


Abstract
Aim: Our previous study showed that methylation of the mutL homolog 1 (MLH1) promoter may spread upstream from the Alu elements in intron 1. In this study, we investigated if the Alu methylation could also spread downstream. Materials and Methods: Two colorectal cancer cell lines (RKO, SW48), and four colorectal and three gastric carcinomas [all Microsatellite Instability (MSI)-positive] were selected as cases. Normal colorectal and gastric mucosa and human peripheral blood, and one MSI-negative colorectal cancer case served as controls. After extraction of DNA, bisulfite genomic sequencing was used to analyze the methylation level of exon 2 and the adjacent part of intron 2 of the MLH1 gene. Results: Exon 2 and the partial intron 2 exhibited a high level of methylation in controls. In contrast, demethylation in these regions was seen in gastrointestinal cancer. Conclusion: Exon 2 methylation is not likely to influence MLH1 gene expression. It seems that de-methylation of exon 2 and intron 2, in combination with intron 1, is associated with methylation spreading of the MLH1 promoter. The region around exon 2 could possibly bind proteins that regulate methylation, and therefore affect gene expression.
MutL homolog 1 (MLH1) is a key gene for DNA mismatch repair, which is involved in maintaining the stability of the genome. Loss of function of MLH1 may cause a high rate of gene mutations in the genome, and result in carcinogenesis. Promoter methylation of MLH1 is a frequent event in cancer, resulting in loss of MLH1 expression (1-3). However, the mechanism of MLH1 methylation is relatively unknown.
Alu elements were found to be hypermethylated in normal cells (4). Hence, Alu elements are known as methylation centres in the genome (5). Our previous study showed that MLH1 promoter methylation may spread from adjacent intron 1 Alu elements (6). Our results suggest that methylation of Alu elements in intron 1 may spread upstream to the promoter. Based on this, we hypothesized that Alu methylation could also spread downstream to the 3’ part of the MLH1 gene.
To test the hypothesis, we analysed the methylation status of all CpG sites of MLH1 exon 2 and adjacent partial intron 2 in colorectal cancer cells and tissues, gastric cancer tissues and normal tissues using bisulfite genomic sequencing.
Materials and Methods
Ethics statement. This research has been approved by the Review Board of Huazhong University of Science and Technology. We obtained tissue samples with written informed consent from the participants involved in the study. The Ethics Committee of Huazhong University of Science and Technology specifically approved the procedures.
Normal and cancerous samples. Samples used in this study are the same as in our previous study (6). Two colorectal cancer cell lines, RKO and SW48, with MLH1 promoter methylation (2) were ordered from the Chinese Academy of Sciences (Shanghai, China). Five colorectal and three gastric cancer tissues were obtained from Tongji Hospital (Wuhan, China). The normal colorectal and gastric mucosa and the peripheral blood of a healthy individual were also obtained from Tongji Hospital.
Cell culture and DNA isolation. Cells were grown in Dulbecco's modification of Eagle's medium (DMEM) supplemented with 10% foetal bovine serum at 37°C with 5% CO2. DNA was extracted from harvested cells, cancer tissues, normal mucosa and blood samples using a DNA isolation kit (Sangon, Shanghai, China) and TIANamp genomic DNA kit (Tiangen, Beijing, China).

The bisulfite Polymerase Chain Reaction. Lane 1, DNA marker; lane 2: sample; lane 3, water.
