


Abstract
Metastasis is a major cause of death of patients with malignant tumors. Matrix metalloproteinases (MMPs) are important for the migration and invasion of various types of cancer cell. Propofol is a known anesthetic agent, widely used for short-term anesthesia and for longer-term sedation. Propofol inhibits the proliferation of a variety of tumor cells, but there is no available information regarding propofol-inhibited migration and invasion of tumor cells in vitro. In this study, we investigated the effects of propofol on the migration and invasion of human lung carcinoma A549 cells. Wound healing assay and Boyden chamber assays indicated that propofol inhibited the migration and invasion of A549 cells in vitro. Gelatin zymographic analysis showed the inhibitory effect of propofol on the activation of expression MMP-2. Western blot analysis also indicated that propofol suppressed the protein expiration of growth factor receptor-bound protein 2 (GRB2), Jun N-terminal kinases 1/2 (p-JNK1/2), p-p38, MMP-2 and MMP-9 in A549 cells. Results from real-time PCR assay also showed that propofol inhibited the mRNA gene expression of MMP-2, -7 and -9, and enhanced that of tissue inhibitor of metalloproteinase 1 (TIMP1) and TIMP2 in A549 cells. Taken together, these data show that propofol inhibits MMP-2 and -9 mRNA and protein expressions, resulting in suppression of lung cancer cell invasion and migration in vitro.
Lung cancer remains the most common fatal malignancy in the Western world (1-3) and is one of the leading causes of death worldwide (4). Based on the 2011 report from the Department of Health, R.O.C. (Taiwan), approximately 26 people per 100,000 die annually from lung cancer in Taiwan. Patients with lung cancer commonly present with advanced metastatic disease that if untreated leads to a mean survival of 4-5 months and 1-year survival rate of less than 10% (4, 5).
The invasive and metastatic capacity of tumor cells plays an important role in malignant tumor development (6, 7). It is well known that metastasis occurs through a series of sequential steps for cancer cells to invade adjacent tissues, intravasation, transport through the circulatory system, arrest at a secondary site, extravasations, and growth in a secondary organ (8). The most important steps in tumor metastasis are directly correlated with cell adhesion and detachment (1) and there are both regulated by cell-cell and cell-extracellular matrix (ECM) interactions which coordinate the invasive mechanism (9).
Currently, the treatment of lung cancer includes chemotherapy and radiotherapy, but the survival is still very poor when the disease is diagnosed at an advanced stage and better treatments are desperately needed. Much evidence indicates that new targets in cancer cells associated with metastasis are needed.

Propofol affects the percentage of viable A549 cells. Cells were placed in F12K medium + 10% fetal bovine serum (FBS) and treated with 0, 15, 20 and 25 μg/ml of propofol for 24 and 48 h. The cells from each treatment were collected and analyzed for cell viability by flow cytometry as described in the Materials and Methods. Each point is the mean±S.D. of three experiments.
Propofol (2,6-di-isopropylphenol) is a sedative and anesthetic agent, widely used in the intensive care units for short-term anesthesia and for longer-term sedation (10). Propofol is also being used with increasing frequency for pediatric and obstetric procedures requiring anesthesia or sedation. It was reported that propofol interacts with both γ-aminobutyric acid (GABAA) receptors and N-methyl-D-aspartate (NMDA) glutamate receptors (11, 12) and mediated the activation of GABAA receptors and the inhibition of NMDA receptors; both receptors are involved in the alterations in calcium fluxes through slow calcium ion channels (13).
