


Abstract
Background: The hepatocyte growth factor (HGF)-Met pathway plays a role in progression to malignant characteristics in tumors, including that of resistance to anticancer drugs. The purpose of this study was to elucidate the possibility of the combination therapy of NK4, a competitive antagonist for HGF. Materials and Methods: We established a genetically modified murine colon cancer cell, CT26, to produce abundant NK4 (CT26/NK4). Cell proliferation, apoptosis, cell cycle, intracellular signaling, and 5-fluorouracil (5-FU) metabolism of this cell line were examined. Results: There was no difference in thymidylate synthase mRNA level between mock-transfected control CT26 cells and CT26/NK4 cells, suggesting that NK4 expression does not change 5-FU metabolism. NK4 gene expression enhanced 5-FU-induced caspase-3 and -7 activation of CT26. Cell cycle analysis showed that NK4 gene expression and 5-FU treatment caused an increase in the proportion of sub-G1 cells. On 5-FU treatment, phosphorylation of Akt and Erk1/2 was suppressed in CT26/NK4 less than in mock-transfected cells. 5-FU showed a stronger cytotoxic activity towards CT26/NK4 cells than control CT26 cells. Conclusion: 5-FU exerts an additional effect on apoptosis of NK4-expressing CT26 cells by down-regulating intracellular signaling of the HGF/c-Met pathway.
Hepatocyte growth factor (HGF) was originally identified as a major mediator of liver regeneration, and has been shown to affect various biological activities, such as cell proliferation, motility, morphogenesis, and apoptosis, in a wide variety of cells. In tumor tissues, however, tumor cells utilize the biological activities of HGF for tumor growth, invasion, and metastasis (1-4).
The prognosis of colon cancer has been greatly improved by the development of molecular target therapies, such as vascular endothelial growth factor (VEGF) and epidermal growth factor (EGF) receptor inhibitors, but some advanced or metastatic colon tumors remain incurable. The plasma HGF level rises in patients with colorectal cancer and can be a prognostic indicator in such patients (5, 6). In addition, c-Met is widely expressed in epithelial cells and in solid tumors, including colon cancer (7-9). Therefore, inhibition of the HGF/c-Met pathway may be an important approach to control the malignant behavior of colon tumors.
NK4, a product of proteolytic digestion of HGF, was isolated as a competitive antagonist of HGF (10). NK4 can bind to its receptor, c-Met, and abrogate HGF-induced tyrosine phosphorylation of c-Met. NK4 does not inhibit the binding of other growth factors, such as fibroblast growth factor (FGF), VEGF, and EGF, to their receptors, and hence NK4 is an HGF-specific antagonist. NK4 also exerts an antiangiogenic effect independent of its activity as an HGF antagonist (11). Thus, NK4 is expected to serve as a novel antineoplastic modality via an HGF antagonistic effect and an antiangiogenic mechanism.
We have previously reported potent antitumor effects of NK4 by stable gene expression, and adenovirus vector-mediated gene expression using CT26 murine colon adenocarcinoma cells (14). Emerging preclinical data suggest that inhibitors of the HGF/c-Met signaling pathway may also be effective in combination with chemotherapy (12, 13). Here, we investigated the effect of the combination of NK4 gene therapy and 5-fluorouracil (5-FU) chemotherapy on CT26 cells.
Materials and Methods
Cell cultures. CT26 is an undifferentiated colon adenocarcinoma cell line originally derived by intrarectal injections of N-nitroso-N-methylurethamine in a female BALB/c mouse. CT26 cells express c-Met receptors on the cell surface and respond to HGF (14). CT26 cells were genetically modified to produce human NK4 (CT26/NK4), as described previously (14). Briefly, CT26 cells were transfected with pcDNA3/NK4 by electroporation. Cells transfected with the neomycin-resistance gene (pSVneo) alone were used as a control (CT26/NEO). The CT26 transfectants were maintained in RPMI-1640 (Nacalai Tesque, Kyoto, Japan) supplemented with 100 IU/ml penicillin, 100 mg/ml streptomycin (Sigma, Welwyn Garden City, UK), and 10% heat-inactivated fetal bovine serum (FBS; JRH Bioscience, Lenexa, KS, USA).
