


Abstract
Background: Antineoplastic phospholipids (ALPs) represent a promising class of drugs with a novel mode of action undergoes rapid turnover in the cell membrane of tumors, interfering with lipid signal transduction, inducing cell death. The aim of this study was to investigate the synthetic phosphoethanolamine (Pho-s) as a new anticancer agent. Materials and Methods: Cell viability and morphology were assessed by (3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT), Hoechst and rhodamine staining. Apoptosis was assessed by Annexin V and propidium iodide (PI) staining, caspase-3 activity, mitochondrial membrane potential (ΔmΨ) and cell cycle analysis, combined with evaluation of tumor growth in Ehrlich Ascites Tumor (EAT) bearing mice. Results: We found that Pho-s 2.30 mg/ml induced cytotoxicity in all tumor cell lines studied without affecting normal cells. In vitro studies with EAT cells indicated that Pho-s induced apoptosis, demonstrated by an increase in Annexin-V positive cells, loss of mitochondrial potential (ΔmΨ) and increased caspase-3 activity. It was also shown to increase the sub-G1 apoptotic fraction and inhibit progression to the S phase of the cell cycle. Additionally, antitumor effects on the EAT-bearing mice showed that Pho-s, at a concentration of 35 and 70 mg/kg, inhibited tumor growth and increased the lifespan of animals without causing liver toxicity. Conclusion: These findings suggest that Pho-s is a potential anticancer candidate drug.
Development of new drugs has played an important role in the treatment of cancer and a number of new agents and combinations have entered clinical trials, with promising results. The antineoplastic phospholipids (APs) analogs of lysophosphatidylcholine collectively named alkylphospholipids (ALPs) exert cytotoxic effects against a wide variety of tumors (1). These agents do not target the DNA, but act at the level of cell membranes. Although the antitumor mechanism remains unclear, the biological effects of APs are encouraging for treatment of malignant tumors, because of the high degree of cytotoxic effects against a wide variety of cancer cell types resistance to chemotherapy drugs (2-4).
APs represent a group of compounds that can be used in combination with other chemotherapy agents or radiotherapy. There are actually some APs that have been tested in clinical trials, such as (ET-18-OCH3; edelfosine), which exhibits cytotoxic activity against human and murine tumor cells in vitro and in vivo, as well as miltefosine (used clinically for the treatment of mammary carcinomas). These antineoplastic effect reducing the expression of integrins and adhesion molecules, which them reduce tumor cell attachment and metastasis (5-6). Although there is no consensus for the antineoplastic mechanisms of these lipids, antitumor action may be attributed to modulation of the biosynthesis of lipids, i.e. rapid turnover of phospholipids, leading to their accumulation in cell membranes, thereby inhibiting mitogenic signal transduction and inducing cell death (7).
Many anticancer drugs, such as edelfosine and perifosine, have been considered promising drugs in cancer treatment, inducing mitochondrial damage by increasing the mitochondrial transition pore, resulting in translocation of proapoptotic protein BAX and cytochrome c from mitochondria to the cytosol, resulting in the activation of caspase cascade-mediated apoptosis (8-11).
Therefore, the first-generation antineoplastic lipids, the ALPs, in particular edelfosine, were structurally modified to overcome their metabolic instability allowing them to be used for treatment of malignant tumors. Later studies with ALPs have shown that a long alkyl chain and a phosphocholine moiety may be sufficient for antineoplastic effects (12).
In a recent study, we also showed that treatment with Pho-s in an animal model of B16 F10 tumor resulted in tumor growth inhibition and reduction of metastasis. We also found that Pho-s triggered apoptosis by reducing expression of proapoptotic BAD/BAX, leading increased caspase-3 activity, and blocked S and G2/M phases of the cell cycle (13).
Based on these findings, we evaluated the biological activity of Pho-s in different tumor cell lines, as well the effect on a model of Ehrlich ascitic tumor (EAT).
Materials and Methods
Compound and antibody. Synthetic phosphoethanolamine was synthesized supplied and in the Laboratory of Chemistry and Polymers of Technology, University of Sao Paulo, Sao Carlos, Brazil. An antibody against Caspase-3 was obtained from Biotium Inc. (Hayward, CA, USA) and Hoechst 33342/PI, rhodamine 123, propidium iodide and RNase were purchased from Sigma (St. Louis, MO, USA). Annexin V- fluorescein isothiocyanate (FITC) apoptosis detection kit was obtained from BD Biosciences (Franklin Lakes, NJ, USA).
Tumor model and EAT cell culture. EAT cells was obtained from Institute of Biomedical Sciences, University of Sao Paulo (Sao Paulo, SP. Brazil) and maintained in male BALB/c mice, 2-4 weeks in ascites form by successive transplantations. For in vitro assays, MCF-7 (human breast adenocarcinoma), Skmel-28 (human malignant melanoma), Mewo (human malignant melanoma), B16F10 (murine malignant melanoma), H292 (human mucoepidermoid pulmonary carcinoma) and Huvecs – CRL 1730 (human umbilical vein endothelial) cells were obtained from American Type Culture Collection (Mannasa, VA, USA). FN1 (human normal fibroblast) and lymphocyte (murine lymph node) cells were obtained from Biochemistry and Biophysical (Butantan Institute, SP, Brazil). All cells were cultured in RPMI 1640 supplemented with 10% fetal bovine serum (FBS), 100 U/ml penicillin, 100 μg/ml streptomycin and 100 μg/ml L-glutamine in culture flasks at 37°C in a humid atmosphere containing 5% CO2. For in vivo assays BALB/c mice with 2-4 weeks were inoculated with 106 EAT cells (0.2 ml/mouse) intraperitoneally (i.p.). After 24 h from tumor implantation, the animals were randomized and divided into different groups (n=5). The treatment of Pho-s was started at doses of 35 and 70 mg/kg/day continued for 15 days.
