


Abstract
In the present investigation, we report on the possible underlying mechanism for the cytotoxicity of compounds: 3-(4-(2-(dimethylamino)ethoxy)-phenyl)-7-methoxy-4-phenyl coumarin 5, 3-(4-(2-(pyrrolidin-1-yl)ethoxy)phenyl)-7-methoxy-4-phenylcoumarin 6, and 3-(4-(2-(diethylamino) ethoxy)phenyl)-7-methoxy-4-methylcoumarin 7 in the human lung (A549) cancer cell line, using Ray Biotech's Human Apoptosis Arrays and apoptotic protein antibodies. Apoptosis array results showed differential apoptotic proteins expression in the extracts of cells treated with compounds 5-7. Western blotting demonstrated that compound 5 induced apoptosis and caused cell death in the A549 cell line via an increase (up-regulation) in Bax protein expression (pro-apoptotic pathway) and a slight decrease (down-regulation) in Bcl-2 protein expression (anti-apoptotic pathway) after 6 h of treatment. These observations may provide valuable information on the mechanism by which these coumarin-based benzopyranone derivatives induce cytotoxicity in the human lung (A549) cancer cell line.
Coumarins are best-known as oxygen heterocylic compounds exhibiting diverse and interesting biological activities and in fact, have been used as therapeutic agents in the treatment of various diseases. Some therapeutic applications of coumarin and derivatives include photochemotherapy, anti-cancer and anti-HIV (1, 2), central nervous system [CNS] stimulants (3), anti-bacterials (4), anti-inflammatory (5) and anti-coagulants (6). For example, studies have shown that coumarin and derivatives demonstrate growth-inhibitory activities in both in vivo and in vitro human tumor cell lines such as A549 (lung), ACHN (renal), H727 (lung), MCF-7 (breast) and HL-60 (leukemia) (7, 8). Coumarin nucleus has emerged as a valuable molecular template for the development of different structural analogues with improved pharmacological profiles. Also, the pharmacological and biochemical properties, including the therapeutic applications of coumarin and derivatives depend on the pattern of substitutions of the coumarin core structure (9-11).