Agents. 5-FU, cisplatin, and docetaxel were kindly provided by Kyowahakkokirin (Tokyo, Japan), Nippon Kayaku (Tokyo, Japan), and Sanofi-Aventis (Paris, France), respectively.
In vitro cell growth assay. CT26/NEO and CT26/NK4 cells were cultured in 96-well plates at a density of 5×103 cells per well overnight, then treated with 5-FU (0, 0.1, 1, 10 μM), cisplatin (0, 0.1, 1, 10 μM), or docetaxel (0, 0.001, 0.01, 0.1 μM). After treatment for 72 h, viable cells were measured by 2-(2-methoxy-4-nitrophenyl)-3-(4-nitrophenyl)-5-(2,4-disulfophenyl)-2H-tetrazolium, monosodium salt (WST-8) assay (Nacalai Tesque). Each result was corrected by values from a control study. Experiments were performed in triplicate and repeated at least three times.
Caspase-3/-7 activity assay. CT26/NEO and CT26/NK4 cells were cultured in a black 96-well plate at a density of 5×103 cells per well overnight, then treated with 0, 0.1, 1, 10 μM 5-FU. After treatment with 5-FU for 24 h, caspase-3 and -7 activity was assayed using APO-One Homogeneous Caspase-3/7 Assay kit (Promega, Madison, WI, USA). The assay was repeated three times and similar results were obtained.
Flow cytometry and measurement of apoptosis. Flow cytometric cell cycle analysis was performed by staining of permeabilized cells with propidium iodide (PI) for DNA content. CT26/NK4 and CT26/NEO were seeded in a 6-well plate at a density of 1×105 cells per well and incubated overnight. Cells were treated with 0, 0.1, 1, 10 μM 5-FU for 24 h, trypsinized, and harvested by centrifugation. Cells were resuspended in PBS (Nacalai Tesque), fixed with 0.2% Triton-X (Nacalai Tesque), and stained with PI (Sigma) solution (0.05 mg/ml PI, 1 mg/ml RNase (Qiagen, Valencia, CA, USA), 0.1% TritonX-100 in PBS). DNA content was then analyzed using a flow cytometer (FACScan, Becton-Dickinson, San Diego, CA, USA) equipped with a 488 nm argon-ion laser at 15 mW with a 530/30 nm bandpass filter. Mod Fit LT ver. 1.00 (Verity Software House Inc., Topsham, ME, USA) was used for cell cycle analysis.
Apoptotic cells were measured by flow cytometry, using MEBSTAIN Apoptosis Kit® (Medical & Biological Laboratories, Nagoya, Japan), which is based on the TUNEL method (16, 17). The kit was used according to the manufacturer's instructions. 1×105 cells were sampled after treatment with 5-FU for 24 h. The number of TUNEL-positive cells per 1×104 cells was determined with a flow cytometer.
Western blotting. CT26 transfectants were cultured with 0, 0.1, 1, 10 μM 5-FU for 10 min in the presence of 10 ng/ml HGF, which was purified from the conditioned medium of Chinese hamster ovary cells transfected with human HGF cDNA. The cells were washed twice with PBS and lysed in Pro-Prep (iNtRON Biotechnology, Gyeonggi-do, Korea). Protein extracts were run on 7% polyacrylamide gel and were transferred onto polyvinylidene difluoride (PVDF) membranes (Millipore, Billerica, MA, USA). Nonspecific binding was blocked by incubation of the membranes with 5% nonfat milk at 4°C overnight, followed by incubation with the primary antibody at room temperature overnight. The primary antibodies were used at 0.5 μg/ml for detection of phospho-HGF R/c-Met (Affinity-purified rabbit anti-phospho-HGF R/c-Met antibody (Y1234/Y1235); R&D Systems, Minneapolis, MN, USA) and anti-β-actin (Abcam, San Francisco, CA, USA). Anti-rabbit IgG antibody (Cell Signaling, Danvers, MA, USA) conjugated with horseradish peroxidase was used as the secondary antibody for detection of phospho-HGF R/c-Met and β-actin. Both of the secondary antibodies were applied at 1:2000 dilution for 1 h at room temperature, and the signal was detected with a chemiluminescence kit (GE Healthcare, Amersham, Buckinghamshire UK).