MTT assay. Cells EAT, MCF-7, Skmel-28, Mewo, B16 F10, H292, FN1, lymphocytes and Huvecs were plated in triplicate in 96-well plates at a density of 5×104 cells/well into flat microtiter plates and incubated for 1 day at 37°C in a humidified incubator containing 5% CO2 and the next day the cells were treated with Pho-s 10 to 0.5 mg/ml in a final volume of 100 μl. After 24 h of treatment, cells were exposed to 5 mg/ml MTT for 2 h, and the precipitated formazan was dissolved in 0.1N Hcl in isopropanol and measured at 540 nm with a microplate reader Thermo Plate (Rayto Life and Analytical Sciences C. Ltd, Germany) and the IC50 values determined.
Hoechst 33342/PI assay by fluorescence. EAT cells (106/ml) were plated on glass and incubated overnight at 37°C with 5% CO2. The culture medium was exchanged and after 1 h 2.30 mg/ml Pho-s was added to independent wells, and the cells furthers incubated for 12 h. The next day, 1 ml of culture medium was added containing 10 mg/ml Hoechst 33342 (H), a membrane-permeable dye that specifically binds to chromatin and appears green when inspected under UV light and 1 mg/ml PI, (Sigma) added at 20 min. 37°C, 5% de CO2. Cells positive stained with PI show an increase in membrane permeability (necrosis), while cells positively stained with PI and H are pink (end-stage apoptosis) and those positively stained with H alone present cells with intact nuclei. Analysis was performed by fluorescence microscopy using a Nikon Eclipse E1000 instrument (Nikon, Kanagawa, Japan). Digital images were obtained with a CCD camera (Applied Imaging ER model No. 339, Santa Clara, CA, USA) and the documentation system used was Cytovision V.2.8 Tax (Applied Imaging Corp).
Measurement of mitochondrial membrane potential. The mitochondrial membrane potential (ΔmΨ) was measured by rhodamine (Rho123) assay, monitored by flow cytometry. EAT cells (106 cells/well) were seeded in 6-well plates and incubated for 24 h then treated for 12 h with Pho-s at 2.30 mg/ml. In the present study, Rho123 (100 mg/l) was added to cell cultures maintained at 37°C and after 45 min fluorescence was measured with flow cytometry with FACScalibur (Becton Dickinson). A total of 10000 cells/sample were analyzed and the mean fluorescence intensity recorded.
Apoptosis assay by Annexin-V/PI double-staining. The Annexin V-FITC/PI apoptosis detection kit (BD Bioscience) was used to detect the effects of Pho-s. EAT cells (106 cells/well) were seeded in 6-well plates and incubated for 24 h. The cells were treated with 2.30 mg/ml Pho-s for 12 h and adherent and floating cells were collected and washed in PBS. The cells were centrifuged and the cell pellet was suspended with binding buffer (100 μl). The cells were then stained with Annexin V-FITC (2 μl) and PI (2 μl), incubated for 15 min, at room temperature in the dark. After incubation, 400 μl binding buffer was added and cells were analyzed with FACScalibur (Becton Dickinson) using CellQuest software, determining the percentage of apoptotic cells. A minimum of 10000 events was acquired for each sample.
Determination of caspase-3 activity by cytometry flow. Caspase-3 activity in the EAT cells was determined by flow cytometry using NucView assay. NucView™ 488 Caspase-3 substrate (Biotium Inc., Hayward, CA, USA) is a cell membrane-permeable caspase substrate designed for detecting caspase-3 activity. The substrate is cleaved by caspase-3 in the cytoplasm of apoptotic cells, releasing a fluorescent DNA dye that is able to enter the nuclei and stain the DNA. EAT cells (106 cells/well) were seeded in 6-well plates and incubated for 24 h and treated for 12h with Pho-s 2.30 mg/ml. Briefly, 200 μl of cell suspension were transferred to a flow cytometry tube and 2 μl of NucView™ 488 caspase-3 substrate were added to the cells, followed by incubation at room temperature in the dark for 15 min, then analyzed using flow cytometry FACScalibur (Becton Dickinson). The dot plots of events were recorded in the FL2 channel and 10000 events were acquired for each sample in three independent experiments.
Cell cycle phase analysis by flow cytometry. EAT (106) cells were treated for 12 h at the 2.30 mg/ml IC50 concentration value of Pho-s obtained by MTT assay. Cells were then collected and fixed with cold 70% ethanol/20μg/ml RNase (Sigma) and stored at −20°C, washed and resuspended in PBS. Cells were incubated at 37°C for 45 min in 0.5 ml PBS and then stained with PI (Sigma) for 30 min at 37°C. Quantification of DNA content 10000 events was acquired for each sample by flow cytometry.
Confocal laser scanning microscopy. A total of 106 EAT cells were incubated in the dark at 37°C with Pho-s 2.30 mg/ml for 12 h, then rinsed in PBS three times and stained with 10 μM Rho123 (Sigma) for 30 min at 37°C; excess probe was then washed off and live cells were submerged in PBS. Image analysis was accomplished with a confocal laser scanning microscope (Carl Zeiss LSM 700; Leica, Mannheim, Germany). Post-acquisition image processing, background correction, adjustment of brightness and contrast and export to tiff format, were done with Image J software (version 14.1) National Institutes of Health (Bethesda, Maryland, USA).