Propofol has been shown to possess antioxidant properties both in vitro (14-17) and in vivo (18), and has the capability to scavenge hydrogen peroxide (H2O2) (19), to reduce the formation of lipid peroxides (20) and the expression of nitric oxide synthase (21), and to stabilize the mitochondrial membrane (22). When propofol was administered together with ketamine to infant mice, it caused neurodegeneration and subsequent neurocognitive disturbances (23). Furthermore, many reports have shown that propofol attenuated reperfusion injury in the cerebral cortex (24), kidney (25), cerebral parenchymal arterioles (26) and intestinal mucosa (27). It has been reported that propofol can induce apoptosis of human leukemia HL-60 cells (28) and murine leukemia RAW264.7 cells (29). A previous study demonstrated that propofol suppresses the invasion and metastatic actions of tumor cells through modulating Rho A signaling and that it exhibits an anti-invasive effect in vivo (30). However, the ability of propofol by itself to inhibit migration and invasion of human lung cancer cells has not been systematically evaluated or reported. Therefore, the present study investigated whether propofol inhibits migration and invasion of human lung cancer A549 cells.
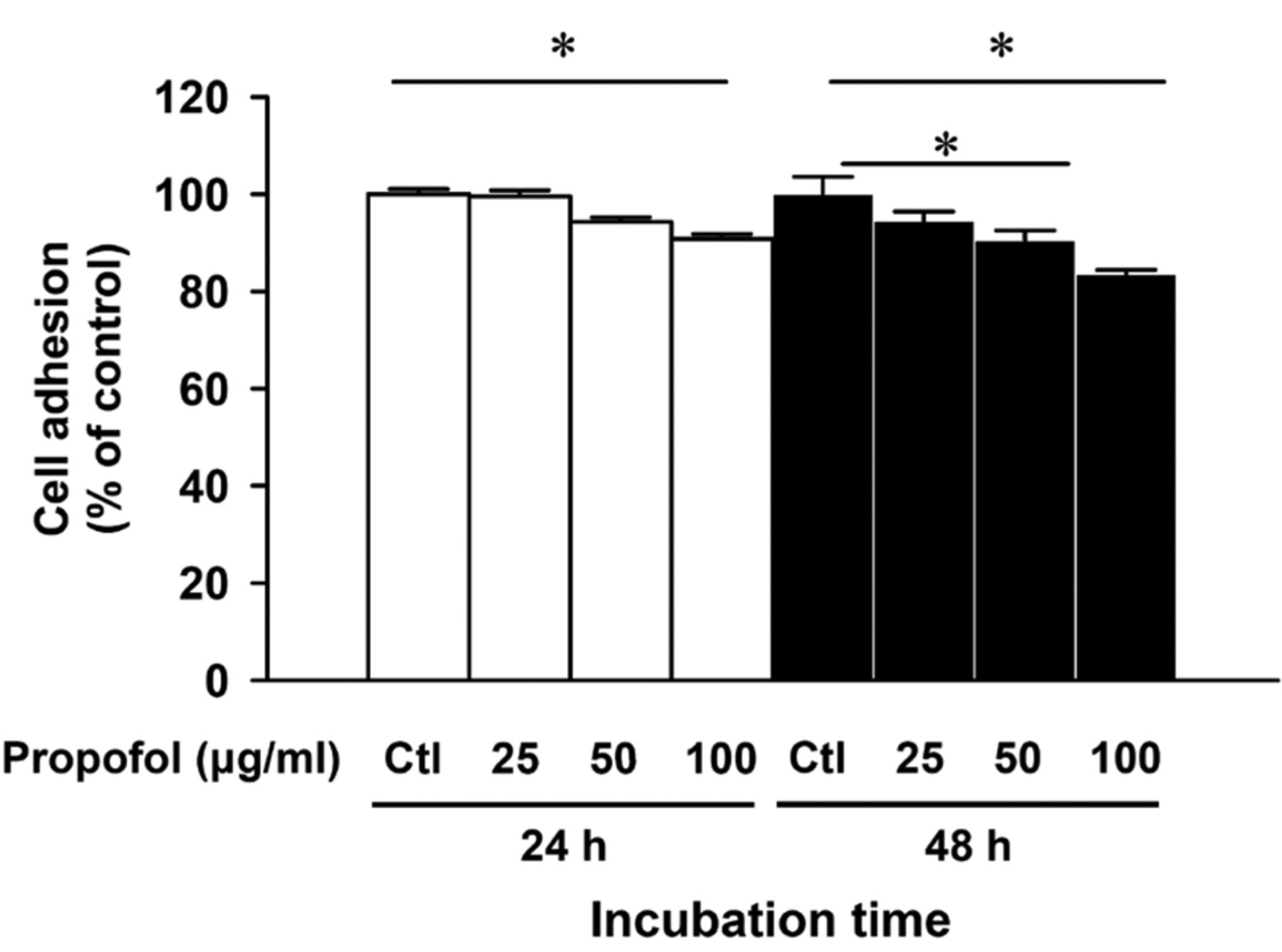
Propofol inhibits the adhesion of A549 cells. Cells were exposed to 25, 50 and 100 μg/ml of propofol for 24 and 48 h, and PBS as control sample (Ctl). The adhesion assay was performed on 24-well plates coated with collagen for 2-h incubation as described in the Materials and Methods. The results are presented as the percentage of the vehicle-treated control (Ctl). Each bar represents the mean±SD of three independent experiments. *p<0.05, statistically significant when compared with the untreated control.