ELISA. To examine the phosphorylations of Akt at serine-473 and Erk1/2 at threonine-185/tyrosine-187, cell lysates were prepared in the same method as described for Western blotting and analyzed with each phospho/total protein enzyme-linked immunosorbent assay (ELISA) kit (BioSource, Camarillo, CA, USA). Changes in the quantity of phospho-proteins were normalized by each total protein level.
Quantitative real-time RT-PCR (qRT-PCR). Cells were incubated with 0, 0.1, 1, 10 μM 5-FU for 12 h. Total RNA was then extracted from CT26 transfectants using the ISOGEN RNA extraction kit (Nippon Gene, Tokyo, Japan) according to the manufacturer's instructions. qRT-PCR was performed with the QuantiTect SYBR Green PCR kit (Roche, Basel, Switzerland) for the thymidylate synthase (Tyms) and glyceraldehyde-3-phosphate dehydrogenase (Gapdh) genes. Detection of the emission intensity in real-time of SYBR Green bound to double-stranded DNA was carried out using the Roche PCR Thermal Cycler MP. The PCR conditions for Tyms and Gapdh amplification consisted of an initial 15 s degradation at 95°C, followed by 40 cycles of 94°C for 15 s, 60°C for 30 s, and 72°C for 30 s. This was followed by a final 15 s extension at 65°C. Primer sequences for Tyms were 5’-GGA GTA GAC CAG CTG CAA AAA GTG-3’ and 5’-GAT ATG TGC AAT CAT GTA GGT GAG-3’. Gapdh primer sequences were 5’-CAT GGC CTT CCG TGT TCC TA-3’ and 5’-GCG GCA CGT CAG ATC CA-3’.
Statistical analysis. Two-tailed Student's t-test was used to statistically evaluate the difference between two values. The level of statistical significance was set at p<0.05.
Results
NK4 gene expression enhances 5-FU-induced growth inhibition of CT26 cells. To select the best partner for NK4 gene therapy from among conventional chemotherapeutic agents, we examined the effect of 5-FU, cisplatin, and docetaxel on in vitro proliferation of NK4-expressing CT26 cells by WST-8 assay. There was little difference in cell proliferation between CT26/NK4 and CT26/NEO under conditions without drugs. All drugs reduced cell proliferation of both transfectants in a dose-dependent manner. 5-FU, in particular, dose-dependently reduced the cell proliferation of CT26/NK4 cells to a greater extent compared with that of CT26/NEO cells than did cisplatin and docetaxel (Figure 1a-c). From the results of the cell proliferation assay, we selected 5-FU as the best chemotherapeutic agent for combination with NK4 gene therapy and examined the assays using 5-FU thereafter.

Effect of NK4 gene expression on CT26 cell proliferation in vitro. CT26/NK4 and CT26/NEO cells were treated with 5-fluorouracil (5-FU), cisplatin, and docetaxel. After 72 h treatment, viable cells were measured by WST-8 assay. a: 5-FU; b: cisplatin; c: docetaxel. Each result was corrected by a control study. Each error bar denotes the standard deviation (*p<0.05).

Effect of NK4 gene expression on apoptosis of CT26 cells in vitro. CT26/NK4 and CT26/NEO were treated with 5-FU. After 5-FU treatment for 24 h, caspase-3 and-7 activities were assayed by APO-One Homogeneous Caspase-3/-7 Assay kit. Each result was corrected by the control study. Representative data are shown.
NK4 gene expression enhances 5-FU-induced apoptosis of CT26 cells. We then examined whether apoptosis is involved in the cytotoxicity induced by the combination of NK4 gene expression and 5-FU. We measured the activity of caspase-3 and-7, the critical mediators of apoptosis, using Z-DEVD-R110 as a fluorogenic substrate for caspase-3 and -7. 5-FU dose-dependently activated caspase-3 and -7 of CT26 transfectants after treatment for 24 h, and at 10 μM 5-FU-induced caspase-3 and-7 activation of CT26/NK4 cells was higher than that of CT26/NEO cells (Figure 2).
