


Abstract
The widely used anticancer drug cisplatin (CS) is believed to cross the cell membrane by passive diffusion, carrier-mediated transport and pinocytosis. One carrier involved in the transport of CS into the cell is the copper transporter CTR1. However, CS is found to trigger the down-regulation of CTR1 and its proteasomal degradation. The proteasome inhibitor bortezomib (Bort) has been reported to block CS-induced down-regulation of CTR1 so that in the presence of Bort, the cellular uptake of CS may be increased. Increased platinum accumulation may result in increased platinum–DNA binding so that CS in combination with Bort may produce pronounced cell kill. In this study, synergism in activity from the sequenced combination of CS and Bort in human ovarian A2780, A2780cisR and A2780ZD0473R cancer cell lines was studied. We also investigated the effect on cell kill due to the administration of CS in two aliquots with a time gap. Addition of Bort 2 h before CS was found to produce greater cell kill than the converse and the bolus, especially in the resistant A2780cisR and A2780ZD0473R cell lines, in line with increased platinum accumulation and platinum–DNA binding levels. Thus, the prevention of CTR1 degradation by Bort may play a significant role, especially in the resistant cell lines. Administration of CS in two aliquots with a time gap was also found to maximise the cell kill in the ovarian cancer cell lines. If such findings are found to be true in vivo, the results may be significant clinically.
Although cisplatin (CS) is a widely used anticancer drug, its use has also been limited due to intrinsic and/or acquired resistance. One of the mechanisms of CS-resistance relates to the reduced cellular platinum accumulation that may be due to reduced intake and/or increased efflux of the drug (1-3). Besides passive diffusion and pinocytosis, CS is believed to enter cells by carrier-mediated transport, such as that via the copper transporter CTR1 (4-7). However, CS is found to trigger the down-regulation of CTR1 and its proteasomal degradation (8). The present study aimed to determine the effect on cell kill and cellular uptake of CS due to its combination with the proteasome inhibitor bortezomib (Bort). We also determined the same associated with the combination of a designed trans-planaramine-platinum(II) coded as YH12 with Bort (9). A separate aim of the study was to determine the effect on cell kill due to the administration of CS in two aliquots with a time gap. This question arose from the fact that drugs with different mechanisms of action may act synergistically in combination (thus offering a means of overcoming drug resistance) and that CS brings about apoptosis by multiple pathways (10-13). It is thought that different pathways of CS action may come into play at different time points (or at least play a more prominent role) depending on the status of the cell (e.g. the presence or absence of oxidative stress) so that the administration of the drug in two aliquots with a time gap may be looked at as the combination of two drugs with somewhat different mechanisms of action.
Materials and Methods
Materials. CS and YH12 were prepared according to previously described methods (9, 14). Bort was purchased from LC Laboratories Woburn, MA, USA. Foetal calf serum (FCS), RPMI-1640, 200 mM L-glutamine, and 5.6% sodium bicarbonate were obtained from Trace Biosciences Pty Ltd., Australia. Other chemicals were mostly purchased from Sigma-Aldrich, Sydney, Australia. A2780, A2780cisR and A2780ZD0473R ovarian cancer cell lines were gifts from Ms. Mei Zhang, Royal Prince Alfred Hospital, Sydney, Australia. Stock solutions of platinum compounds (1 mM) were prepared in 1:1 DMF-mQ water mixture and that of Bort (1 mM) were made in ethanol.
Cell culture. Human ovarian cancer cell lines A2780, its cisplatin-resistant form A2780cisR and its ZD0473-resistant form A2780ZD0473R were seeded in 25 cm2 tissue culture flasks in an incubator at 37°C in a humidified atmosphere consisting of 5% CO2 and 95% air. The cells were maintained in logarithmic growth phase in complete medium consisting of RPMI-1640, 10% heat inactivated FCS, 20 mM Hepes, 0.11% bicarbonate, and 2 mM glutamine without antibiotics (15). Each cell line was seeded at a density of 4,000 and 6,000 cells/well in a flat-bottomed 96-well culture plate in 10% FCS/RPMI-1640 culture medium. The plate was then incubated for 24 h at 37°C in a humidified atmosphere to allow cells to attach.