Evaluation of effects antitumor. The antitumor activity of Pho-s was evaluated by tumor growth inhibition. To estimate effect of Pho-s on tumor growth dynamics, the animals body weights were monitored every day from tumor cell inoculation and at the end of the treatment with Pho-s at 15 day. For the survival analysis, the animals were weighed daily until death, then the effect of Pho-s on lifespan was calculated on the basis of the mortality of the experimental mice. The animal model experiments were carried out in accordance with the guidelines for animal experimentation determined by the Institutional Animal Care from Butantan Institute (process number 566/09).
Biochemical analysis. Blood samples were collected from the maxillary vein after 15 days of treatment from each animal collected and were separated into one vial. These samples were centrifuged at 2000 rpm for 10 min and placed at −20°C to separate the serum for the biochemical determinations, including aspartate aminotransferase (AST) and alanine aminotransferase (ALT) activities, and lactate dehydrogenase (LDH) levels. The biochemical determinations were carried out using the fully automated COBAS System Analyze (Roche, Burgess Hill, West Sussex,UK).
Statistical analysis. Data are expressed the as mean±standart deviation (S.D.) of three independent experiments performed in triplicate. One-way analysis of variance (ANOVA) was carried out determine statistically significant differences from untreated controls.
Results
Evaluation of the cytotoxic effects of Pho-s in tumor cell lines by MTT assay. The results show that the Pho-s had no effect on cell viability of normal human endothelial cells, fibroblasts and murine lymphocytes. However, after 24 h of treatment, Pho-s exhibited potent cytotoxic activity against all tumor cells used in this study, with the greatest effect towards, human melanoma Skmel-28 cells (IC50=1.20 mg/ml). These results indicated in response to Phos-s there is no different between tumor cell lines as shown by the mean IC50 values (Table I).
Apoptotic effect of Pho-s on morphological changes in EAT cells. EAT cells were treated with Pho-s at 2.30 mg/ml for 12 h, with changes on mortality and viability being assessed by Hoechst 33342/PI staining. Quantitative analysis Pho-s at 2.30 mg/ml were cytotoxic to EAT cells (***p<0.001) (Figure 1A). Compared with untreated cells, in the presence of Pho-s, EAT cells retracted and the cytoplasm became condensed. The extent of the apoptotic effect after treatment was evidenced by typical chromatin margination (H+/PI+ or H+/PI−) in the cell nuclei of virtually all cells, and as reduction in cell volume (Figure 1B).