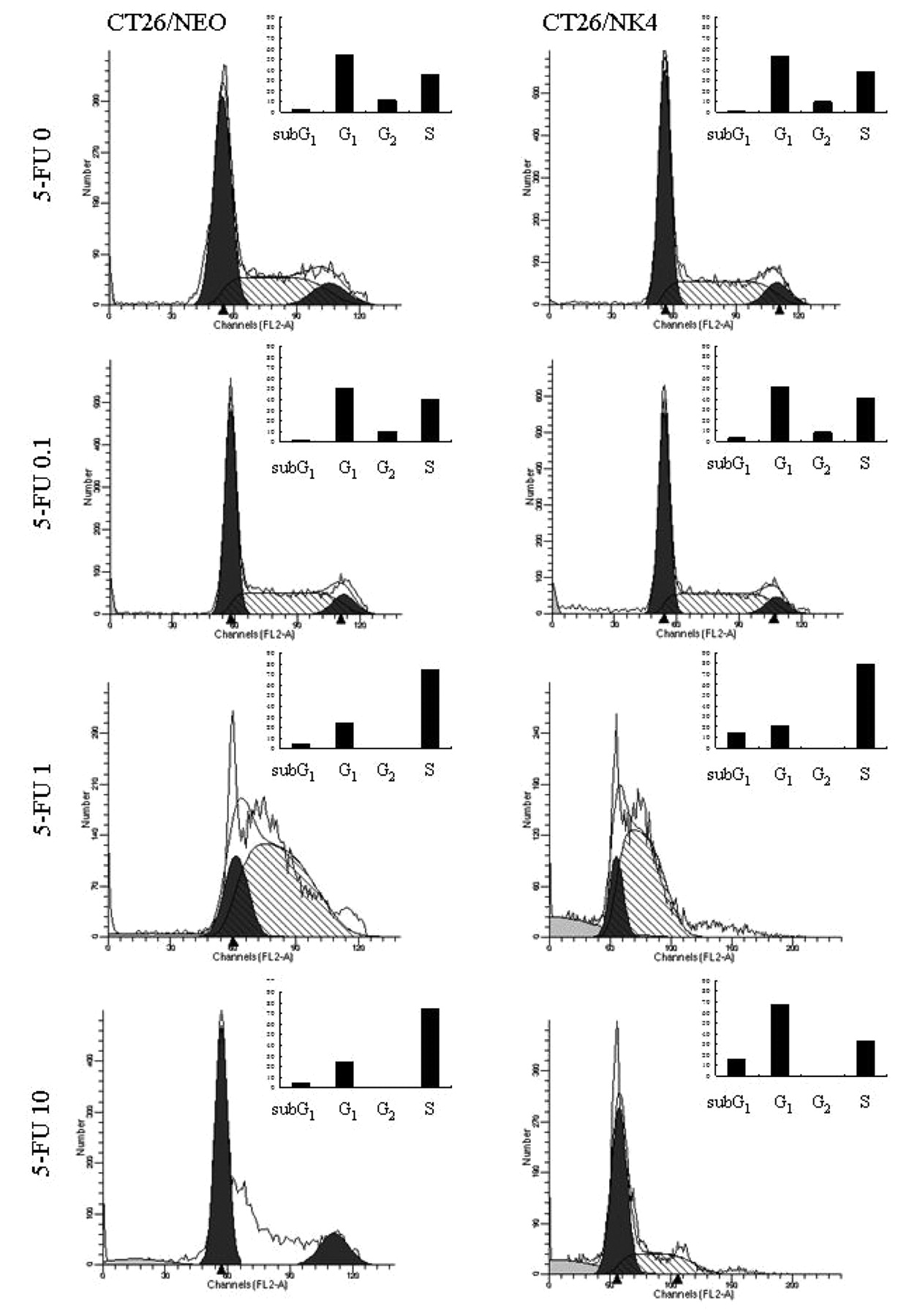
5-FU acts as a cytostatic agent by arresting cells in the G2 phase (18, 19); therefore, we performed flow cytometric analysis after treatment with 5-FU for 24 h to confirm the G2 arrest with these transfectants. NK4 gene expression and 5-FU treatment caused relative depletion of cells in the G1 phase of the cell cycle, accumulation in S phase, G2 disappearance in both cell transfectants, and an increase in the proportion of sub-G1 cells, except at 10 μM (Figure 3). 5-FU at 10 μM had a similar effect to that at 0 μM, suggesting that other effects, such as osmolarity, might have influenced the result. TUNEL assay showed that 5-FU at the higher concentrations increased the proportion of cells in apoptosis 24 h after treatment (Figure 4) with CT26/NK4 cells being more greatly affected than CT26/NEO cells.
