understand how miR-34b/c impaired DNA DSB repair, CCND1, a putative target of miR-34b/c, seems to play a crucial role. Reportedly, CCND1 binds directly to RAD51 and is recruited to sites of DNA damage, and a reduction in CCND1 impairs the recruitment of RAD51 to damaged DNA and increases the cellular radiosensitivity (10). We confirmed that miR-34b/c suppresses CCND1 with or without IR and that the suppression was greater in miR-34b/c transfectants at 24 hours after IR. Accordingly, miR-34b/c is assumed to impair DNA DSB repair by suppressing CCND1. This is considered to be a part of the mechanism by which miR-34b/c enhances radiosensitivity.
Secondly, we estimated the cell-cycle distribution and related proteins. We observed that the proportion of sub-G1 phase miR-34b/c transfectants at 24 hours after IR was significantly greater than that for scramble transfectants, indicating that radiation-induced apoptosis took place in miR-34b/c transfectants. Of note, BCL2 expression was inhibited and that of cPARP and cleaved caspase-3 were increased in miR-34b/c transfectants after IR. As other cell-cycle-related targets of miR-34b/c, CCND1, CCNE2, CDK4 and CDK6 are known as key molecules for the G1 checkpoint (4, 6, 14), and some of these targets were confirmed to be down-regulated in miR-34b/c transfectants in the present study. Indeed, G1 arrest occurred in miR-34b/c transfectants. However, a question arises here. Generally cells in the G1 phase are considered to be relatively radioresistant compared with those in other phases (15). There are several reports that explain this contradiction. Kodym et al. reported that flavopiridol, a pan-CDK inhibitor, was reported to increase the radiosensitivity of both proliferating and quiescent lung cancer cells by suppressing DNA DSB repair thorough a cell cycle-unrelated mechanism (11). Other studies have demonstrated CDK inhibitor-mediated G1 arrest and radiation-induced apoptosis in breast cancer (16), as well as colonic and gastric cancer (12). We observed that miR-34b/c suppressed CDK4/6, which indicates that miR-34b/c has similar effects to a CDK inhibitor. Considering these, the effects of miR-34b/c on impairing DNA DSB repair and suppressing cell-cycle-related proteins resulted in an enhancement of radiosensitivity of MPM cells.
In conclusion, we revealed that the restoration of miR-34b/c in MPM cells enhanced radiosensitivity and promoted radiation-induced apoptosis, indicating that miR-34b/c, used in combination with IR, may be a potential therapeutic target for MPM.
Acknowledgements
We greatly acknowledge the grant support from the 13 fields of occupational injuries and illnesses of the Japan Labor Health and Welfare Organization (T. Kishimoto). We thank Mr. Seiji Tabara and Mr. Hirofumi Uno (Department of Radiology, Okayama University Hospital) for irradiating the MPM cells. We also thank Professor Adi F. Gazdar (Hammon Center for Therapeutic Oncology Research, University of Texas Southwestern Medical Center, Dallas, TX, USA) for the kind gift of cell lines.
- Received August 18, 2012.
- Revision received October 8, 2012.
- Accepted October 9, 2012.
- Copyright© 2012 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.