Materials and Methods
Materials and reagents. Propofol was obtained from B. Braun Melsungen AG (Schwarzenberger Weg, Melsungen, Germany). Stock solution of propofol was prepared in phosphate buffered saline (PBS), and an equal volume of PBS (0.1%) was added to the controls. Propidium iodide (PI), crystal violet, Tween-20, formaldehyde and β-Actin were purchased from Sigma-Aldrich Corp. (St. Louis, MO, USA). F12K medium, L-glutamine, fetal bovine serum (FBS), penicillin-streptomycin and trypsin-EDTA were obtained from Invitrogen Life Technologies (Carlsbad, CA, USA). Anti-MMP-9 (Cat. AB19016), Millicell Hanging Cell Culture Inserts (Cat. PIEP12R48) and Immobilon-P transfer membrane (Cat. IPVH00010) were brought from Merck Millipore Corp. (Billerica, MA, USA). Primary antibodies to growth factor receptor-bound protein 2 (GRB2), phosphorylated c-Jun N-terminal kinases (p-JNK1/2), p-p38, matrix metalloproteinase-2 (MMP-2), MMP-9 and β-actin, and secondary antibodies were purchased from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA) and diluted in PBS Tween-20 before use. Matrigel was obtained from BD Biosciences (Franklin Lakes, NJ, USA).
Cell culture. Human lung carcinoma cell line (A549) was purchased from the Food Industry Research and Development Institute (Hsinchu, Taiwan, R.O.C.). Cells were placed onto 75 cm2 tissue culture flasks and maintained in F12K medium supplemented with 10% FBS, 100 units/ml penicillin, 100 ng/ml streptomycin and 2 mM L-glutamine, and grown at 37°C under 5% CO2 and 95% air humidified at one atmosphere (31).

Propofol affects the motility and migration of A549 cells. Cells were exposed to 15, 20 and 25 μg/ml of propofol for 12 and 24 h. Cell migration was examined by wound healing assay (A) and the migrated cells were quantitied (B) as described in the Materials and Methods. Each point is the mean±S.D. of three experiments. *p<0.05, statistically significant when compared with the untreated control.
Determination of percentage of viable cells by PI extrusion. A549 cells at a density of 2×105 cells/well were placed in 12-well plates and treated with 0, 15, 20 and 25 μg/ml of propofol for 24 and 48 h, respectively. Cells were harvested, stained with PI (4 μg/ml) and then immediately analyzed by flow cytometry (FACSCalibur; BD Biosciences, San Jose, CA, USA) as previously described (32, 33).
Cell adhesion assay. A549 cells at a density of 2×105 cells/well were placed in 12-well plates and then exposed to propofol at final concentrations of 0, 25, 50 and 100 μg/ml for 24 and 48 h. Cells were then harvested and seeded for 2 h onto 24-well plates which were coated with 150 μl type I collagen (10 μg/ml) (Merck Millipore, corp.) overnight. At the end of incubation, non-adherent cells were removed and washed with PBS, and adherent cells were fixed in 70% ethanol for 15 min. Cells were stained with 0.2% crystal violet for 10 min, and fixed cells were lysed in 0.2% Triton X-100 for 30 min. The lysed solution (150 μl) was added to each well of the 96-well ELISA plates, and then the absorbance at 540 nm was measured as previously described (34, 35).