Cytotoxicity of Rho-s towards tumor cell lines and normal cells.
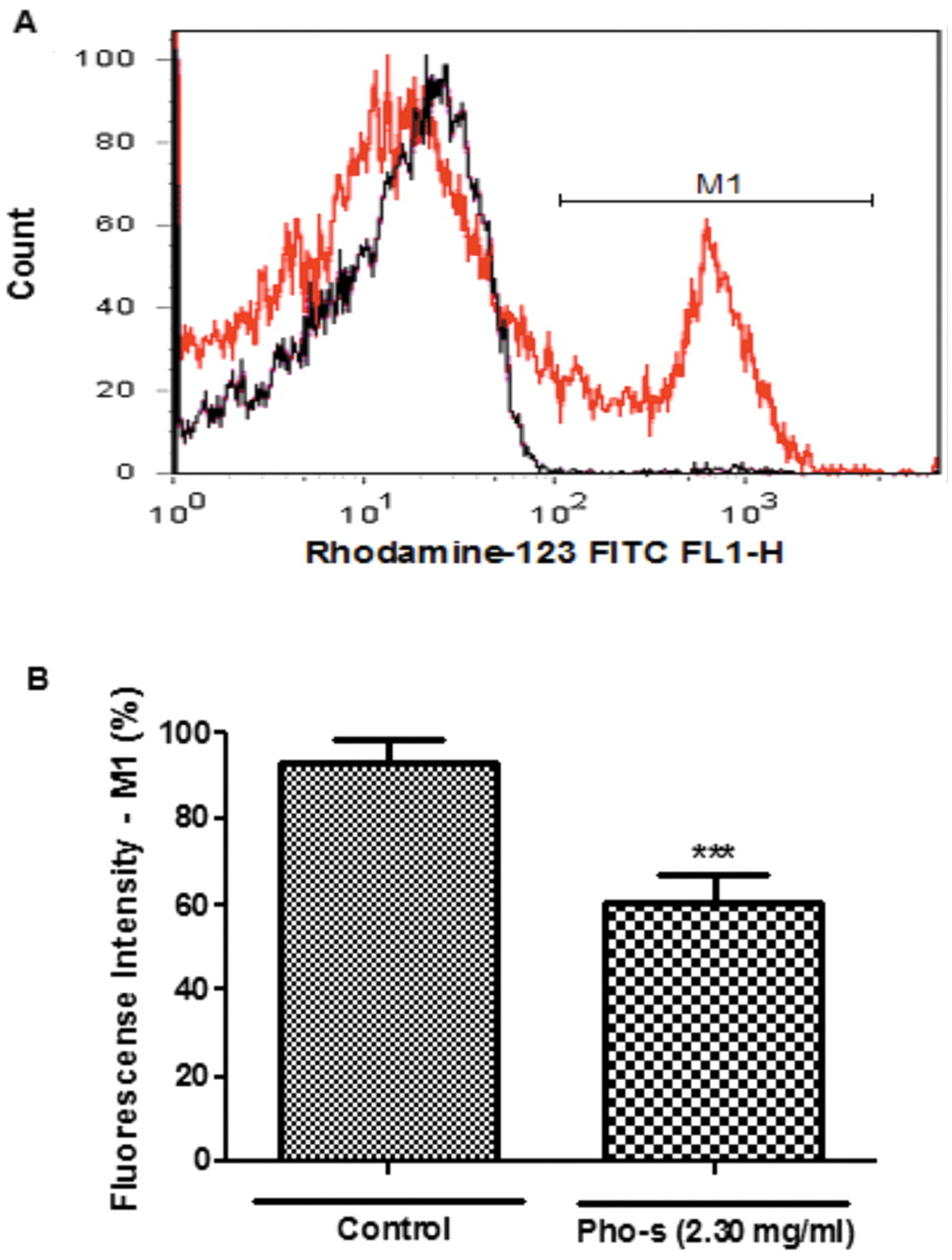
Pho-s induced changes in ΔmΨ of EAT cells. Measurement of ΔmΨ was conducted to verify whether dysfunctional mitochondria were linked to apoptosis. We found that Pho-s reduced the ΔmΨ (56±2.3%) measured by fluorescence intensity in treated cells compared with control cells (89±6.2%) (***p<0.001). The histograms are presented in overlapping spectra of the control (red line) and the cells treated with Pho-s (black line). The mean values and standard deviations are shown in Figure 2A and B.
Mitochondrial morphological change on treatment of EAT cells with Pho-s. Changes in mitochondrial morphology were studied by using the fluorochrome Rho123. EAT control cells showed no morphological alteration, as indicated by the brilliant green cytoplasmic fluorescence uniformly distributed in the tumor cells. Pho-s at 2.30 mg/ml induced progressive reduction of the mitochondrial fluorescence and morphological changes. After 12 h, Phos-s had induced change in mitochondrial shape from round to an elongated rod-like shape, as shown in the three-dimensional analysis performed by Image J software (version 14.1) National Institutes of Health (Bethesda, Maryland, USA) (Figure 3). Pho-s induced apoptosis and increased caspase-3 activity in EAT cells. Double staining of EAT cells with Annexin V/PI showed that Pho-s induces morphological cell change (Figure 4A and B) and cell death by apoptosis, with externalization of phosphatidylserine. Treatment with Pho-s increased the number of cells in early apoptosis (41±2.1%) and late apoptosis (33±4.5%), compared with controls (***p<0.001) (Figure 4C and E). Treatment with Pho-s significantly increased caspase-3 activity (67±5.3%) in relation to the control (17±3.2%) (***p<0.001).The data demonstrated that the number of apoptotic cells was found to be increased (***p<0.001) in dose-dependent manner (not shown) and the caspase-3 activation was correlated with apoptosis induced by Pho-s (Figure 4F).

Morphological analysis of (EAT) cells at 6 h after Pho-s treatment by staining with Hoechst 33342 (H) and propidium iodide (PI). Control cells showed no change in membrane structure and consisted mostly of living cells with blue-stained nuclei (A). EAT cells treated with Pho-s 2.30 mg/ml IC50 demonstrated reduced cell viability (**p <0.001) and membrane integrity displayed extensive double staining with H+&PI+ in pink (arrows and circle), accompanied by an increase of apoptotic cells (scale bar, 30 μm) (B).
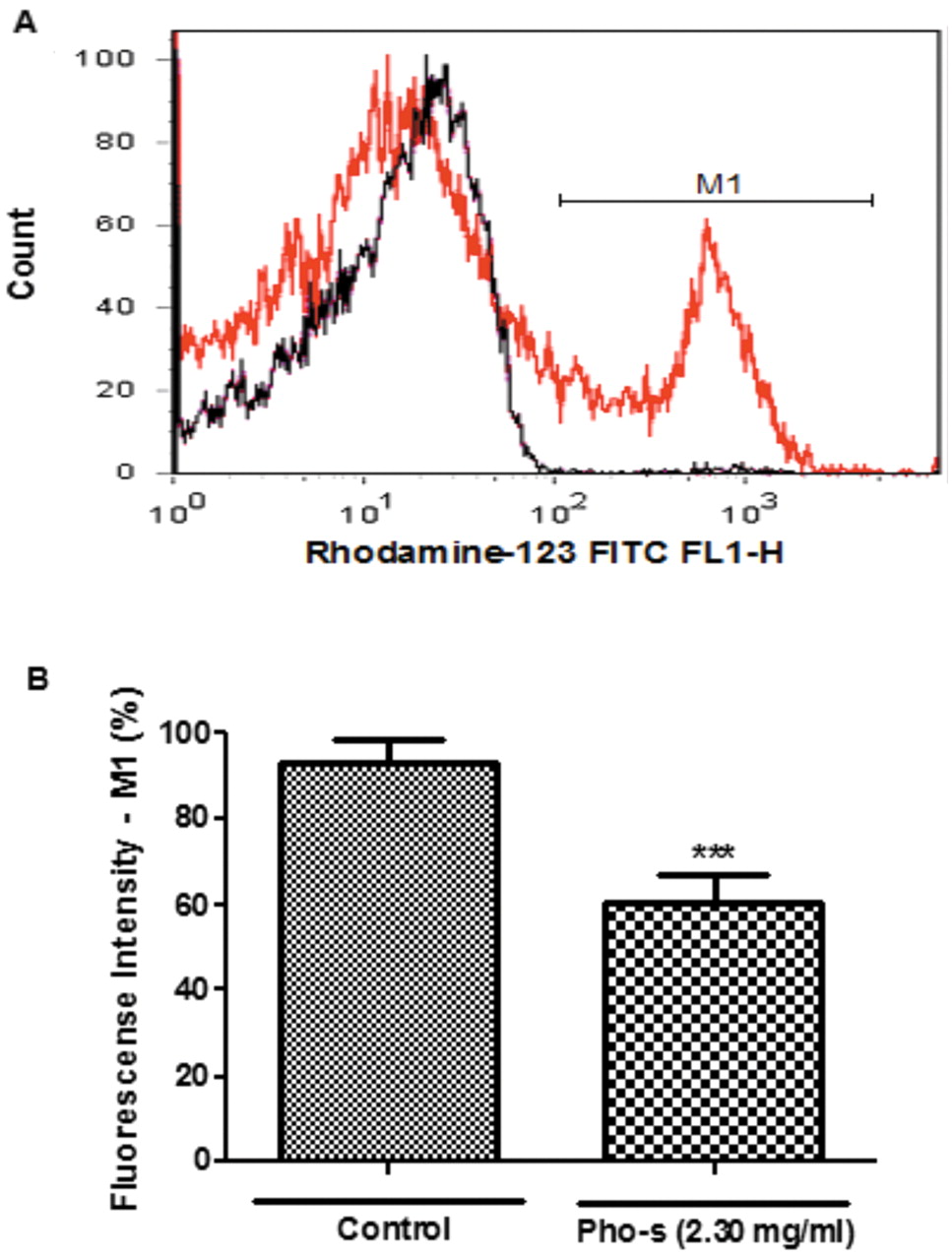