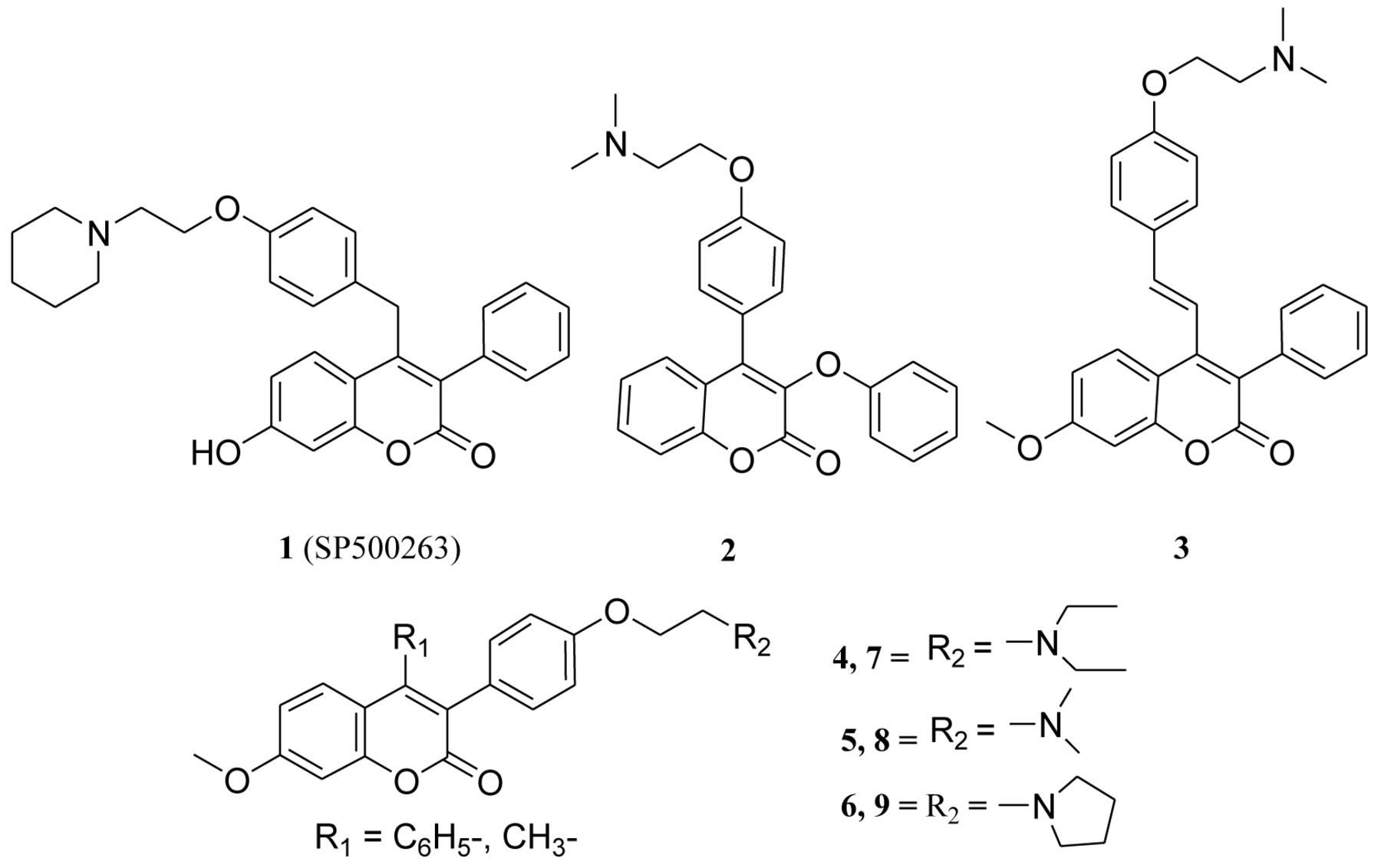
Studies have shown that attachment of the basic amino side chain (tertiary aminoalkoxy group) to the coumarin-based benzopyranone core structure has led to the identification of novel therapeutic agents (Figure 1), such as 4-(4-(2-(piperidin-1-yl)ethoxy)benzyl)-7-hydroxy-3-phenylcoumarin 1 (SP500263), 3-phenoxyl-4-[4-(2-N,N-dimethyl-amino)-ethoxy)-phenyl]coumarin 2 and 7-methoxy-3-phenyl-4-phenylvinyl coumarin 3 (12-14). On this basis, we recently developed several series of coumarin-based benzopyranone derivatives with different tertiary aminoalkoxy groups such as: diethylaminoethoxy, dimethylaminoethoxy, morpholinoethoxy, piperidinylethoxy and pyrrolidinylethoxyl groups (15-17). Our recent results revealed that the toxicity of 3-(4-(2-(diethylamino) ethoxy)phenyl)-7-methoxy-4-phenylcoumarin 4, 3-(4-(2-(dimethylamino)ethoxy)phenyl)-7-methoxy-4-phenyl coumarin 5, 3-(4-(2-(pyrrolidin-1-yl)ethoxy)phenyl)-7-methoxy-4-phenyl coumarin 6 (4-phenyl analogs) in the A549 cancerous cell line decreases significantly when the phenyl group at position 4 is replaced with a methyl group to afford compounds: 3-(4-(2-(diethylamino)ethoxy)phenyl)-7-methoxy-4-methyl coumarin 7, 3-(4-(2-(dimethylamino) ethoxy) phenyl)-7-methoxy-4-methylcoumarin 8, and 3-(4-(2-(pyrrolidin-1-yl)ethoxy)phenyl)-7-methoxy-4-methylcoumarin 9 (4-methyl analogs) (Figure 1). Furthermore, preliminary in vitro results obtained from the National Cancer Institute Developmental Therapeutics program have also revealed that compound 5 showed anti-proliferative activity against several cancer cell lines such as A549, EKVX, HOP-62, NCI-H226, NCI-H23, NCI-H322M, NCI-H460 and NCI-H522.
As part of our group's continued investigation of coumarin-based benzopyranone derivatives with basic amino side chain groups as an anticancer agent, we herein report the mechanism for cytotoxicity of compounds 5-7 in a human lung (A549) cancer cell line, using Ray Biotech's Human Apoptosis Arrays and apoptotic proteins (Bax and Bcl2) expression by western blot analysis.