Wound healing assay. A549 cells at a density of 1×106 cells/well were placed in 6-well plates for 24 h to form a confluent monolayer and then cells were wounded with a 200-μl pipette tip. All cells in each well were treated with or without 15, 20 and 25 μg/ml of propofol for 12 and 24 h. At the end of incubation, cells in each well were photographed under phase-contrast microscopy. The cell-free area of each treatment in the well was measured as previously described (36, 37). Cell migration was calculated as the percentage of the remaining cell-free area compared with the area of the initial wound.
Boyden chamber assay for migration assay. The chemotactic directional migration assay was used for the measurements of migration of A549 cells (37, 38). Briefly, a 24-well Transwell with 8 μM pore filter (Merck Millipore, corp.) were coated with 30 μg type I collagen (Merck Millipore) for 1 h. A549 cells (1×104 cells/0.4 ml) in F12K medium were placed in the upper chamber and treated with PBS (as a control) and with propofol (15, 20 and 25 μg/ml) before undergoing migration for 0, 12 and 24 h. Non-migrated cells were removed from the upper chamber by using a cotton swab and the filter from each treatment was individually stained with 2% crystal violet in 2% ethanol after fixing with 3.8% formaldehyde in PBS. Migrated cells adhering to the underside of the filter were examined, photographed and counted using a light microscope at ×200 magnification. Each treatment including the control condition was assayed twice and three independent experiments were performed as previously described (36, 38).
Boyden chamber assay for invasion assay. Matrigel-coated transwell cell culture chambers (insets of 8 μm pore size, Merck Millipore, corp.) were used for the measurements of invasion of A549 cells as previously described (35, 36). A549 cells were cultured for 24 h in serum-free-F12 medium, collected and re-suspended in serum-free medium. A549 cells (1×104 cells/well) were placed in the upper chamber of the transwell insert and treated with PBS (as a control) or propofol (15, 20 and 25 μg/ml). F12K medium containing 10% FBS was placed in the lower chamber. All samples were incubated for 24 and 48 h at 37°C in a humidified atmosphere with 95% air and 5% CO2. In the upper chamber, the non-invasive cells were removed by using a cotton swab and the invasive cells were fixed with 3.8% formaldehyde in PBS and stained with 2% crystal violet in 2% ethanol. Invasive cells on the lower surface of the filter which penetrated through the Matrigel then were counted and photographed using a light microscope at ×200 magnification as previously described (36, 38).


Propofol influences migration and invasion of A549 cells. Cells were treated with or without 15, 20 and 25 μg/ml of propofol for 24 and 48 h. A, B: Cell migration was examined by the Boyden chamber assay and type I collagen-coated transwell. C, D: Cell invasion was examined by using Matrigel-coated transwell cell culture chambers. Ability of migration (B) and invasion (D) of A549 cells were quantified by counting the number of cells that migrated or invaded the underside of the porous polycarbonate membrane under a phase-contrast microscope. Data represent the average of three experiments. *p<0.05, statistically significant when compared with the untreated control (Ctl).
Gelatin zymography assay. A549 cells at a density of 2×105 cells/well were placed in 12-well plates and then were incubated in serum-free F12K medium in the presence or absence of 15, 20 and 25 μg/ml of propofol for 24 and 48 h. The conditioned medium was then collected and proteins were separated by electrophoresis on 10% SDS-PAGE containing 0.18% gelatin (Sigma-Aldrich Corp.). After electrophoresis, the gels were soaked in 2.5% Triton X-100 in ddH2O twice for a total of 60 min at 25°C, and then were incubated in substrate buffer (50 mM Tris base, 0.2 M NaCl, 5 mM CaCl2 and 0.02% Brij 35 in distilled water, pH 8.0) at 37°C for 24 h. Bands corresponding to activity of MMP-2 were visualized by negative staining using 0.3% Coomassie blue (Sigma-Aldrich Corp.) in 50% methanol and 10% acetic acid as previously described (35, 38).