Single-drug treatment. For single-drug treatment, each of the stock solutions was subjected to serial dilutions to give final concentrations ranging from 0.0008 to 20 μM. The dilutions were performed using 10% RMPI-1640 medium without serum as the vehicle and were added to equal volumes of cell culture in triplicate wells, then cells were left to incubate for 72 h. These treatments were carried out to determine (IC50) values i.e. drug concentrations required for 50% cell kill.
Combination analysis. Dose-response curves and combination indices (CIs) were used as measures of synergism and generated/calculated using the program CalcuSyn (16-18). The CI for binary combinations of drugs was calculated according to the equation:
 where D1 and D2 represent mean doses of compounds 1 and 2 respectively in combination required to cause x% inhibition, whereas D1x and D2x represent the doses of compounds 1 and 2 respectively required to cause x% inhibition when present alone. Dx can be readily calculated from the following form of median effect equation:
where D1 and D2 represent mean doses of compounds 1 and 2 respectively in combination required to cause x% inhibition, whereas D1x and D2x represent the doses of compounds 1 and 2 respectively required to cause x% inhibition when present alone. Dx can be readily calculated from the following form of median effect equation:
 where Dx denotes the dose of drug, Dm is the median-effect dose, fa is the fraction of cells affected so that fu=1−fa and m is the exponent defining the shape of the dose–effect curve. CI values of <1, 1 and >1 indicate synergism, additivity and antagonism in combined drug action, respectively. Recently, Huq defined a new term called the ‘enhancement factor’ (EF) giving a direct relationship between EF and synergism (10).
where Dx denotes the dose of drug, Dm is the median-effect dose, fa is the fraction of cells affected so that fu=1−fa and m is the exponent defining the shape of the dose–effect curve. CI values of <1, 1 and >1 indicate synergism, additivity and antagonism in combined drug action, respectively. Recently, Huq defined a new term called the ‘enhancement factor’ (EF) giving a direct relationship between EF and synergism (10).
Platinum cellular accumulation and platinum–DNA binding studies. As the action of platinum-containing drugs is associated with their binding with DNA, cellular accumulation of platinum and platinum–DNA binding levels in A2780 and A2780cisR cell lines were determined for combination of CS-Bort applying to the sequences 0/0 h and 0/2 h. Furthermore, the platinum content in cells treated with CS in bolus and CS in two aliquots with a time gap (2 h and 4 h) were also determined. Combinations of CS with Bort (at IC50 values) and the addition of CS in two aliquots (at 50 mM final concentration) were added to culture plates containing exponentially growing A2780 and A2780cisR cells in 10 ml 10% FCS/RPMI-1640 culture medium (cell density=1×106 cells/ml). The cells containing the drugs were incubated for 24 h at the end of which cell monolayers were trypsinized and cell suspensions (10 ml) were transferred to centrifuge tubes and spun at 3,500 rpm for 2 min at 4°C. The cells were washed twice with ice-cold phosphate-buffered saline (PBS) and the pellets were stored at −20°C until assayed. At least three independent experiments were performed.
Cellular accumulation. Following drug incubation, the cell pellets were suspended in 0.5 ml 1% Triton-X, held on ice then sonicated. Total intracellular platinum contents were determined by graphite furnace atomic absorption spectrophotometry.
Drug–DNA binding. Following drug incubation, high molecular weight DNA was isolated from the cell pellet using H440050 JETQUICK Blood DNA Spin Kit/50 from Austral Scientific Pty Ltd., Sydney, Australia, and the modified protocol of Bowtell (19). The cell pellets, resuspended in PBS to a final volume of 200 μl and mixed with 10 μl of RNase A, were incubated for 4 min at 37°C. About 25 μl Proteinase K and 200 μl Buffer K1 (containing guanidine hydrochloride and a detergent) were added to the mixture followed by incubation for 10 min at 70°C. Then 200 μl of absolute ethanol was added and mixed thoroughly to prevent any precipitation of nucleic acids. The samples were centrifuged for 1 min at 10,600 rpm through the silica membrane using JETQUICK micro-spin column. The columns containing the samples were washed with 500 μl of buffer KX (containing high-salt buffer to remove residual contaminations) and centrifuged for 1 min at 10,600 rpm. These were again washed with 500 μl buffer K2 (containing low-salt buffer to change the high-salt conditions on the silica membrane to low-salt) and centrifuged for 1 min at 10,600 rpm. To further clear the silica membrane from residual liquid, the sample columns were centrifuged again for 2 min at full speed (13,000 rpm). The column receivers were changed and the purified DNA in the column was eluted from the membrane with 200 μl of 10 mM Tris–HCl buffer (pH 8.5). DNA content was determined by UV spectrophotometry (260 nm) and platinum levels were determined by graphite furnace AAS.
