


Abstract
We report the synthesis and in vitro activity of trans-bis-(2-hydroxypyridine)dichloroplatinum(II) (coded as DH3) in humour ovarian tumour models. The compound is less active than cisplatin against the parent cell line A2780 but more so against the cisplatin-resistant A2780cisR cell line, thus indicating that it is better able to overcome mechanisms of resistance operating in the A2780cisR cell line. DH3 is marginally less active than cisplatin against ZD0473-resistant A2780ZD0473R cell line but with a much lower resistance factor than cisplatin. DH3 has higher platinum–DNA binding levels than cisplatin in the A2780 and A2780ZD0473 cell lines and a lower value in the A2780cisR cell line. Even though DH3 has a lower activity than cisplatin, the higher platinum–DNA binding levels observed for DH3 than cisplatin in A2780 and A2780ZD0473R cell lines may not be entirely unexpected when we note that the two compounds are likely to differ in their nature of binding with DNA. Whereas cisplatin binds with DNA forming mainly intrastrand 1,2-Pt(GG) and 1,2-Pt(AG) adducts, DH3 is expected to form more 1,2-interstrand Pt(GG) and monofunctional adducts. The higher activity of DH3 than cisplatin in the A2780cisR cell line despite its lower level of platinum–DNA binding can also be seen to indicate the complexity of the situation. Although platinum–DNA binding may be an essential requirement for apoptosis, it is not sufficient to cause cell death that is actually brought about by downstream processes in the cycle. The results of interaction with pBR322 plasmid DNA combined with BamH1 digestion show that DH3 is less able to prevent BamH1 digestion than is cisplatin, indicating that cisplatin causes a greater conformational change in the DNA than DH3. Conclusion: DH3 is less active than cisplatin against the parent cell line A2780, but more so against the cisplatin-resistant A2780cisR cell line, thus indicating that it is better able to overcome mechanisms of resistance operating in the A2780cisR cell line.
Although widely used in the clinic, cisplatin has a limited spectrum of activity due to intrinsic and acquired resistance (1, 2) and a number of dose-limiting side effects (3-6). Currently attention is also being given to platinum compounds that would bind with DNA differently from cisplatin, with the idea that the difference in nature of binding with DNA may translate into an altered spectrum of activity. One such class of compounds is trans-planaramine-platinum(II) complexes that are expected to bind with DNA forming mainly monofunctional Pt(G) and bifunctional interstrand Pt(GG) adducts as against 1,2-intrastrand Pt(GG) and Pt(AG) adducts formed by cisplatin (7). A number of trans-planaramine-platinum complexes have been prepared by Huq and co-workers (8-10) and others (11-13) that have shown in vitro activity similar to that of cisplatin against the various cancer cell lines. One of the key findings is that the activity of trans-planaramine-platinum(II) complexes can vary widely depending on the nature of the actual planaramine ligand, thus illustrating the structure–activity relationships. For example, it was found that 2-hydroxypyridine and 3-hydroxypyridine are more activating than is 2,3-diaminopyridine. We report here the synthesis, characterization, cytotoxic activity, cellular accumulation and DNA binding of trans-bis-(2-hydroxypyridine)2dichloroplatinum(II) coded as DH3.
Materials and Methods
Materials. Potassium tetrachloroplatinate K2[PtCl4] and N,N-dimethylformamide (DMF) (Sigma Chemical Company, St. Louis, USA); thiazole (Sigma-Aldrich Chemie GmbH Germany); acetone and silver nitrate (Ajax Chemicals, Auburn, NSW, Australia); methanol and ethanol (Merck Pty. Limited, Kilsyth, Vic., Australia); pBR322 plasmid DNA (ICN Biomedicals, OH, USA); foetal calf serum (FCS), 5× RPMI-1640, 200 mM L-glutamine, and 5.6% sodium bicarbonate (Trace Biosciences Pty Ltd Sydney Australia), commercially available JETQUICK Blood DNA Spin Kit/50 (Astral Scientific Australia) used to isolate high molecular weight DNA from cell pellets.