Effect of Pho-s on mitochondrial membrane potential (ΔmΨ). EAT cells were incubated with Pho-s at 2.30 mg/ml for 6 h and ΔmΨ measured by Rho 123 fluorescence analyzed by flow cytometry (A). There is a substantial shift in fluorescence emission, indicating a decrease of the ΔmΨ of the Pho-s treated (line red) compared to control (line black) cells. The difference (**p<0.001) was considered to be significant as shown in bar graphs (B). The data represent averages of three independent experiments.
Pho-s reduces the proportion of cells in the G2/M phase and increases the number of apoptotic sub-G1. Cells tumor growth inhibition and induction of apoptosis by Pho-s correlated with ability to induce change in the cell cycle. As regards the G0/G1 and S phases of the cycle, there was significant (***p<0.001) change in the median percentage of cells in each phase after treatment with Pho-s at 2.30 mg/ml in comparison with the population of control cells G0/G1 (Figure 5A). As shown in Figure 5B, after 12 h of treatment with Pho-s, the proportion of cells in the S and G2/M phases decreased. Results showed that Pho-s number increased the number of tumor cells in the sub-G1 peak with DNA fragmentation, which was accompanied by a decrease in S and G2/M phases, inducing apoptosis; these results were confirmed with Annexin-V assay. These date suggest that Pho-s mediates antiproliferative and proapoptotic activity on EAT cells associated with a decrease of cells in the S phase, as well as DNA fragmentation (apoptosis).
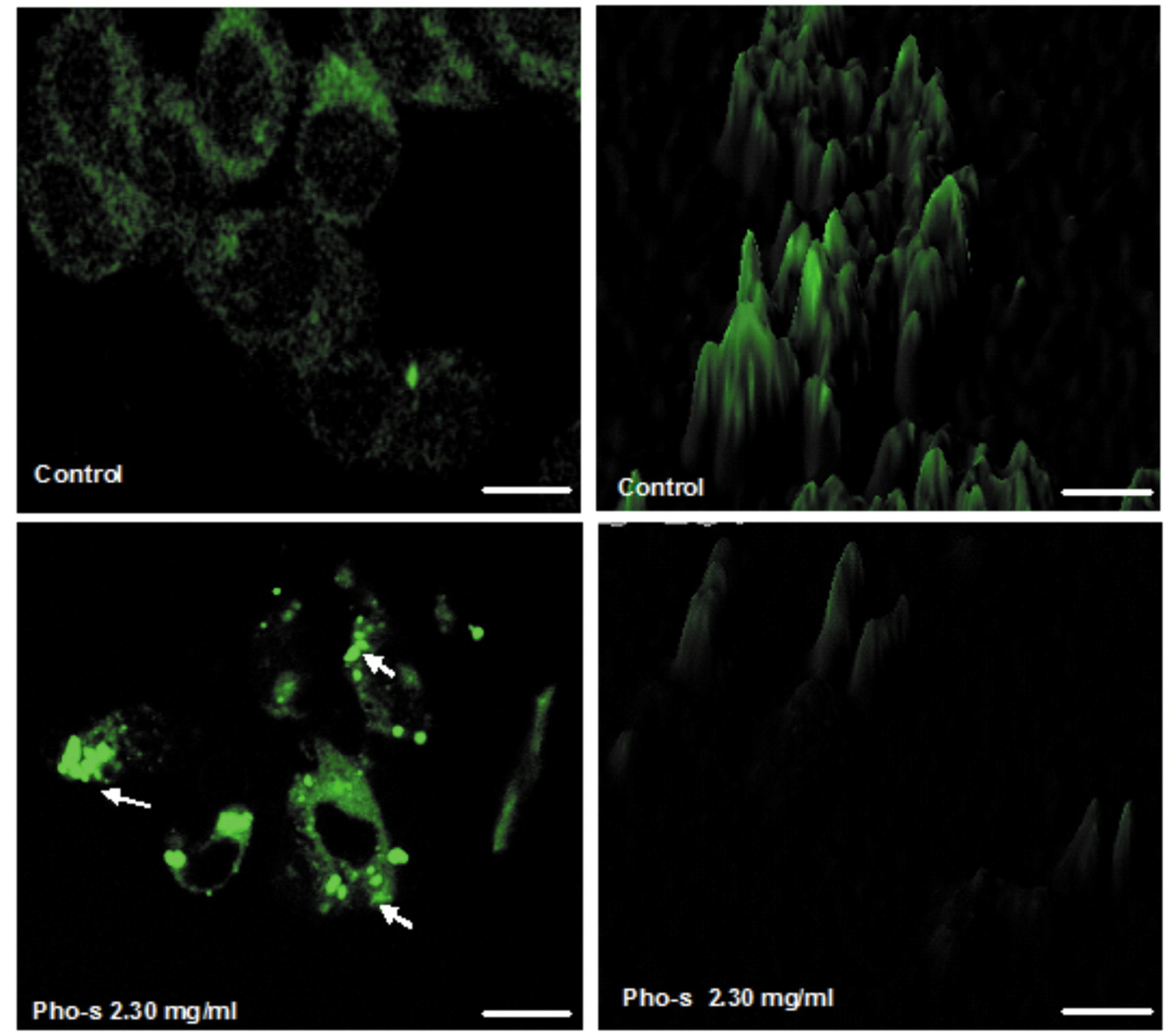
Representative confocal fluorescent micrographs of mitochondrial morphology staining of EAT cells with Rho 123.Control cells showing the normal heterozygous intracellular mitochondria of living cells with high green fluorescence signal. Notice EAT cells treated with Pho at 2.30 mg/ml exhibited lower intensity fluorescence and marginalization of mitochondria (arrows),a vital and dynamic process that occurs in apoptosis (scale bar, 30 μm).
Pho-s inhibits tumor growth and increase survival of EAT-bearing mice. No significant alterations of the body weight of treated mice were observed during the sequential treatment with Pho-s at concentrations of 35 and 70 mg/kg (***p<0.001) compared with untreated mice (Figure 6A). Pho-s resulted in the inhibition of neoplasic ascites and of tumor growth (Figure 6B). In addition, to elucidate whether the tumor inhibition seen with Pho-s treatment was complete or partial, we stopped treatment on day 15. The animals body weigh did not change for a further 15 days, suggesting that cancer cells had been completely eradicated (Figure 6C). In accordance with these results, log-rank tests indicated a significant increase in survival probability in the overall survival of mice treated with Pho-s at 35 mg/kg (p=0.0457) and 70 mg/kg (p=0.0219).