Materials and Methods
Chemicals. F12K medium, penicillin-streptomycin anti-biotic solution (100×), fetal bovine serum (FBS), Trypsin-EDTA solution (1×), phosphate buffer (PBS), 50% glutaraldehyde and crystal violet were obtained from Sigma-Aldrich Company (St. Louis, MO, USA). The potassium phosphate, EDTA, Tris, boric acid, sodium chloride, SDS, Triton X-100, ethanol were obtained from Thomas Scientific Company (Swedesboro, NJ, USA). The primary antibodies (Bax and Bcl-2), secondary antibody conjugated to horseradish peroxidase (HRP), LumiGLO® chemiluminescent reagent and peroxide were obtained from Cell Signaling Technology, Inc. (Danvers, MA, USA). The human apoptosis arrays were obtained from Ray Biotech, Inc. (Norcross, GA, USA).
Preparation of samples for apoptosis array. Approximately 9.2×106 cells were plated in T-175 cm2 flasks in complete F12K medium and allowed to stabilize overnight. The cells were treated with compounds at 20 μM concentration. The flasks were incubated for 6 h at 37°C in a 5% CO2 incubator. After 6 h of incubation, the cells were trypsinized and pelleted by centrifuging at 2,500 rpm for 5 min. The cell pellet was suspended in 500 μl of 1× cell lysis buffer (from the Apoptosis Array Kit) and homogenized in a vial placed in ice for 15 s using a Polytron homogenizer. The homogenate was transferred to an eppendorff tube and centrifuged at 10,000 rpm for 10 min at 4°C to remove the lysed cell membrane debris. The supernatant was transferred to a fresh tube and the cell extract was stored at −80°C for use in the Apoptosis array.
Protein estimation. The protein concentration was determined using a BCA method kit (Pierce). Diluted albumin standards (BSA) and working reagent were prepared according to the kit. Different concentrations of each standard and each unknown sample (25 μl) were pipetted in triplicate wells of a 96-well plate and 200 μl of working reagent was added to each well. The plate was incubated at 37°C for 30 min and then read at 562 nm in a Bio-Tek plate reader. The protein concentration was measured from the standard curve.
Apoptosis array analysis. The apoptosis proteins expression was determined in the cell lysate using Ray Biotech's Human Apoptosis Array, (catalog # AAH-APO-G1), according to manual instructions with minor changes. The membranes were blocked for 1 h at room temperature, hybridized with 400 μg of protein cell lysate for 16 h at 4°C and then washed with buffer I (3×) and buffer II (2×) for 5 min. Following this, the membranes were incubated with biotin-conjugated primary antibodies for 2 h at room temperature, washed and then incubated with the HRP-conjugated streptavidin secondary antibodies at room temperature for 2 h. The arrays were washed again, developed by incubation with detection buffer for 5 min and then exposed to Kodak BioMax XAR X-ray film. The x-ray film was scanned using Cannon scanner and the densities of spots were analyzed by Un-Scan-It gel TM Program (Silk scientific, Inc., Orem, Utah, USA).
Bax and Bcl-2 expression in cell lysates. The cell pellet was suspended in 500 μl of 50 mM phosphate buffer with 0.1% Triton X-100 and sonicated for 15 s (3×) in a vial placed in ice. The tubes were centrifuged at 3000 rpm to remove cell membrane debris. The supernatant was transferred to the fresh tube and twenty-five microliters of sample were used to measure the protein concentration. Twenty micrograms of each protein sample in 1X SDS gel loading buffer was loaded into 10-15% SDS PAGE and electrotransfered to a nitrocellulose membrane. After transfer, the nitrocellulose membrane was washed with 25 ml of Tris buffered saline (TBS) buffer for 5 min, at room temperature. The membrane was incubated in 25 ml of blocking buffer for 1 h at room temperature, washed for 5 min each (3x) with 15 ml of 0.1% Tween 20 (TBS/T) and then incubated with a primary antibody (at the appropriate dilution) in 10 ml primary antibody dilution buffer with gentle agitation overnight at 4°C. Later, the membrane was again washed 5 min each (×3) with 15 ml of TBS/T, incubated with the appropriate HRP-conjugated secondary antibody (1:2000) in 10 ml of blocking buffer with gentle agitation for 1 h at room temperature and then washed for 5 min each (3×) with 15 ml of TBS/T. The membrane was then incubated with 10 ml LumiGLO® (0.5 ml 20X LumiGLO®, 0.5 ml 20X Peroxide and 9.0 ml Milli-Q water) with gentle agitation for 5 min at room temperature. The excess developing solution was drained from the membrane, wrapped in plastic wrap and exposed to x-ray film or to a ChemiDoc machine. The density of the bands on the blot was quantified using the Un-Scan-It get TM program (Silk scientific, Inc., Orem, Utah, USA).