Synthesis of DH3. To 1 mmol (0.415 g) of potassium tetrachloroplatinate dissolved in 4 ml of milli Q (mQ) water was added 0.25 ml of concentrated hydrochloric acid. The solution was heated to 50°C to which was slowly added 20 mmol (1.9 g) of 2-hydroxypyridine dissolved in 2 ml of DMF. The temperature of the solution was increased to approximately 65°C and maintained at that temperature for 1 h while the solution was stirred. The volume of the solution was reduced to about 5 ml to which was added 40 ml of 6 M hydrochloric acid. The mixture was heated under reflux for 24 h at 90°C, following which it was cooled to room temperature. The yellow precipitate of DH3 formed was collected by filtration at the pump, washed with ice-cold water and ice-cold ethanol and air dried, giving a yield of 60%.
Characterization
Microanalyses: Platinum was determined by graphite furnace atomic absorption spectroscopy (AAS) using a Varian Spectra-240 Atomic Absorption Spectrophotometer at the University of Sydney. C, H, N, and Cl were determined at the Australian National University. DH3: Calc.% C: 26.3; H: 2.2, N: 6.1; Cl: 15.6; Pt: 42.8. Obs.%: C: 26.3±0.4, H: 2.2±0.4, N: 6.1±0.4, Cl: 15.9±0.4, Pt: 42.8±1.0.
Molar conductivity: The limiting molar conductivity value (in Ω−1 cm2 mol−1) at zero concentration (Λ0) for DH3 and cisplatin are 730 and 270, respectively, indicating DH3 is more fully ionized than cisplatin in solution in the 1:10 (v:v) mixture of DMF and mQ water. Spectral studies: Infrared, mass and 1H-NMR) spectra were used to aid in the structural characterization of DH3.
Infra-red (IR) Spectral analysis: (KBr). IR spectrum was collected using a Bruker IFS66 spectrometer equipped with a Spectra- Tech Diffuse Reflectance Accessory (DRA) (Bruker Australia Pty Ltd Bruker Sydney NSW Australia), an air-cooled DTGS detector and a KBr beam splitter. Spectrum was recorded at a resolution of 4 cm−1, with the addition of 128 scans and a Blackman-Harris 3-term apodization function was applied. The peaks listed below can be seen to provide support for the suggested structure of DH3. In particular it indicates the presence of various functional groups. 3245.4 cm−1: N-H stretch; 1597.4, 1538.4, 1403.8 cm−1: C=N and C=C stretch; 1239.9 cm−1: C–N stretch; 920.0 cm-1: pyridine ring stretch; 983.4 cm−1: C–H in-plane bending; 818.0, 503.3 cm−1: C–H out-of-plane bending; 473.7, 449.0 cm−1: Pt–N stretch, 239.8 cm-1: Pt–Cl stretch.
Mass spectroscopy: The mass spectrum of DH3 was obtained using a Finnigan LCQ ion trap mass spectrometer (Thermo Scientific Massachusetts USA) in which fragmentation was produced by electrospray ionization (EIS). EIS-MS (mQ water) (m/z: M=378). The peak at m/z = 383 corresponds to (M–2Cl–2H) and the peak at m/z=415 corresponds to (Pt(2-hydroxypyridine)2Cl–2H) and the peak at m/x=288 corresponds to (Pt(2-hydroxypyridine)–2H).
1H-NMR: 1H-NMR spectrum of DH3 dissolved in dimethylsulfoxide (DMSO) was recorded in a Bruker DPX400 spectrometer (Bruker Australia Pty Ltd Sydney NSW Australia) using a 5 mm high-precision Wilmad NMR tube at 300. 1H NMR DMSO δ ppm: 8.50 (s, due to OH); 7.94 (d, due to ortho CH); 4.50 (d, due to meta CH); 3.50 (q, due to meta CH); 2.69 (q, due to para CH); NH–Pt); 2.55 (d, due to dimethylsulfoxide (DMSO)).