Apoptotic effect on EAT cells treated with Pho-s at 2.30 mg/ml was assessed using Annexin -V and PI double staining of caspase-3 activity by flow cytometry. For all populations cells are gated into two populations according to FSC and SSC and it was found that compared with control (A) Pho-s changed cell morphology reducing density of tumor cells in R1 and R2 marked region (B). Pho-s treatment compared with control (C) significantly increased the number of cells are in an early stage of apoptosis, and a population of cells are at a later stage of apoptosis (D) as shown in the bar graphs (E). Measurement of caspase-3 activity was determined after 6 h of treatment with Pho-s showing increased caspase-3 activity correlates with the frequency of apoptotic cells shown in the Annexin-V assay (F). The bar graphs are the averages ± S.D. of multiple determinations.

Pho-s induced G2/M decrease and apoptosis of EAT cells. Apoptotic fraction was recognized as the sub-G1 population compared with control (A), showing a significant increase of this in the cells treated with Pho-s at 2.30 mg/ml. In addition, a reduction of S phase and a decrease in G2/M phase indicate that Pho-s has antiproliferative activity (B). Data are the mean ± S.D. of three different experiments performed in duplicate.

Pho-s ameliorated weight loss and increased the survival rate of EAT bearing mice. EAT cells (1×105 per mouse) were injected i.p. in BALB/c mice. Forty-eight after tumor implantation, the mice were treated daily via i.p. injection with Pho-s (35 and 70 mg/kg). Pho-s treatment at both concentrations significantly reduced ascites tumor measured by changes in body weight at the end of the experiment compared to untreated control (A) as showing the macroscopic aspects of ascitic tumor at the end of treatment (B). In addition, suppression of tumor growth, as shown by a Kaplan-Meier curve, by Pho-s positively overall survival rate (C). The survival curves were significantly different with Pho-s at 35 mg/kg (p=0.0457) and 70 mg/kg (p=0.0219).
Pho-s prevent increases in the serum level of biochemical markers. Biochemical parameter analysis from the untreated mice revealed high serum level of AST (245±45.7U/l), ALT (123±10.8U/l) and LDH (8266±1114U/l), in agreement with increased tumor growth. Animals treated with Pho-s at a concentration of 70 mg/kg showed no significant alterations of AST (80±4.5U/l), ALT (20±1.2U/l) Figure 7A, LDH (1088±73 U/l), as well as, on the concentration of 35 mg/kg (68±4.3U/l), ALT (19±6.7U/l) and LDH (1317±101U/l) compared to untreated mice (Figure 7A-7B). It is reasonable to suggest that inhibition of neoplastic ascites by Pho-s prevents failure liver.