Western blotting analysis. A549 cells at a density of 2×106 cells/well were placed in 6-well plates and then were incubated with or without propofol (15, 20 and 25 μg/ml) for 24 and 48 h. Cells were collected from each treatment and were re-suspended in PRO-PREP™ protein extraction solution (iNtRON Biotechnology, Seongnam, Gyeonggi-Do, Korea) to extract the total protein fraction. Each sample was centrifuged at 13,000 ×g for 10 min at 4°C to remove cell debris and for collecting the supernatant. Bio-Rad protein assay kit (Hercules, CA, USA) using bovine serum albumin (BSA) as the standard was used to measure the total protein from each treatment (32, 36). Protein abundance of GRB2, p-JNK1/2, p-p38, MMP-2 and MMP-9 were measured by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and western blotting as previously described (32, 33, 39).
Real-time PCR assay. A549 cells (2×106 cells/well) maintained in 6-well plates were maintained with or without 25 μg/ml propofol for 12 and 24 h. The total RNA from each treatment was extracted by using the Qiagen RNeasy Mini Kit (Qiagen, inc, Valencia, CA, USA) as previously described (32, 40). RNA samples from each treatment were then individually reverse-transcribed with High Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Carlsbad, CA, USA). Quantitative PCR from each sample was carried out for amplifications using 2× SYBR Green PCR Master Mix (Applied Biosystems) and forward and reverse primers for each gene as shown in Table I. Each assay was run on an Applied Biosystems 7300 Real-time PCR system in triplicates and expression fold-changes were derived using the comparative threshold cycles (CT) method (38, 41).
Statistical analysis. All data are presented as means±S.D. of three experiments. Statistical comparisons were performed using the Student's t-test and differences were between propofol-treated and control groups considered significant at a p-value of less than 0.05

Propofol alters matrix metalloproteinase-2 (MMP-2) enzyme activity in A549 cells. Cells (2×106 cells/well) were incubated with 0, 15, 20 and 25 μg/ml of propofol for 24 and 48 h. The supernatant was harvested after treatment and protein separated by gelatin zymography as described in the Materials and Methods. The MMP-2 activity was quantitied using ImageJ software.

Propofol affects the level of proteins associated with migration and invasion of A549 cells. Cells (1×106 cells/well) were treated with 15, 20 and 25 μg/ml of propofol for 24 and 48 h. The total proteins were collected and the levels of growth factor receptor-bound protein 2 (GRB2), phospho-c-jun N-terminal kinase 1/2 (p-JNK1/2), p-p38, matrix metalloproteinase-2 (MMP-2) and matrix metalloproteinase-9 (MMP-9) were examined by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and western blotting as described in the Materials and Methods. Direct reprobing with anti-β-actin antibody was used as an internal control.
Results
Propofol affects the percentage of viable A549 cells. After A549 cells were treated with different concentrations of propofol for 24 and 48, cells were collected for PI staining to measure the cell viability. As shown in Figure 1, the percentage of viable A549 cells was reduced in cells treated with up to 25 μg/ml of propofol when compared to the untreated control group. These results indicated that propofol did not significantly reduce the percentage of viable A549 cells at the examined concentrations.