Statistical analysis. Data for all treated cells were presented as percentage values in comparison to the untreated controls (100%). The data were analyzed for significance by one-way ANOVA, and then compared by Dunnett's multiple comparison tests, using the GraphPad Prism Software, version 3.00 (GraphPad Software, Inc., San Diego, CA, USA). Differences with the respective untreated control were considered statistically significant when p<0.05.
Results
Apoptosis array analysis. To explore the molecular mechanism associated with the cytotoxicity of compounds 5-7 in the human lung, A549 cancer cell line, we used RayBio® Human Apoptosis Antibody Arrays to investigate the effect of these compounds on 43 apoptotic proteins. We selected these compounds based on their toxicity and selectivity. In the present investigation, our results indicate that these compounds showed both up- and down-regulation (30% more or less expression than the control, untreated cells) in 10 proteins out of 43 apoptotic proteins on the array (Figure 2).
The differential protein expressions are as follows: compound 5 showed an up-regulation of TNF-alpha, IGF-BP-4, Bax, p53, caspase-3, IGF-1, caspase-8 and sTNF-R1 protein expression; compound 6 showed an up-regulation of IGF-BP-4, Bax, Bcl-w, caspase-3, IGF-1, caspase-8 and sTNF-R1 and down-regulation of CD40L protein expression; while compound 7 showed an up-regulation of p53, caspase-3, IGF-1, caspase-8 and sTNF-R1 proteins and a down-regulation of CD40L and TNF-alpha protein expression.

4-(4-(2-(piperidin-1-yl)ethoxy)benzyl)-7-hydroxy-3-phenylcoumarin 1 (SP500263), 3-phenoxyl-4-[4-(2-N,N-dimethylamino)ethoxy)-phenyl]coumarin 2, 7-methoxy-3-phenyl-4-phenyl-vinyl benzopyranone derivative 3, coumarin derivatives 4-6 (4-phenyl analogs) and 7-9 (4-methyl analogs).
Bax and Bcl-2 expression in cell lysates. To gain further insights into the molecular events associated with the induced cytotoxicity of compounds 5 and 6 (4-phenyl analogs) in the human lung cancer A549 cell line, the expression of Bax and Bcl-2 proteins was investigated using immunoblotting assay. In this investigation, the treated cell extracts of compounds 5 and 6 at concentrations of 20 and 40 μM (for 6 h) were blotted on the membranes and then hybridized with Bax and Bcl-2 antibodies (Cell Signaling). Our results (Figure 3) indicate that compound 5 induced apoptosis and caused cell death in the A549 cell line via (i) an increase (up-regulation) in Bax protein expression (pro-apoptotic pathway) and (ii) a slight decrease (down-regulation) of Bcl-2 proteins expression (anti-apoptotic pathway). However, compound 6 induced down-regulation of both Bax and Bcl-2 protein expressions. It was also observed that both protein expressions decreased with compound 5 and increased with compound 6 with increasing concentration in comparison to controls (untreated cells).