Discussion
Current studies aimed at developing effective drugs targeting with focused mainly on the utilization of composts that can induce selective toxicity in cancer cells. Active research in has led to extensive investigations into novel phospholipids that have been demonstrated to play a major role in the regression of cancer with low general toxicity (14-18). Phospholipids as miltefosine, ALPs and edelfosine are known to exhibit significant antitumor activity and have been used very successfully in anticancer therapies against skin metastases of breast cancer and in the topical treatment of cutaneous malignant lymphomas. However, the precise mechanism of the cytotoxic effect of ALPs remains unclear (19-21). Thus, in this study we used EAT cells in vitro to study the morphological changes and the mechanisms of cell death induced by Pho-s treatment. Firstly, in vitro anticancer cytotoxic activity of Pho-s was measured by MTT. Interestingly, the level of cytotoxic sensitivity among tumor cells analyzed reveals that the Pho-s induces cytotoxicity in all cell tumor lines used in this work, independently of molecular features, such as apoptosis and chemotherapy resistance.
The first act of many anticancer agents is inducing apoptosis, which can follow the intrinsic or extrinsic pathway. Mitochondrial outer membrane permeabilization (MOMP) has a central role in the signal transduction cascade of cell death to release apoptotic proteins that are associated with early events to trigger apoptosis. Our investigation confirms that Pho-s reduces ΔmΨ of EAT cells by depolarization, with increased MOMP, which demonstrates that Pho-s most likely induces apoptosis by an intrinsic pathway (22). Once the IC50 value was obtained, the effects of Pho-s in the cell morphology was investigated. Hoechst/PI stains have provided significant evidence that Pho-s induces apoptosis. The cells treated with Pho-s showed morphological changes, such as, nuclear retraction followed by chromatin condensation and shrinkage cell, which are apoptosis features.
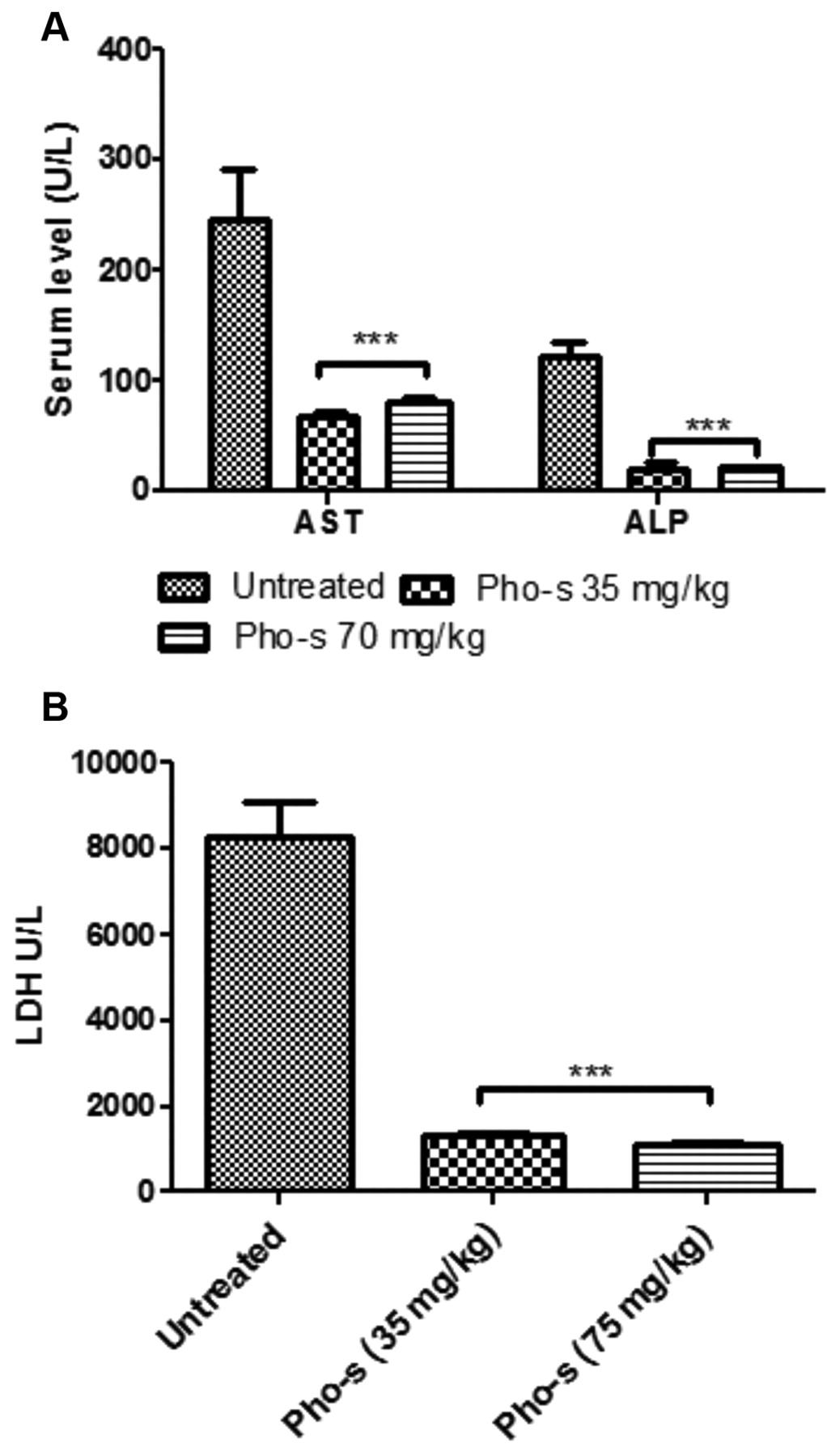
Biochemical evaluation. Serum levels of hepatic transaminases (A), aspartate aminotransferase (AST), alanine aminotransferase (ALT), and (B) lactate dehydrogenase (LDH) reveals that Pho-s (35 and 70 mg/kg) prevents elevation of serum levels induced by EAT cells compared to the untreated control. This protective effect likely correlates with the antitumor effects of Pho-s.