Results
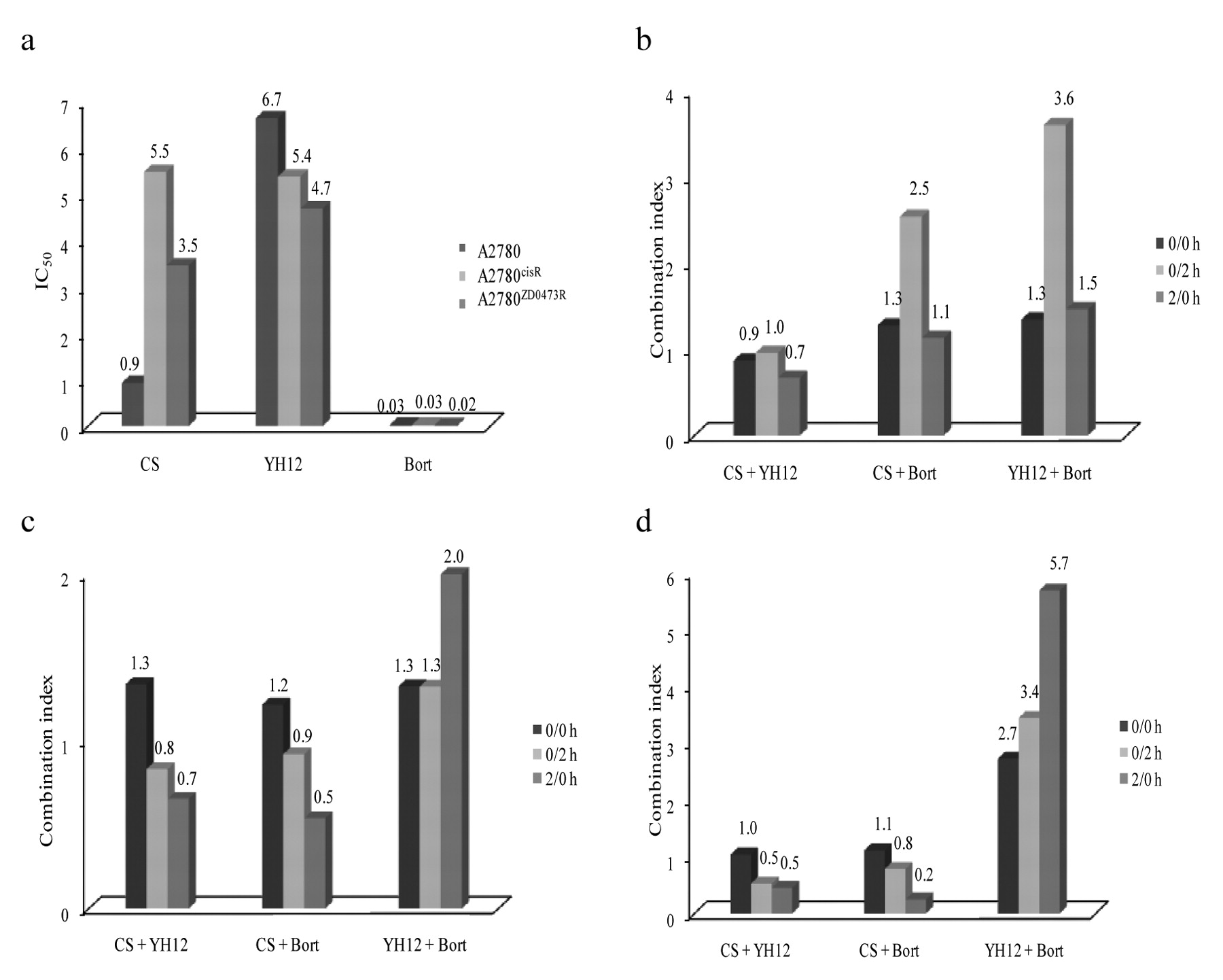
Cytotoxicity. Figure 1a gives the IC50 values of the compounds for the A2780, A2780cisR and A2780ZD0473R cell lines. Whereas the IC50 values for CS increase in going from the parent A2780 cell line to the resistant A2780cisR and A2780ZD0473R cell lines, the values for YH12 and Bort were found to decrease or stay nearly the same. Bort was found to be 30 times more active than CS against the parental A2780 cell line and about 180 times more active against A2780cisR and A2780ZD0473R cell lines.