Cytotoxicity assays. Cytotoxicity of DH3 against human ovarian A2780, A2780cisR, and A2780ZD0473R cancer cell lines was determined using the 3-(Dimethyl-2-thiazolyl)-2,5-diphenyl-2H-terazolium bromide (MTT) reduction assay (14, 15). Cisplatin was used as a reference. A2780 (parent), A2780cisR (cisplatin-resistant type) and A2780ZD0473R (ZD0473-resistant type) ovarian cancer cell lines were obtained from Ms. Zhang from Royal Prince Alfred Hospital, University of Sydney, Australia. Briefly, between 4500 to 5500 cells were seeded into the wells of the flat-bottomed 96-well culture plate in 10% FCS/RPMI 1640 culture medium. The plate was incubated for 24 h at 37°C in a humidified atmosphere to allow cells to attach. Platinum complexes dissolved first in a minimum volume of DMF were diluted to the required concentrations with mQ water and filtered to sterilise. Serial dilutions ranging from 0.0064 μM to 0.8 μM in 10% FCS/RPMI 1640 medium were prepared and added to equal volumes of cell culture in quadruplicate wells. The plates were left to incubate under normal growth conditions for 72 h. The inhibition of cell growth was determined using the MTT assay (15). Four hours after the addition of MTT (50 μl per mg ml−1 MTT), the yellow formazan crystals produced were dissolved in 150 μl of DMSO and the absorbance of the resulting solution was read with Bio-Rad Model 3550 Microplate Reader (BioRad Sydney Australia) set at 570 nm.
Platinum accumulation and platinum–DNA binding. DH3 and cisplatin (at 50 μM final concentration) were added to culture plates containing exponentially growing A2780, A2780cisR and A2780ZD0473R cells in 10 ml 10% FCS/RPMI 1640 culture medium (cell density=1×106 cells ml−1). The cells containing the drugs were incubated for periods of 2 h, 4 h and 24 h at the end of which cell monolayers were trypsinized and cell suspension (5 ml) was transferred to centrifuge tubes and spun at 3500 rpm for 2 min at 4°C (16). Ice-cold phosphate-buffered saline (PBS) was used to wash the cells twice and the pellets were stored at −20°C until assayed. At least three independent experiments were performed.
For the determination of drug accumulation in the cells, cell pellets were suspended in 0.5 ml of 1% triton-X, held on ice then sonicated (5 min). Total intracellular platinum contents were determined by graphite furnace AAS.
For the determination of the level of platinum–DNA binding, the modified method of Bowtell (17) was used. Briefly, high molecular weight DNA was isolated from cell pellets using JETQUICK Blood DNA Spin Kit/50. The cell pellets were resuspended in PBS to a final volume of 200 μl and mixed with 10 μl of RNase A, then incubated for 4 min at 37°C. 25 μl Proteinase K and 200 μl Buffer K1 (containing guanidine hydrochloride and a detergent) were added to the mixture followed by incubation for 10 min at 70°C. Absolute ethanol (200 μl) was added and mixed thoroughly to prevent any precipitation of nucleic acids due to high local alcohol concentration. The samples were centrifuged for 1 min at 10600 rpm through the silica membrane using JETQUICK micro-spin columns. The columns containing the samples were washed with 500 μl of buffer KX (containing high-salt buffer to remove residual contamination), then centrifuged for 1 min at 10,600 rpm. The columns were washed again with 500 μl K2, low-salt buffer to change the high-salt conditions on the silica membrane to low-salt, and centrifuged for 1 min at 10600 rpm. The columns were centrifuged again for 2 min at full speed (13000 rpm) to further clear the silica membrane of residual liquid. The receivers were changed and the purified DNA in the column was eluted from the membrane with 200 μl of 10 mM Tris-HCl buffer (pH 8.5). DNA content was determined using UV spectrophotometry set at 260 nm (Varian Cary 1E UV-Visible spectrometer with Varian Cary Temperature Controller). Platinum levels were determined by graphite furnace AAS. A260/A280 ratios were found to be between 1.75 and 1.8 for all samples, reflecting high purity of the DNA.
IC50 values (μM±SD) and resistance factors (RF) for DH3 and cisplatin as applied to the human ovarian A2780, A2780cisR and A2780ZD0473R cancer cell lines.
Interaction with pBR322 plasmid DNA. Interaction between DH3 and cisplatin with pBR322 plasmid DNA, with and without BamH1 digestion, was studied using gel electrophoresis using a method described by Stellwagen (18). Exactly 1.5 μl of supplied pBR322 plasmid DNA in solution was added to solutions of DH3 and cisplatin at different concentrations ranging from 0.55 μM to 70 μM. The DNA blank was prepared by adding 18.5 μl mQ water to 1.5 μl of pBR322 plasmid DNA. The samples and the DNA blank were incubated for 4 h on a shaking water bath at 37°C. The reaction was stopped by rapid cooling to 0°C for 20 min. The samples were mixed with 1 μl of marker dye ethidium bromide and 16 μl of each sample was loaded onto 1% agarose gel made in (TAE) buffer. The gel was photographed using Eastman Kodak Company, Molecular Imaging Systems (Carestream Health Inc. Rochester New York USA).