Since the mitochondrial studies indicated that Pho-s induces change of ΔmΨ of EAT cells, a second investigated strategy was to identify changes in mitochondrial morphology by confocal laser scanning microscopy. It was found that the reduction of ΔmΨ was linked to structural changes in the mitochondria, with diffuse distribution of Rho123 at the cytoplasm and a decrease in the intensity of fluorescent staining, as well as the formation of peripherals clusters and marginalization of mitochondria.
An increase in MOMP leads to the release of apoptogenic proteins from the mitochondria, mainly cytochrome c, that complex with the cytoplasmic protein (Apaf-1), which then oligomerizes and binds to procaspase-9. The complex apoptosome results in the activation of caspase-9 that is able to cleave and activate the effector caspases -3,- 6 and 7. Analysis of caspase activation on EAT cells strongly supports that increase of caspase-3 on EAT cells treated with Pho-s might be due to ΔmΨ shift (23-24). Once activated, the effector caspases, such as caspase-3 can lead to cell death and might target specific substrates, such as actin and nuclear lamin A, leading to DNA fragmentation, chromatin condensation, and the formation of apoptotic bodies (25-27). These apoptotic features of EAT cells treated with Pho-s suggests the involvement of an intrinsic apoptotic pathway.
In the present study, we demonstrated that in EAT cells, Pho-s mediated cell cycle reduced of the G2/M follow of S phase decreased. In addition, these results support that despite of Pho-s induce apoptosis increased of the sub-G1 peak apoptotic, can also act as a compound with antiproliferative activity inhibits cell cycle progression on EAT cells. However, the exact mechanism by which Pho-s inhibits cell cycle progression remains unclear. Thus, further studies are needed to indentify how Pho-s regulates of signal transduction in the cell cycle and inhibits cell tumor proliferation. This mechanism is the target of many citotoxic drugs that induce cell cycle arrest (28-29).
In the in vivo study, we have also shown that treatment of mice bearing EAT cells resulted in inhibition of growth of tumor cells in the peritoneal cavity as measured by body weight. This antiproliferative activity of Pho-s prevented a gradual increase of body weight see in untreated tumor-bearing mice, and can be partially explained from the in vitro results that showed Pho-s blocking, of the S phase in the cell cycle, with significant increase of apoptosis. The most important finding of our in vivo study was that Pho-s was able to increase the median survival of EAT-bearing mice and controlled completely ascites growth without apparent toxicity.
The study of liver functions by biochemical determination of ALT, AST and LDH revealed no abnormal serum levels compared to untreated animals and indicates that Pho-s treatment induced no toxicity during the experimental period. Therefore, the increase of biochemistry markers of EAT-bearing untreated mice was probably due to the strong pressure exited into peritoneal cavity which increased pressure of the blood. Thus, this pressure causes accumulation of extracellular fluid that forces fluid out of the blood vessels which them collects in the abdominal cavity, causing liver failure (30).
In conclusion, Pho-s exhibited cytotoxic effects in all cell tumor lines and non-normal cells. Based on results from experiments in vitro with EAT cells, we can assume that the Pho-s induces apoptosis most likely by an intrinsic pathway, due to many features associated with apoptosis, including apoptotic morphology, externalization of phosphatidylserine, reduced ΔmΨ and activation of caspase-3. In addition, Pho-s induces a decrease of G2/M and S phases of the cell cycle in EAT cells, but these antiproliferative effects still need further studies. In agreement with in vitro data, Pho-s exerted antitumor effects in the EAT-bearing mice model inhibiting tumor growth and substantially increasing the lifespan of animals, without causing liver toxicity.
Acknowledgements
This work was supported by a Sao Paulo Research Foundation – (FAPESP) grant (2007/50571-3; Doctorate and Master Fellowship 2008/56089-1; 2010/50220-9).
Footnotes
-
Conflict of Interest Statement
The Authors declare that there are no conflicts of interest.
- Received August 29, 2011.
- Revision received November 25, 2011.
- Accepted November 29, 2011.
- Copyright© 2012 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Preclinical Development of a Nontoxic Oral Formulation of Monoethanolamine, a Lipid Precursor, for Prostate Cancer Treatment
- More Convoluted Than a Brazilian Soap Opera: How an Eager Chemistry Professor and a Well-Intended but Misguided Federal Judge Ignited an Industry of False Hopes
- Meclizine Inhibits Mitochondrial Respiration through Direct Targeting of Cytosolic Phosphoethanolamine Metabolism