Combination studies. Figure 1b, c and d show the CIs for the combinations of CS and YH12 with Bort in A2780, A2780cisR and A2780ZD0473R cell lines. For the combinations of CS with YH12 in the A2780 cell line, the 0/0 h combination was slightly synergistic, the 0/2 h sequence was additive, whereas the 2/0 h sequence was most synergistic. In the A2780cisR and A2780ZD0473R cell lines, 2/0 h combination of CS with YH12 was found to be most synergistic, whereas the 0/0 h combination was slightly antagonistic. For the combinations of CS with Bort in the A2780 cell line, all three sequences of administration were found to be antagonistic, with the 0/2 h sequence being the most antagonistic. In the A2780cisR and A2780ZD0473R cell lines, 0/2 h and 2/0 h combinations of CS with Bort were found to be slightly synergistic, whereas the 0/0 h combination was slightly antagonistic. For the combinations of YH12 with Bort, all the three sequences of administration were found to be antagonistic in all the three human ovarian cell lines.

a: IC50 (μM) values of CS, YH12 and Bort for human ovarian cancer cell lines: A2780, A2780cisR and A2780ZD0473R; b: Combination indices for combinations of CS and YH12 with Bort in A2780 ovarian cancer cell line; c: Combination indices for combinations of CS and YH12 with Bort in A2780cisR ovarian cancer cell line; d: Combination indices for combinations of CS, YH12 and Bort in A2780ZD0473R ovarian cancer cell line.
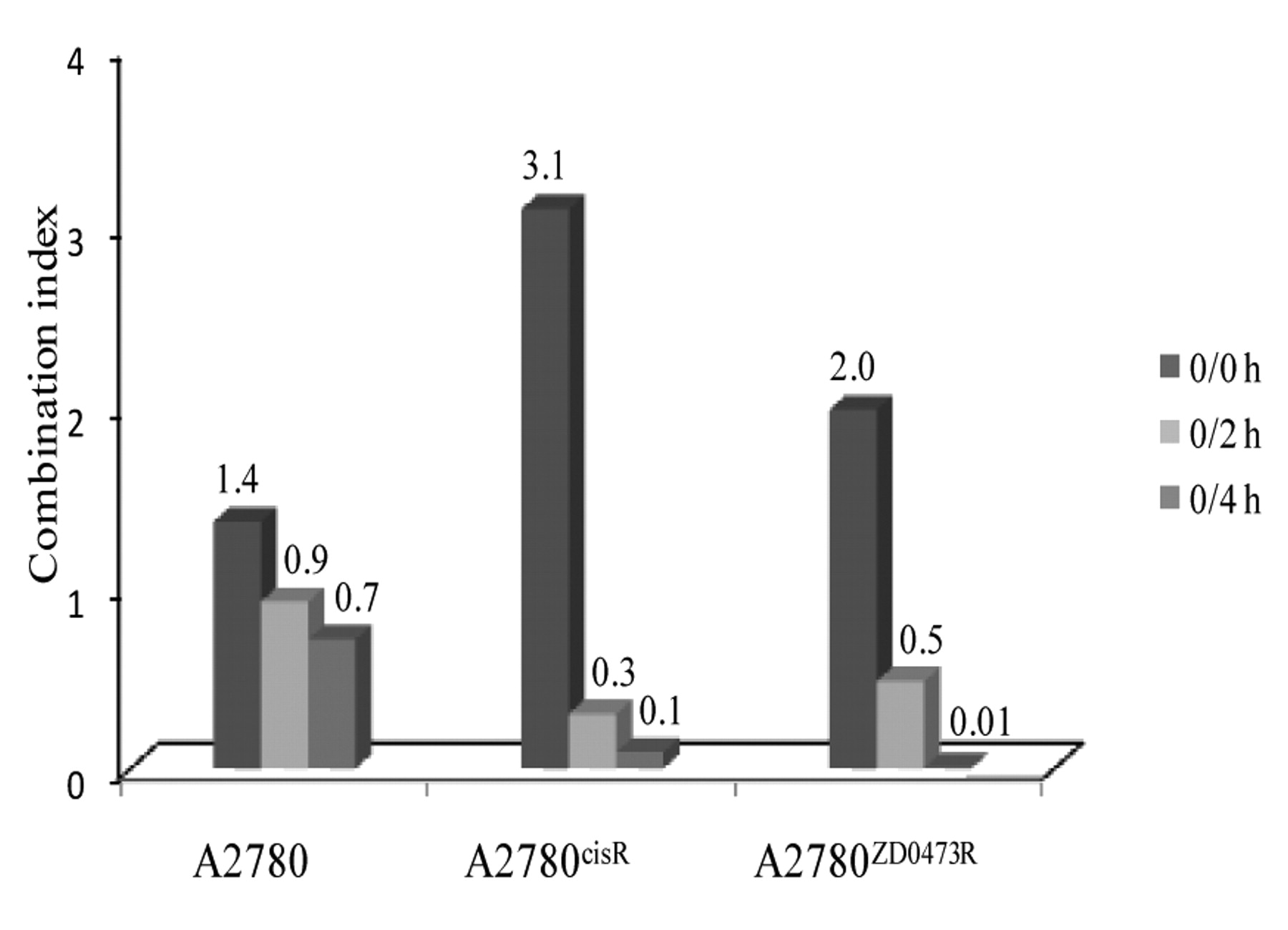