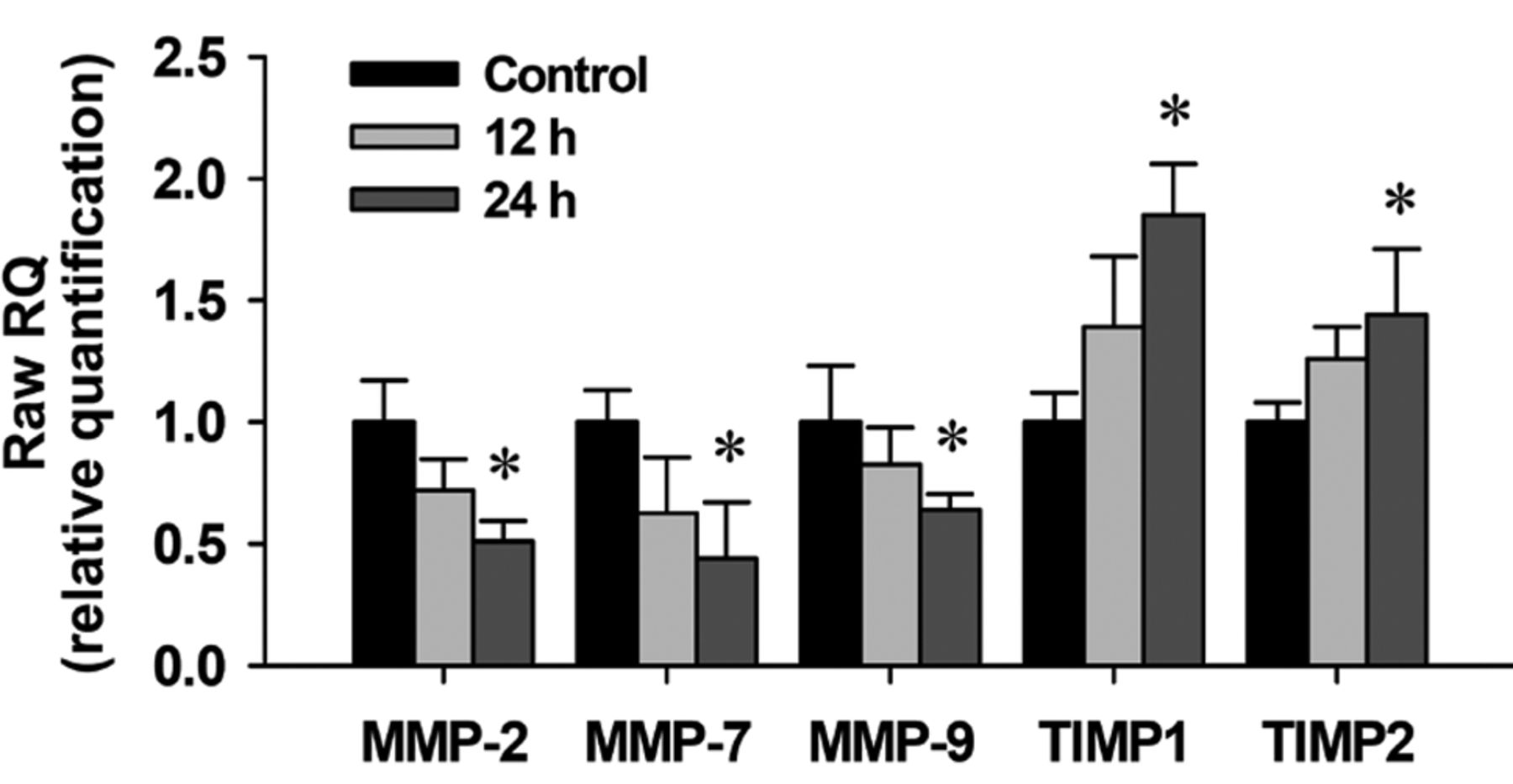
Propofol affects on matrix metalloproteinase-2 (MMP-2), MMP-7, MMP-9, tissue inhibitor of metalloproteinases 1 (TIMP1) and TIMP2 mRNA expression in A549 cells. The total RNA was extracted from A549 cells treated with propofol (20 and 40 μg/ml) for 12 and 24 h, and RNA samples were reverse-transcribed for real-time PCR as described in the Materials and Methods. The ratios of MMP-2, MMP-7 MMP-9, TIMP1 and TIMP2 mRNA expression to that of glyceraldehyde-3-phosphate dehydrogenase (GAPDH) are presented. Data represent the mean±S.D. of three experiments. *p<0.05 significantly different from the untreated control.