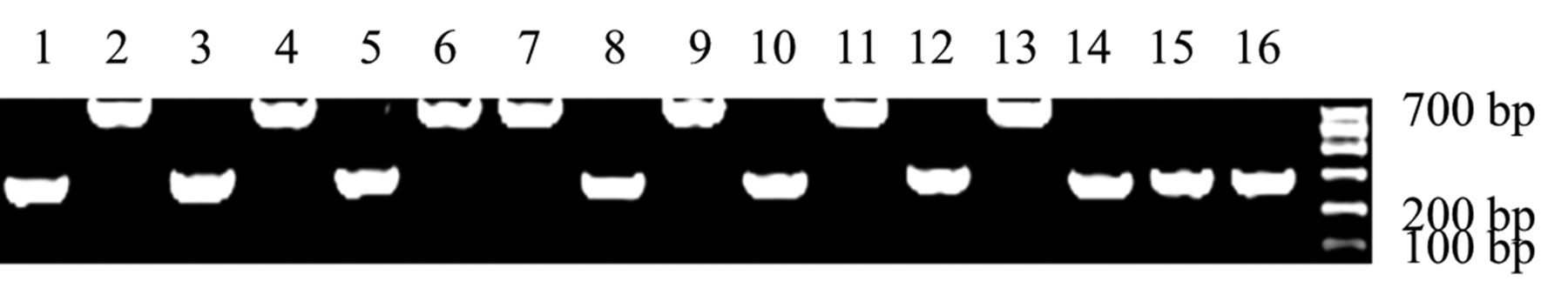
The colony Polymerase Chain Reaction. Lanes 1, 3, 5, 8, 10, 12, 14, 15 and 16 show the 250 bp PCR products, indicating empty plasmids. Lanes 2, 4, 6, 7, 9, 11 and 13 display 483 bp of products, suggesting successful cloning. Lane 17: DNA marker.
Oligonucleotide sequences of the primers used for bisulfite genomic sequencing.
Bisulfite genomic sequencing. The extracted DNA was converted using the EZ DNA Methylation Kit (Zymo Research Corporation, Orange, CA, USA). The methylation status of MLH1 exon 2 and partial intron 2 was determined using bisulfite genomic sequencing. In total there were five Polymerase Chain Reaction (PCR) amplicons designed to cover the region from IVS1-149 to IVS2+2067, relative to the translation start site. The primers used are shown in Table I. PCR was performed at 95°C for 5 min followed by 40 cycles of 95°C for 30 s, 55-61°C for 45 s and 72°C for 1 min with a final extension at 72°C for 7 min. A hot start was used by adding the enzyme during the first cycle at about 72°C, after a pre-incubation time of 5 min at 95°C. The PCR products were tested in 2% agarose gels (Figure 1) and then cloned into pEASY-T1 vector (TransGen Biotech, Beijing, China). Colony PCR was undertaken to screen the positive colonies (Figure 2). The clones with the right sizes of PCR products were sequenced on an ABI sequencer with dye terminators (Applied Biosystems, Foster City, CA, USA). With sequencing results of five clones, the methylation frequency was determined for each CpG site (Figure 3).
Results
One Alu element was identified in intron 2, by searching a database of human Alu repeat elements (Figure. 4). A total of 24 CpG sites located on exon 2 and intron 2 were measured for methylation status using bisulfite genomic sequencing (Figure 4). The two colorectal cancer cell lines, four MSI-positve colorectal cancer tissues, and three MSI-positive gastric cancer tissues exhibited different patterns compared to the normal colorectal and gastric mucosa and peripheral blood, and the MSI-negative colorectal cancer tissue (Figure 4). The normal colorectal and gastric mucosa and peripheral blood, and MSI-negative colorectal cancer tissue displayed hypermethylation in exon 2, also showing a high level of methylation in intron 2 except the normal gastric mucosa. In contrast, for the colorectal cancer cells and tissues (MSI-positive) and gastric cancer tissues, exon 2 and the regions between exon 2 and Alu element were hypomethylated or de-methylated. There was no significant difference between the methylation levels of intron 2 Alu element of cases and controls.
Discussion
Methylation of the promoter regions has been detected in many genes. However, very few studies have focused on the downstream regions, e.g. exon 1 and exon 2. Here exon 2 and partial intron 2 of MLH1 gene were screened for their methylation status. Our results showed that both were hypermethylated in controls, suggesting that methylation of exon 2 is not involved in deregulation of MLH1 gene expression (Figure 4). Previous studies reported that methylation of exon 2 or exon 3 in some genes, e.g., hepaCAM and POMC affects their expression (7,8). Another study showed that exon 2 methylation in the SOCS-1 gene does not influence gene expression (9). However, exon 1 exhibited hypermethylation in cases and demethylation in controls (Figure 4). Therefore, it is clear that exon 1 and exon 2 methylation play different roles in MLH1 gene expression.