Effect of CS given as a bolus and in two aliquots. Figure 2 shows the CIs for administration of CS as a bolus (0/0 h), and in two aliquots with 2 h and 4 h time gaps (0/2 h and 0/4 h sequences of administration). The CIs were calculated based on the assumption that the combined action of the drug is the same as that resulting from the combination of two drugs. It can be seen that CS caused a greater cell kill in all three human ovarian cell lines when given in two aliquots rather than as a bolus and that a 4 h time gap is better than a 2 h time gap. The effect of sequence of administration was found to be more pronounced in the two resistant A2780cisR and A2780ZD0473R cell lines (more so in A2780cisR) than in the parental A2780 cell line.

The administration of CS in two aliquots using 2 h and 4 h time gaps to A2780, A2780cisR and A2780ZD0473R cell lines.

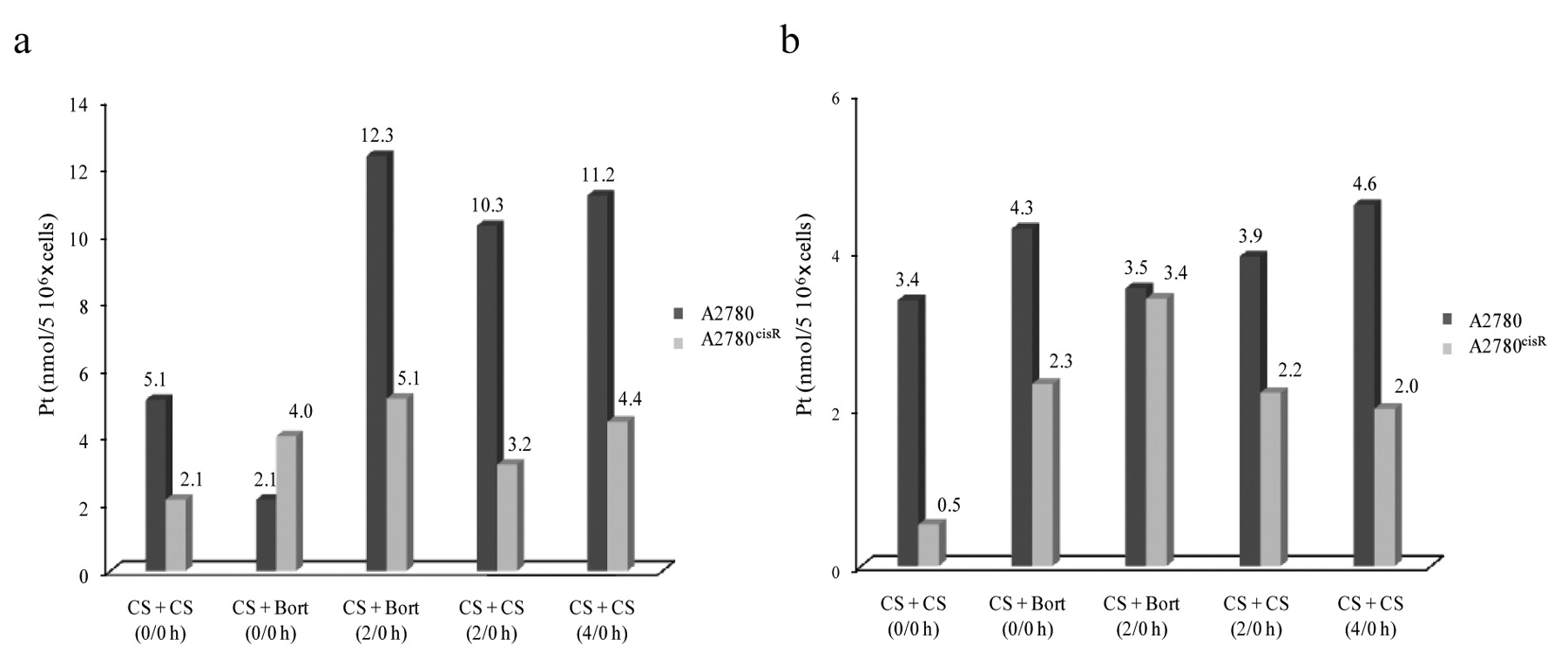
a: Total intracellular platinum accumulation levels found in the cell lines A2780 and A2780cisR. b: Platinum–DNA binding levels in A2780 and A2780cisR cell lines.
Platinum accumulation. Figure 3a shows the platinum accumulation in A2780 and A2780cisR cell lines treated with CS alone, with the 0/0 h and 2/0 h combinations of CS with Bort, and with CS in two aliquots with 2 h and 4 h time gaps. It was found that with CS alone, the cellular accumulation of platinum was greater in the parental A2780 cell line than in the resistant A2780cisR cell line, irrespective of