BamH1 digestion. BamHI is a type II restriction endonuclease that hydrolyses phosphodiester bonds. It binds at the recognition sequence 5’-GGATCC-3’, and chops these sequences just after the 5’-guanine on each strand (19). pBR322 plasmid DNA contains a single restriction site for BamH1 that converts the supercoiled form I and singly nicked circular form II to linear form III DNA. The same identical set of drug-DNA mixtures as described previously, was first incubated for 4 h on a shaking water bath at 37°C and then subjected to BamH1 (10 units μl−1) digestion. To each 20 μl of the incubated drug-DNA mixture was added 2 μl of 10× digestion buffer SB first and then 0.1 μl BamH1 (1 unit). The mixtures were left in a shaking water bath at 37°C for another 1 h at the end of which the reaction was terminated by rapid cooling and 4 μl of ethidium bromide was added to each sample before loading onto the gel. The gel was photographed following the method described previously.
Results
Cytotoxicity. Table I lists the (IC50) values and resistance factors (RF) for QH3 and cisplatin applying to the human ovarian A2780, A2780cisR and A2780ZD0473R cancer cell lines. IC50 value is defined as the drug concentration required for 50% cell kill and resistance factor (RF) is defined as the ratio of the IC50 value in the resistant cell line over that in the parent cell line.
Pt accumulation (nmol Pt per 2×106 cells) in A2780, A2780cisR and A2780ZD0473R cell lines in 2, 4 and 24 h after treatment with 50 μM cisplain and DH3.
Levels of platinum–DNA binding (nmol platinum per mg of DNA) in A2780 cell line in 2, 4 and 24 h after treatment with 50 μM DH3 and cisplatin.
The results show that DH3 is less active than cisplatin in A2780 and A2780ZD0473R cell lines but more active than cisplatin in A2780cisR.
Platinum accumulation and platinum–DNA binding. Table II gives the total intracellular platinum levels found in the A2780, A2780cisR and A2780ZD0473R cell lines after exposure to 50 μM concentration of DH3 and cisplatin for 2, 4 and 24 h. Table III gives the levels of platinum–DNA binding in A2780 cell line in 2, 4 and 24 h from DH3 and cisplatin. DH3 has higher cellular accumulations of platinum in the A2780 and A2780cisR cell lines at all time points, whereas cisplatin has the higher values in the A2780ZD0473R cell line at 4 h and 24 h. DH3 produces higher levels of platinum–DNA binding than cisplatin at all the time points in the cell lines A2780 and A2780ZD0473R even though the compound is less active than cisplatin against A2780 and A2780ZD0473R cell lines.