Propofol inhibits the adhesion of A549 cells. Cells were harvested and assayed for cell adhesion and the results are shown in Figure 2. A significant inhibitory effect on cell adhesion was observed in propofol-treated A549 cells at the concentration of 25, 50 and 100 μg/ml for 24 and 48 h exposure when compared with the untreated (control) cells. Importantly, these inhibitory effects of propofol on A549 cells are not due to a cytotoxic effect of propofol because the viability of these cells was barely affected by propofol at the examined concentration range.
Propofol affects the motility and migration of A549 cells. The wound closure assay was used to examine the effects of propofol on migration of A549 cells and the results are shown in Figure 3A, which indicates that relative wound closure in control cells, was higher than in the propofol-treated cells. Figure 3B indicates that the inhibition rates were 28% and 65% when cells were incubated with propofol at 20 and 25 μg/ml for 24 h, respectively. These effects indicated that propofol inhibited cell migration in a dose-dependent manner.

Schematic presentation of the mechanism of propofol-suppressed migration and invasion in A549 human lung adenocarcinoma epithelial cells.
The DNA sequence was evaluated using the Primer Express software and each assay was run on an Applied Biosystems 7300 Real-time PCR system.
Propofol influences migration and invasion of A549 cells. The effects of propofol on migration and invasion of A549 cells in vitro were investigated by using Boyden chamber assay and the results are shown in Figure 4. Propofol had a significant inhibitory effect on cell migration at concentrations between 15-25 μg/ml. Figure 4B shows that the migration inhibition was 28-37% and 54-77% when cells were incubated with propofol for 24 and 48 h, respectively. The invasion assay indicated that A549 cells moved from the upper chamber to the lower chamber in the absence of propofol (control group). However, the penetration of the EHS-coated filter by A549 cells was inhibited in the presence of propofol. The percentage inhibition at 15-25 μg/ml was 75-99% and 53-89% (Figure 4D) when cells were incubated with propofol for 24 and 48 h, respectively. Importantly, the inhibitory effects of propofol on the migration and invasion of A549 cells are not due to a cytotoxic effect of propofol.
Propofol reduces MMP-2 enzyme activity in A549 cells. It is well documented that MMP-2 plays an important role in the invasion of cancer cells (42, 43). Thus, we examined whether propofol inhibits the secretion of MMP-2 in A549 cells. Cells were exposed to propofol (0, 15, 20 and 25 μg/ml) for 24 and 48 h, and then the conditioned medium was harvested for examining the MMP-2 activity using gelatin zymography. As shown in Figure 5, propofol inhibited MMP-2 activity in A549 cells.
Propofol alters the levels of proteins associated with migration and invasion in A549 cells. To examine whether or not propofol inhibits the migration and invasion of A549 cells through the inhibition of associated proteins, we examined the effects of propofol using western blotting. Our results presented in Figure 6, showed that propofol reduced the protein levels of GRB2, p-JNK1/2, p-p38, MMP-2 and MMP-9 in A549 cells, which may lead to the inhibition of migration and invasion of A549 cells in vitro.
Propofol reduces the mRNA expression of MMP-2, MMP-7 and MMP-9, and increases that of TIMP1 and TIMP2 in A549 cells. It is well known that MMPs and tissue inhibitor of metalloproteinases (TIMPs) play a critical role in tumor invasion and metastasis (43-45). A549 cells were treated with propofol (25 μg/ml) for 12 and 24 h and then cells were harvested for isolation of total RNA to examine MMP-2, MMP-7, MMP-9, TIMP1 and TIMP2 mRNA expression by real-time PCR. Results are shown in Figure 7. The mRNA expression levels of MMP-2, MMP-7 and MMP-9 were reduced by propofol after a 24-h treatment. However, propofol increased the mRNA levels of TIMP1 and TIMP2 in propofol-treated A549 cells.