Discussion
Cancer is the leading cause of death in developed countries and also the second cause of death in developing countries (18). Lung cancer is the worldwide leading cause of death among cancer patients; out of which non-small cell lung cancer (NSCLC) accounts for 80% of total lung cancer cases (19). Even though, treatments against NSCLC have been developed; they often fail in the majority of patients due to drugs insusceptibility to advanced lung cancer stages (20). Therefore, the development of new cytotoxic drugs to improve the prognosis of lung cancer are urgently needed. The therapeutic strategy applied in the development of most anti-cancer agents is to kill cancer cells by triggering apoptosis in the cancer cells. Apoptosis, a major mechanism of cell death, is related to the regulation of developmental processes and homeostasis (21, 22). Our previous cytotoxicity results in the A549 cell line showed the LD50 values (lethal dose, concentration at which 50% of the cells die) of compounds 4 (7.08), 5 (5.0) and 6 (5.83) μM (previously reported) (15), and these of compounds 7 (78), 8 (>100) and 9 (>100) μM (unpublished data) after 48-h treatment. These results showed a significant decrease in toxicity as the result of the replacement of the 4-phenyl group (compounds 4-6) with the 4-methyl group (compounds 7-9) (Figure 1). Based on these results, we may assume that the phenyl ring plays a vital role in the observed cytotoxic activity of these compounds. Previous reported cell-cycle analysis also revealed that compound 6 showed selective increase of A549 cells in the G0/G1 phase in a concentration-dependent manner, thus suggesting apoptotic cell death (15). Based on these findings, we decided to explore whether these synthesized coumarin derivatives really induce cytotoxic activity by apoptosis mechanisms.

The effect of compounds 5-7 on apoptosis protein expression in human lung (A549) cancer cells using apoptosis array.
In this present investigation, apoptotic array results showed that compounds 5-7 exhibited differential expression of apoptotic proteins in the human lung cancer, A549, cell line (Figure 2). Hence, we may presume that this differential expression of proteins may be responsible for the observed toxicity of compounds (5 and 6) and less toxicity of compound 7 in human lung (A549) cancer cell line, as previously reported (15). Furthermore, compounds 5 and 6 (toxic compounds) showed up-regulation of Bax protein, while compound 7 (non-toxic compound) showed down-regulation of the same protein in apoptotic array analysis (Figure 2). These findings indicate that compounds 5 and 6 show cytotoxicity by inducing apoptosis. These results are in agreement with previous reports indicating that coumarin inhibits cancer cell growth and induces apoptosis in human non-small cell lung carcinoma (NSCLC) cell lines (23-26).
Many anticancer agents are said to induce apoptosis by targeting the proteins of the Bcl-2 family (Bax/Bcl-2), which play a critical role in the determination of whether cells will undergo apoptosis (27-29). The up-regulation of Bax protein (pro-apoptotic protein) sensitizes cells to chemotherapeutic agents (30), while the down-regulation of Bcl-2 (anti-apoptotic protein) may delay the response of tumor cells to applied therapies (31, 32). Bax protein, associated with the formation of homodimers and production of channels, resulted to loss of mitochondrial membrane potential and allowed the release of cytochrome c (33, 34). In contrast, Bcl-2 protein expression is known to inhibit cytochrome c released from the mitochondria (34, 35). In the current study, western blot results indicated that compound 5 showed cytotoxicity by the induction of apoptosis; increased in pro-apoptotic Bax expression and decreased in anti-apoptotic Bcl-2 expression. This molecular mechanism is similar to those of other reported coumarin derivatives (33-37). However, compound 6 showed down-regulation of both Bax and Bcl-2 protein expressions by western blot analysis. These results indicate that compound 5 shows cytotoxicity via activation of an apoptosis process, while compound 6 through some other mechanism.
Conclusion
Our investigation has shown the molecular events associated with these coumarin-type derivatives in a human lung (A549) cancer cell line. The apoptosis array analysis showed differential expression of apoptotic proteins in treated cell extracts, which may be responsible for the toxicity and less toxicity, or non-toxicity of these compounds in A549 the cell line. The differences in Bax/Bcl-2 protein expression further suggest that these compounds inhibited cell growth in human lung cancer (A549) cell line by inducing apoptosis. Further investigation would involve the identification of the most important signaling pathways in regulating apoptosis. On the basis of these results, we hope that our findings could contribute to the understanding of how these coumarin-type compounds suppressed the growth of the lung cancer cells.