Interaction with pBR322 plasmid DNA. Figure 2 shows the electrophoretograms applying to the interaction of a fixed amount of pBR322 plasmid DNA with increasing concentrations of DH3 and cisplatin.

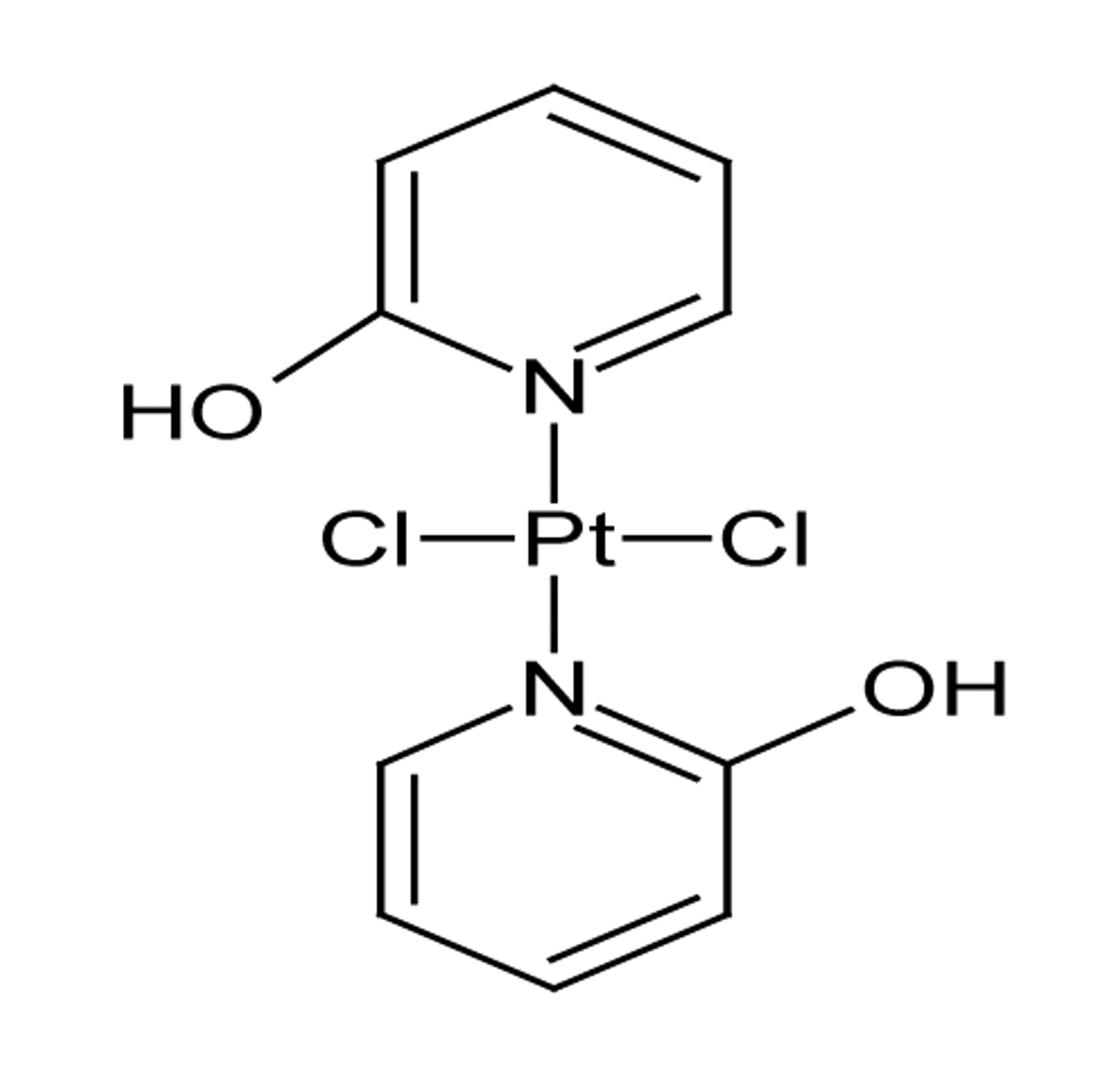
Structure of DH3 [trans-bis(2-hydroxypyridine)dichloroplatinum(II)].
As pBR322 plasmid DNA at a fixed concentration was treated with increasing concentrations of DH3 and cisplatin, a decrease in mobility of the bands was observed in the case of DH3 but there was a significant increase in the mobility of form I band in the case of cisplatin.
BamH1 digestion. BamH1 is a restriction endonuclease that recognizes the sequence G/GATCC and hydrolyses the phosphodiester bond between adjacent GG sites. pBR322 plasmid DNA contains a single restriction site for BamH1 that converts the supercoiled form I and also singly nicked circular form II to linear form III DNA. Thus, when untreated pBR322 plasmid DNA was digested with BamH1, only one band corresponding to form III was observed whereas in the untreated and undigested pBR322 plasmid DNA, generally two bands corresponding to forms I and II were observed (see Figure 2). When the pBR322 plasmid DNA interacted with increasing concentrations of DH3 followed by BamH1 digestion, forms I, II and III bands were observed at concentrations ranging from 10 μM to 35 μM of DH3 (electrophoretograms not shown).
Discussion
Structure. The structure of DH3 could not be confirmed by single crystal x-ray diffractometry as no suitable single crystal was available. However, the results of elemental analyses and spectral studies (described earlier) can be seen to provide support for the suggested structure of the compound (Figure 1).
Activity. It can be seen that DH3 is less active than cisplatin against the A2780 and A2780ZD0473R cell lines but more active than cisplatin against the A2780cisR cell line. However, DH3 has much lower RFs than cisplatin, indicating a smaller difference in activity between the parent and the resistant cell lines for the compound than cisplatin. It has been previously reported that trans-planaramine-platinum(II) complexes generally have lower RFs for A2780 and A2780cisR cell lines than cisplatin (20).