Furthermore, exon 2 and intron 2 were examined for their methylation level in cases, and hypomethylation or demethylation was identified (Figure 4). In combination with our previous data, the region around exon 2 was hypomethylated or de-methylated [(6) and Figure 4]. It is considered that protein binding specifies sites of DNA de-methylation (10). It can be concluded that proteins binding to the region around exon 2 could play a role in demethylation of the regions, which may trigger methylation spreading to the MLH1 promoter region. Further studies are needed to isolate the binding proteins and analyze their function in methylation spreading.
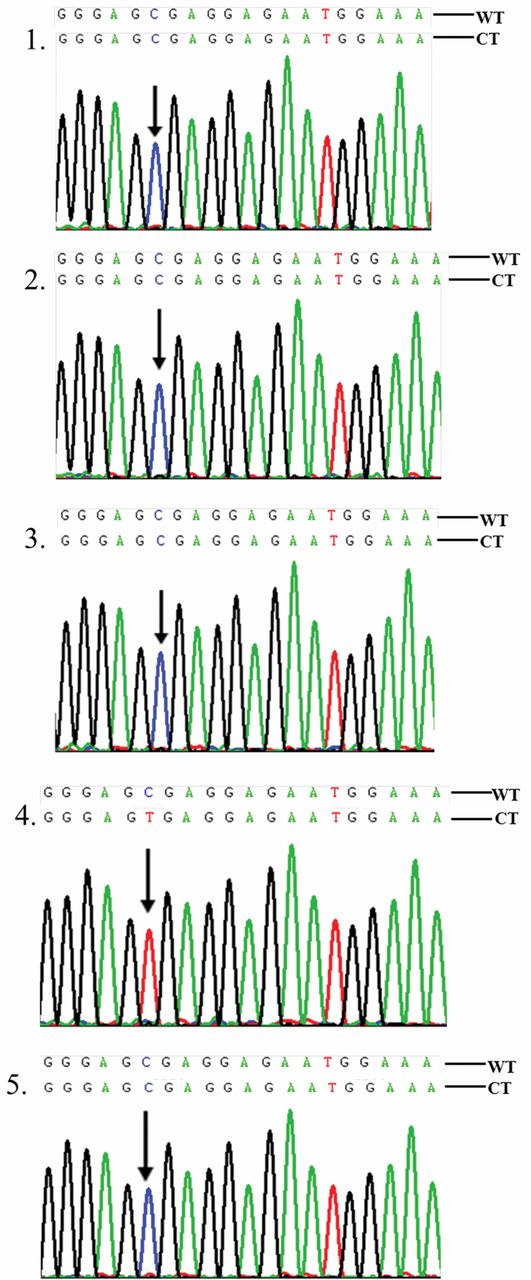
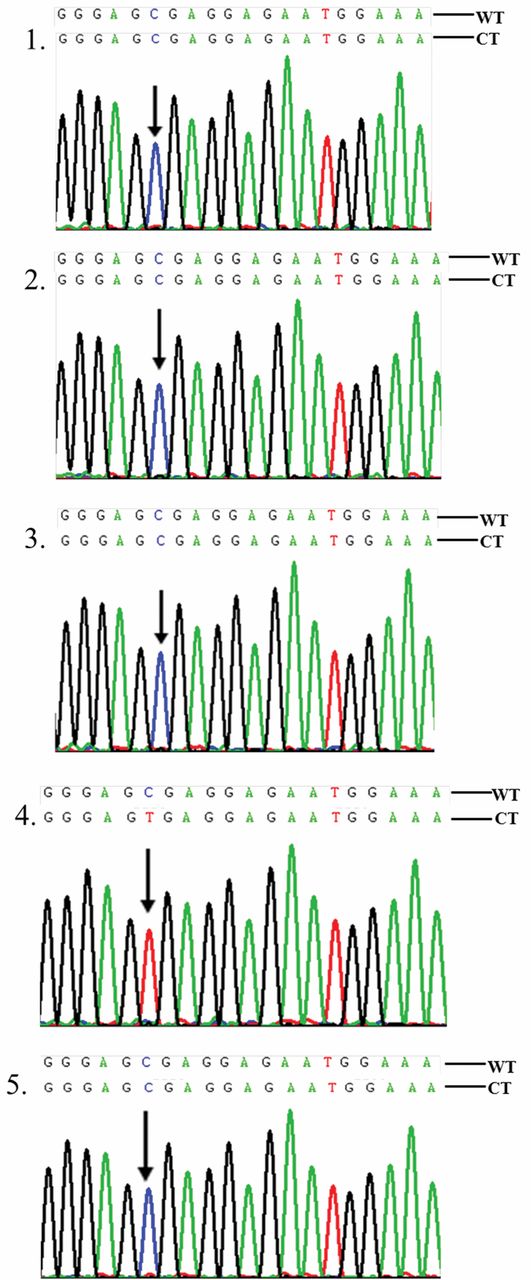
Partial sequencing data for five clones of an Polymerase Chain Reaction amplicon located between IVS2+1724 to IVS2+2067 of mutL homolog 1 (MLH1) in colorectal cancer C35T cell. The CpG sites are underscored. Clones 1, 2, 3 and 5 display methylation, and clone 4 no methylation. The methylation frequency of the CpG site is 80%. WT, Wild type; CT, converted type. The arrows indicate the C/T conversion sites.