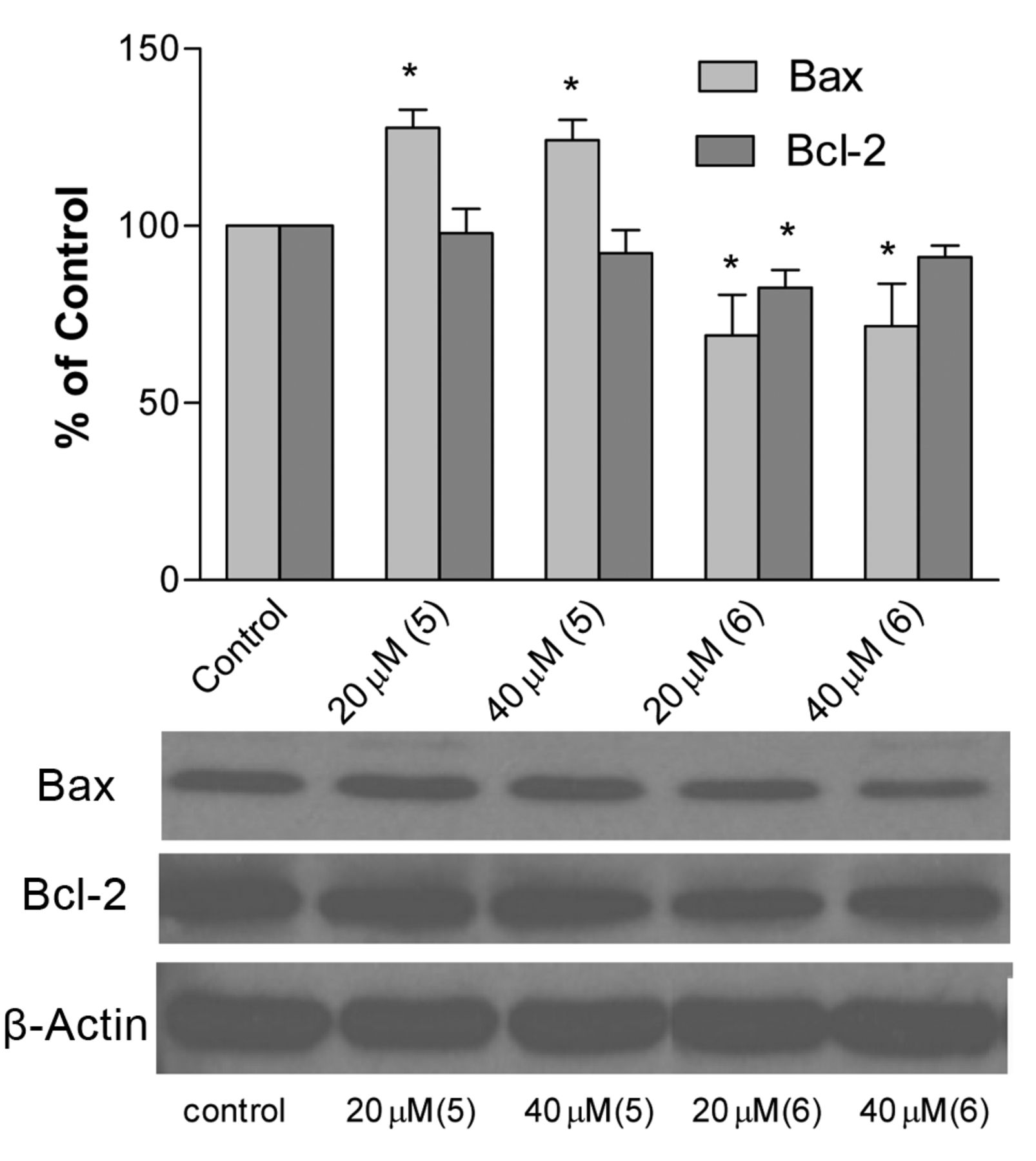
The effect of compounds 5 and 6 on Bax and Bcl-2 protein expression in human lung (A549) cancer cells, after incubation for 6 h, as analyzed by Western blotting.
Acknowledgements
The Authors gratefully acknowledge Florida A & M University TITLE III PROGRAM for their financial support.
- Received August 6, 2012.
- Accepted September 3, 2012.
- Copyright© 2012 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved





{kind=link}
{kind=link}
{kind=link}