whether it was administered as a bolus or in two aliquots with a time gap (2 h or 4 h). When given in two aliquots with a time gap, the 4 h time gap was found to result in greater platinum accumulation than the 2 h time gap. For the combination of CS with Bort, the 2/0 h sequence of administration was found to produce greater intracellular platinum accumulation than the 0/0 h sequence of administration for both the parental A2780 and the resistant A2780cisR cell lines.
Platinum–DNA binding. Figure 3b shows the platinum–DNA binding levels in A2780 and A2780cisR cell lines resulting from treatment with CS alone, with the 0/0 h and 2/0 h combinations of CS with Bort, and with the administration of CS in two aliquots with 2 h and 4 h time gaps. For treatment with CS alone, the platinum–DNA binding level was found to be greater in the parental A2780 cell line than in the resistant A2780cisR cell line, irrespective of whether it was given as a bolus or in two aliquots with a time gap (2 h or 4 h). For administration of CS in two aliquots with a time gap, 4 h time gap was found to be associated with a greatest level of platinum–DNA binding level than the 2 h time gap in the A2780 cell line in line with higher level of cellular accumulation of platinum. However, in the A2780cisR cell line, platinum–DNA binding level from the 4 h time gap was found to be slightly lower than that from the 2 h time gap, even though the converse was found for the cellular accumulation of platinum.