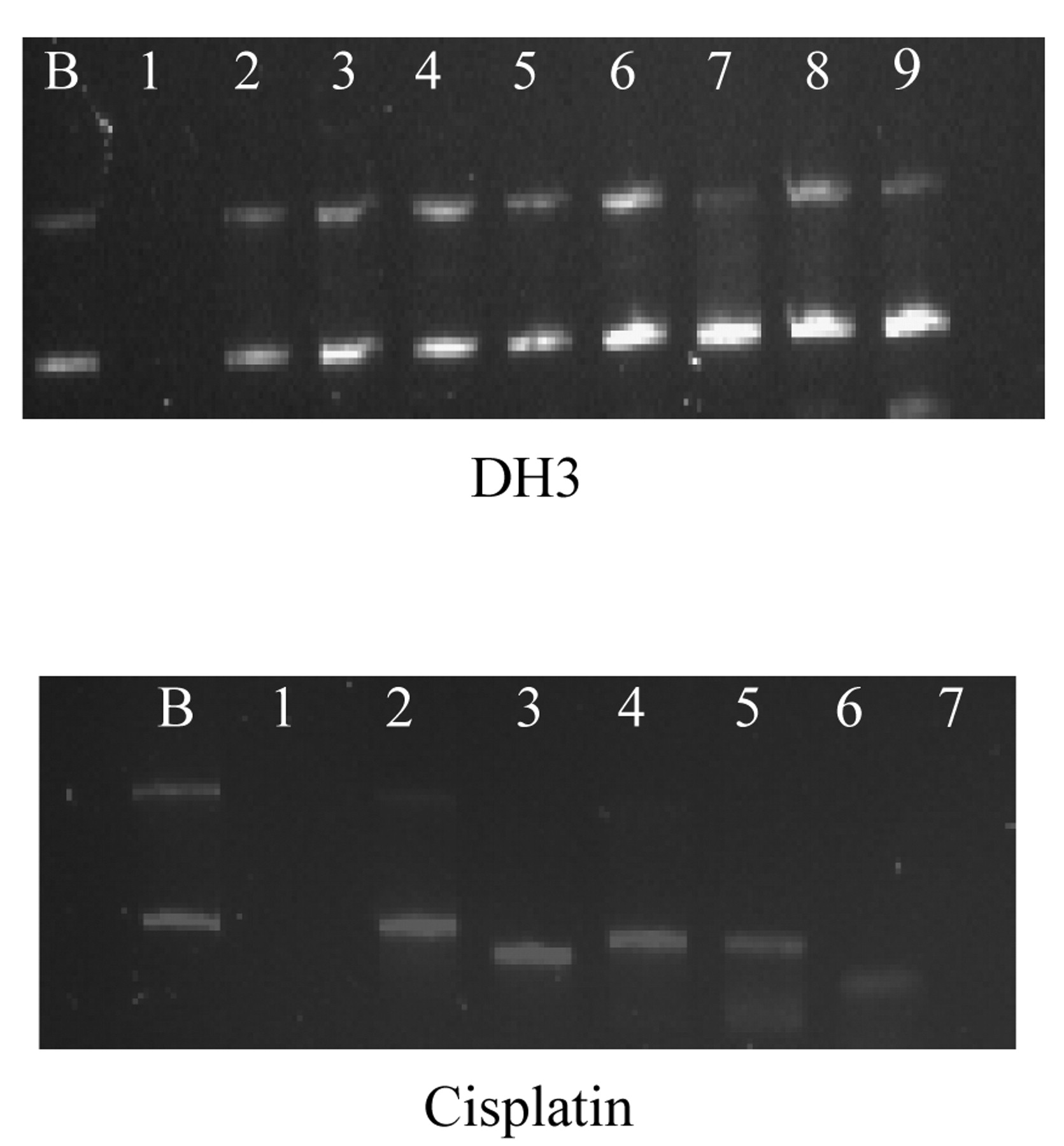
Electrophoretograms applying to the interaction of a fixed amount of pBR322 plasmid DNA with increasing concentrations of DH3 and cisplatin. Lane B: Untreated pBR322 plasmid DNA to serve as a control, lanes 1 to 8: plasmid DNA treated with increasing concentrations of QH3 and cisplatin, lane 1: 0.55 μM, lane 2: 1.09 μM, lane 3: 2.19 μM, lane 4: 4.38 μM, lane 5: 8.75 μM, lane 6: 17.50 μM, lane 7: 35 μM, and lane 8: 70 μM.