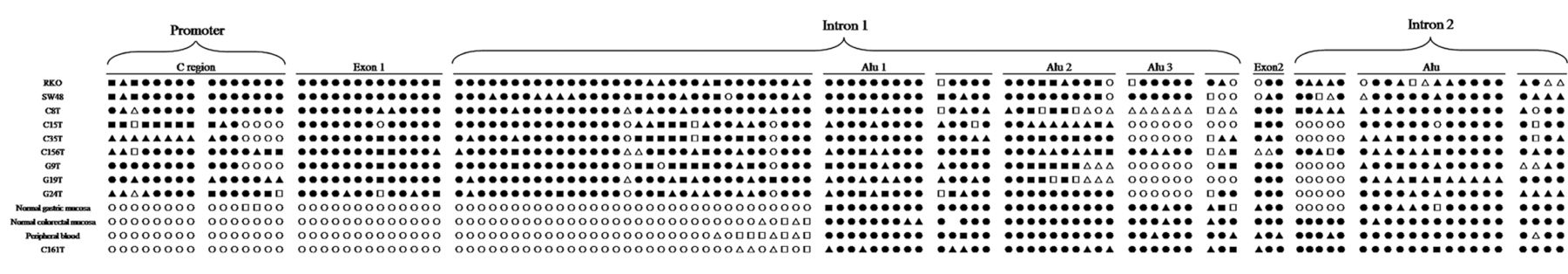
Methylation status analysis of CpG sites of the mutL homolog 1 (MLH1) C region containing promoter, exon1, intron 1, exon 2 and partial intron 2 in normal colorectal and gastric mucosa and peripheral blood, colorectal cancer cells and tissues, and gastric cancer tissues. In total 117 CpG sites were analyzed. ●, ▴, ▪, □, ▵ and ○ represent methylation frequency of 100%, 80%, 60%, 40%, 20% and 0, respectively. RKO and SW48: colorectal cancer cells; C8T, C15T, C35T, C156T: Microsatellite Instability (MSI)-positive colorectal cancer; C161T: MSI-negative colorectal cancer; and G9T, G19T and G24T: MSI-positive gastric cancer.
The normal gastric mucosa also displayed demethylation in the region between exon 2 and the Alu element of intron 2. This suggests a tissue-specific difference in methylation at this region between normal gastric and normal colorectal mucosa. It also suggests that demethylation that had taken place in the normal gastric mucosa cells, however, had not caused methylation spreading to the promoter. Methylation spreading is probably triggered by additional events, and cells could subsequently turn into precancerous cells or cancer cells.
In conclusion, methylation of exon 2 may not reduce MLH1 gene expression. However, demethylation of exon 2 and intron 2, in combination with intron 1, could be associated with methylation spreading of the MLH1 promoter, which subsequently results in decreased or negative expression of MLH1 gene.
Footnotes
-
↵* These Authors contributed equally to this work.
-
This project was supported by the Huazhong University of Science and Technology (2010MS034).
- Received August 8, 2012.
- Revision received October 5, 2012.
- Accepted October 8, 2012.
- Copyright© 2012 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.