Discussion
In this study, synergism in activity (or changes in cell kill) from the combinations of CS and YH12 with Bort in the human ovarian A2780, A2780cisR and A2780ZD0473R cell lines was investigated as a function of the sequence of administration. Experiments on the combination of CS with YH12 were repeated to ascertain the agreement of the results from the present study with the reported values (11). For the combinations of CS with YH12 in A2780 and A2780cisR and A2780ZD0473R cell lines, the 2/0 h sequence of administration was found to be most synergistic, whereas the 0/0 h and 0/2 h sequences were found to be additive to slightly antagonistic. The results are essentially in line with the reported values in which the 4/0 h combination of CS with YH12 was found to be most synergistic, whereas the addition of CS first followed by YH12 4 h later produced the least synergistic outcomes (11). It was suggested that as CS binds with a DNA strand forming intrastrand bifunctional 1,2-Pt(GG) and 1,2-Pt(AG) adducts, the DNA strand becomes bent. As a result, the subsequent interstrand bifunctional binding of YH12 to the bent DNA is hampered due to a greater distance mismatch between the two trans-arms of YH12 and the distance between 1,2-interstrand N7(G) and N7(G) positions especially those close to the CS-binding site.
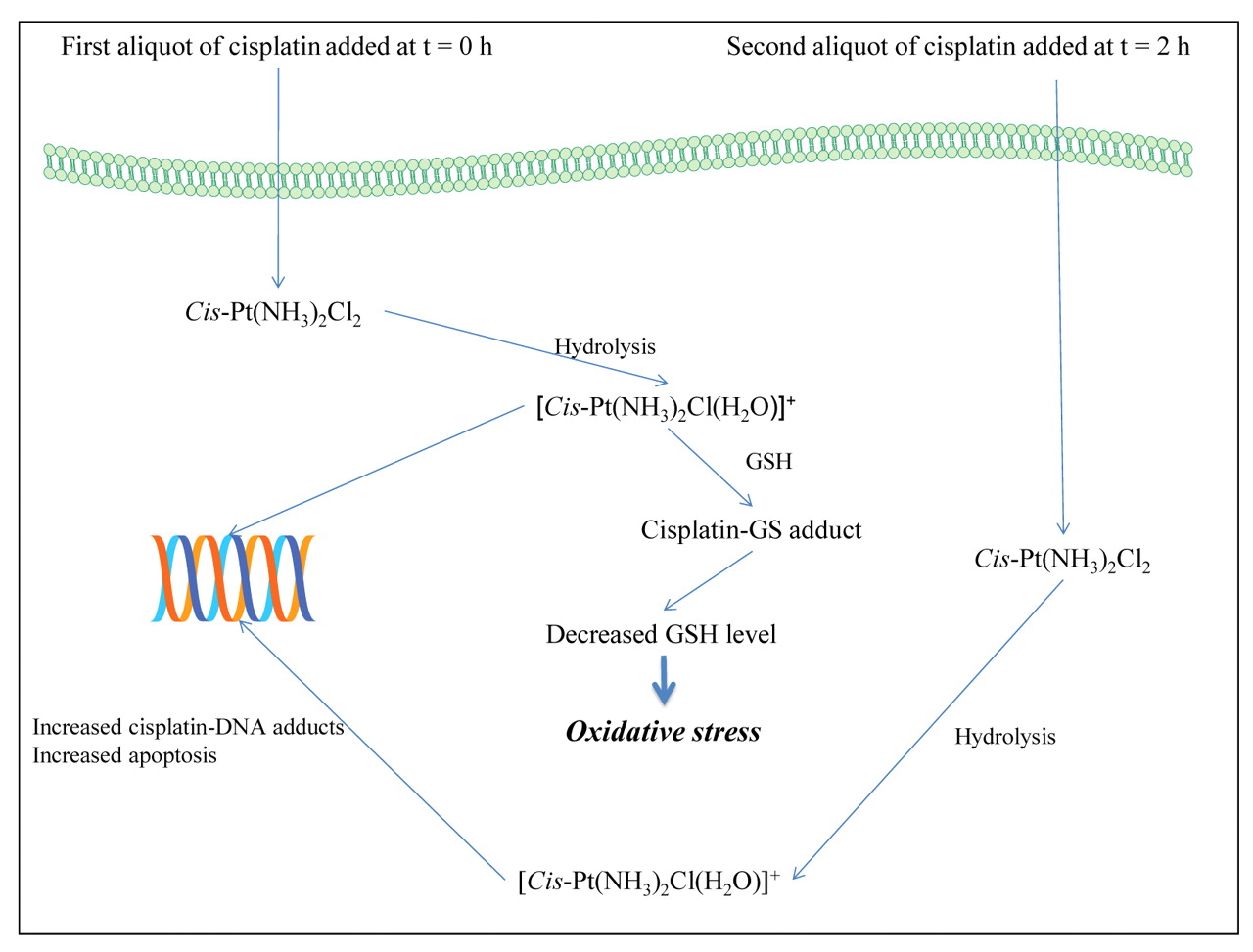
Key events in the administration of CS in two aliquots with a time gap to the human ovarian A2780, A2780cisR and A2780ZD0473R cell lines.