Discussion
In this study, we investigated the inhibitory effects of propofol on the migration and invasion of human lung cancer A549 cells in vitro. Several studies have shown that propofol induced cytotoxic effects in human leukemia HL-60 cells (28) and murine leukemia cells (29). However, there is no information on how propofol affects migration and invasion of cancer cells. Herein, we demonstrated that propofol significantly inhibited the migration and invasion of human lung cancer A549 cells and inhibited the MMP-2 activity at low concentrations (15-25 μg/ml).
Multiple molecular mechanisms regulate invasion and migration of cancer cells. The interaction of cancer cells with the ECM is essential for metastasis which is the principal cause of death of cancer patients (46). MMPs are involved in tumor angiogenesis, metastasis and stimulation of growth factor release from the ECM (44). Increased expression of MMPs is involved in tumor invasion and metastasis (47). Inhibition of MMP expression or MMP activity have been recognized as early targets in preventing cancer metastasis (43-45). Both MMP-2 and -9 are the key enzymes that are involved in degrading type-I and -IV collagens and ECM (43, 45). In particular, both MMP-2 and -9 have been reported to be related to the invasive metastatic potential of tumor cells (42).
In human ovarian cancer cells, MMP-2 plays a key role in this process, and its activity is regulated by complex protein-protein interactions at the posttranslational level (42, 43). Tumor metastasis involves tumor cell attachment, invasion, migration, vessel formation and cell proliferation, and these steps are regulated by an extremely complex mechanism (48, 49). Thus, several agents which can inhibit or block metastasis of cancer cells have been investigated (50). However, the efficacy of available agents remains unsatisfactory.
Propofol is clinically used for patients, but its effects on cancer cell migration and invasion have not been so far reported. Here, we demonstrated that propofol inhibited the protein expression of MMP-2 and MMP-9 in A549 cells (Figure 6). MMP-2 is constitutively overexpressed in highly metastatic tumors (51), whereas MMP-9 can be stimulated by cytokine through activation of nuclear factor-κB (NF-κB) and activator protein-1 (AP-1) (52, 53). Several reports have indicated that activation of PKC can cause translocation of the protein to membranes affecting the expression of MMP-9 via modulation of transcription factors such as NF-κB through the MAPK and PI3K signaling pathways. Propofol inhibited the PKC activity, such as JNK and p38 (Figure 6). Therefore, the downstream regulation of MAPK (p38 and JNK) pathway, may be involved in the propofol-suppressed MMP-2 and -9 expression and invasion of A549 cells. However, we did not demonstrate any significant MMP-9 activity in the A549 lung cancer cell line, suggesting a more important role for MMP-2 in this model. Moreover, propofol up-regulated the levels of TIMP1 and TIMP2 in A549 cells (Figure 6).
In conclusion, our findings suggest that propofol has multiple anti-metastatic activities in A549 cells. The possible signaling pathways of propofol-inhibited migration and invasion in A549 cells may be via blocking MMP-2 and -9 mRNA expressions, leading to in inhibition of MMP-2 and MMP-9 and stimulation of TIMP1 and TIMP2, as summarized in Figure 8.
Acknowledgements
This work was supported by the grant CMU100-ASIA-4 from China Medical University, Taichung, Taiwan.
Footnotes
-
↵* Both Authors contributed equally to this work.
- Received August 1, 2012.
- Revision received October 11, 2012.
- Accepted October 12, 2012.
- Copyright© 2012 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Effect of Propofol and Etomidate on the Proliferation, Cell-cycle Distribution, Apoptosis and Necrosis of Pancreatic Tumour Cells
- Effects of the Hypnotic Alkylphenol Derivative Propofol on Breast Cancer Progression. A Focus on Preclinical and Clinical Studies
- Etomidate Suppresses Invasion and Migration of Human A549 Lung Adenocarcinoma Cells
- Demethoxycurcumin Suppresses Migration and Invasion of Human Cervical Cancer HeLa Cells via Inhibition of NF-{kappa}B Pathways
- Cantharidin Impairs Cell Migration and Invasion of Human Lung Cancer NCI-H460 Cells via UPA and MAPK Signaling Pathways