Platinum accumulation. DH3 has higher cellular accumulations of platinum in the A2780 and A2780cisR cell lines at all time points, whereas cisplatin has the higher values in the A2780ZD0473R cell line at 4 h and 24 h. The results illustrate that cellular accumulation of platinum per se may not be an indicator of the activity of compounds, since the compounds may be deactivated by cellular thiols such as glutathione before binding with DNA. The results may also be seen to support the idea that increased deactivation within the cell may be a dominant mechanism of resistance operating in the A2780ZD0473R cell line. Another point to note is that whereas cisplatin binds with nucleobases in DNA to form mainly bifunctional intrastrand 1,2-Pt(GG) and 1,2-Pt(AG) adducts that cause local bending of a DNA strand, DH3 would form mainly monofunctional Pt(G) and bifunctional interstrand Pt(GG) adducts, the latter causing more of a global change in the DNA conformation.
Platinum–DNA binding. As shown in table III, DH3 produces higher levels of platinum–DNA binding than cisplatin at all the time points in the cell lines A2780 and A2780ZD0473R, even though the compound is less active than cisplatin against the A2780 and A2780ZD0473R cell lines. Conversely, even though DH3 is more active than cisplatin against the A2780cisR cell line, it is associated with lower level of platinum–DNA binding than cisplatin. The results can be seen to illustrate that even the platinum–DNA binding per se may not necessarily be a true measure of the activity of platinum compounds. It may be noted that platinum–DNA binding is necessary but not sufficient to bring about apoptosis, as the subsequent cell death is actually caused by downstream processes involving many proteins. It is hypothesized that non-critical adducts, such as monofunctional Pt(G) and DNA-protein crosslinks, may predominate in the case of DH3 in the A2780 and A2780ZD0473R cell lines.
Interaction with pBR322 plasmid DNA. pBR322 plasmid DNA can exist in three forms, namely supercoiled form I, singly nicked relaxed circular form II and doubly nicked linear form III. The supercoiled form I has the fastest rate of migration, whereas the singly nicked circular form II has the lowest migration rate. As pBR322 plasmid DNA at a fixed concentration interacted with increasing concentrations of DH3 and cisplatin, a decrease in mobility of the bands was observed in the case of DH3 but there was a significant increase in the mobility of form I in the case of cisplatin. A marked decrease in the intensity of the band was also observed at high concentrations of cisplatin. The change in mobility of the DNA bands was indicative of conformational change in the DNA, whereas the decrease in intensity indicated the occurrence of DNA damage. In the case of DH3, non-covalent interactions, such as hydrogen bonding involving 2-hydroxypyridine ligand, are believed to play a key role in bringing about DNA conformational change as well as DNA damage.
BamH1 digestion. As stated earlier, when the pBR322 plasmid DNA interacted with increasing concentrations of DH3 followed by BamH1 digestion, form I, II and III bands were observed at concentrations ranging from 10 μM to 35 μM of DH3. The appearance of only form III band in lanes 2 and 6 at 10 μM and 30 μM concentrations are believed to be an artefact. The results indicate that the binding of DH1 with pBR322 plasmid DNA partially prevents BamH1 digestion of the DNA, indicating changes in DNA conformation brought about by covalent and non-covalent interactions between DH1 and nucleobases in the DNA. In the case of cisplatin, form I and II bands were observed at all concentrations ranging from 5 μM to 50 μM. The results indicated that cisplatin led to greater prevention of BamH1 digestion than did DH3, indicative of greater conformational changes in the DNA.
Finally, a new trans-planaramineplatinum(II) was synthesized here and investigated for activity against human ovarian tumour models and nature of binding with pBR322 plasmid DNA. The lower activity of DH3 as compared to cisplatin suggests that this compound may not offer any advantage over cisplatin as a potential anticancer drug.
Acknowledgements
This work was partly funded by a Biomedical Science Research Initiative Grant and Biomedical Science Cancer Research Donation Fund.
Footnotes
-
Conflict of Interest
Narjes Deqnah, Jun Qing Yu, Philip Beale, Keith Fisher and Fazlul Huq declare that they have no financial and personal relationships with other people or organizations that could inappropriately influence their work.
- Received October 9, 2011.
- Revision received December 8, 2011.
- Accepted December 9, 2011.
- Copyright© 2012 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.