For the combinations of CS with Bort in A2780 cell line, all three sequences of administration were found to be antagonistic, with the 0/2 h sequence being the most antagonistic, while the 2/0 h sequence was the least (and was actually close to being additive). For the A2780cisR and A2780ZD0473R cell lines, while the 0/0 sequence of administration of CS and Bort was found to be antagonistic, 0/2 h and 2/0 h combinations were found to be synergistic, with the 2/0 h sequence being most synergistic. The results suggest that prior incubation of the ovarian cells with Bort may serve to protect them from CS-induced degradation of copper transporter CTR1 and the effect of this protection may be more significant for the resistant A2780cisR and A2780ZD0473R cell lines because of a depressed level of CTR1 expression (20). If this is so, this may lead to increased cellular accumulation of CS, which in turn may result into increased platinum–DNA binding and hence increased apoptosis. The observed higher intracellular platinum accumulation, higher platinum–DNA binding level and greater synergism associated with 2/0 h (and 4/0 h) combinations of CS with Bort (especially in the resistant A2780cisR and A2780ZD0473R cell lines) can be seen to support this idea. Antagonism observed for all combinations of YH12 and Bort for all the three human ovarian cell lines indicates that CTR1 may not be a transporter for YH12. This may be due to the much larger size of YH12 as compared to CS, in line with the observations made by Howell and co-workers (6). It was found that while CS and carboplatin accumulated with high efficiency in mouse CTR1+/+ cells (as compared to that in CTR−/− cells), the accumulation of oxaliplatin, which has a much larger size, did not depend on CTR1 for uptake (3).
For cell kill due to CS alone, it was found that greater cell kill was produced when the drug was administered in two aliquots with a time gap rather than as a bolus and that the 4 h time gap was better than 2 h. The higher cell kill was also found to be associated with higher levels of intracellular platinum accumulation and platinum–DNA binding. It is hypothesized that the administration of the first aliquot of CS places cells under oxidative stress created by depletion of cellular thiols due to their binding with CS. It may be noted that ROS can damage biomolecules and simultaneously act on a multitude of signaling pathways including those involved in cell cycle regulation, drug/ROS detoxification, and apoptosis. When the second aliquot of CS is added, more of it may bind with DNA, thus resulting in increased apoptosis. Since apoptosis due to CS follows multiple pathways, it is suggested that different pathways become more significant at different time points, so that the sequenced administration of CS in two aliquots may be looked upon as being the combination of two drugs with distinctly different mechanisms of action. This idea is illustrated in Figure 4.
Conclusion
Addition of Bort 2 h before CS is found to produce greater cell kill than the converse and the bolus, especially for the resistant A2780cisR and A2780ZD0473R cell lines, in line with the greater platinum accumulation and platinum–DNA binding level. Thus, the prevention of CTR1 degradation by Bort may play a significant role, especially in the resistant cell lines. Administration of CS in two aliquots with a time gap maximises cell kill due to CS alone. If these findings are confirmed in vivo, the results may be significant clinically.
Acknowledgements
This work was supported by Biomedical Science Research Initiative Grant and Biomedical Science Cancer Research Donation Fund.
Footnotes
-
Conflict of Interest
Zaynab Al-Eisawi, Philip Beale, Charles Chan, Jun Qing Yu and Fazlul Huq declare that they have no financial interest and personal relationships with other people or organizations that could inappropriately influence (bias) their work.
- Received April 11, 2011.
- Revision received June 10, 2011.
- Accepted June 13, 2011.
- Copyright© 2011 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved





{kind=link}
{kind=link}
{kind=link}
{kind=link}