5-FU influences intracellular signaling of HGF/c-Met in CT26 cells. Western blotting analysis using anti-phosphorylated c-Met antibody revealed that phosphorylation of c-Met was strongly inhibited in CT26/NK4 cells, even in the presence of HGF, while in CT26/NEO cells, c-Met was phosphorylated by HGF, as we reported previously (14). However, the phosphorylation of c-Met was not influenced by the addition of 5-FU (data not shown). To examine the effects of NK4 gene expression on intracellular signaling of the HGF/c-Met pathway, the phosphorylation status of Akt and Erk1/2 was examined by ELISA because these two signal transducers are key molecules in the growth of CT26 cells (20, 21). 5-FU dose-dependently suppressed the phosphorylations of Akt and Erk1/2 in both CT26/NK4 and CT26/NEO. The phosphorylations were inhibited lower by NK4 gene expression at any dose of 5-FU including 0 μM (Figure 5). The result shows that 5-FU has some additive action on the phosphorylations of Akt and Erk1/2 suppressed by NK4 gene expression.

Flow cytometric cell cycle analysis. CT26/NK4 and CT26/NEO cells were stained with PI for DNA content after treatment for 24 h with 5-FU. DNA content was analyzed using a flow cytometer. Bar graphs show the cell cycle phase distribution.

Flow cytometric analysis for quantification of apoptotic cells. CT26/NK4 and CT26/NEO cells were sampled 1×105 at 72 h after 5-FU treatment. The number of TUNEL-positive (apoptotic) cells per 1×104 cells was determined with a flow cytometer and is shown.
Little change in mRNA expression levels of thymidylate synthase. When considering the additional effect of NK4 gene expression in combination with 5-FU on intracellular signaling downstream of HGF/c-Met, there is a possibility that NK4 gene expression influences 5-FU metabolism. Therefore, we assessed the expression of Tyms, one of the metabolic enzymes of 5-FU. The expressions of Tyms after treatment with 5-FU in CT26 transfectants are shown in Figure 6. mRNA levels of Tyms dose-dependently decreased after treatment with 5-FU. However, there was no difference between levels in CT26/NK4 and these in CT26/NEO cells. This result suggests that NK4 gene expression has no influence on enzymes involved in 5-FU metabolism.
Discussion
In the present study, we demonstrated that the combination of NK4 gene therapy with chemotherapeutic agents, particularly 5-FU, enhanced the growth inhibition of CT26 cells. When we assessed the mechanism involved, 5-FU was found to exert an additional effect on apoptosis of CT26/NK4 by down-regulating intracellular signaling of the HGF/c-Met pathway. The influence of NK4 gene expression on enzymes involved in 5-FU metabolism was not elucidated.
In our previous study, we showed that NK4 gene expression alone exerts potent antitumor activity of CT26 by inhibition of tumorigenesis, peritoneal dissemination, and lung metastasis (14, 22, 23). When considering the clinical application of NK4, however, combination therapy with chemotherapeutic agents is required. Therefore, we examined whether the combination of NK4 gene therapy with chemotherapy is effective on CT26 cells, and if it is effective, which chemotherapeutic agent is the best partner for NK4 gene therapy. In vitro, NK4 gene expression does not seem to inhibit CT26 cell proliferation (Figure 1), probably because HGF does not affect it, even though CT26 cells express c-Met receptor (14). A similar phenomenon was confirmed in other cell lines, such as those for gastric, pancreatic, and colorectal cancer (24-26). Many researchers insist that the inhibitory effect of NK4 on tumor growth is exerted in vivo predominantly by antiangiogenesis rather than by HGF antagonism (14, 27, 28). However, we found that 5-FU inhibited in vitro cell proliferation of CT26/NK4 cells more effectively than of mock transfectants. This suggests that 5-FU and NK4 gene expressions are correlated in regard to cell death. 5-FU has been used widely for colorectal cancer therapy as FOLFOX, FOLFIRI, and other regimens. In addition, molecular target drugs, such as anti-VEGF and anti-EGFR antibody, are now in clinical application. NK4 is an HGF-specific antagonist. NK4 but not anti-HGF antibody inhibits tumor growth via the bifunctional property of acting as an HGF antagonist and an angiogenesis inhibitor. Interestingly, NK4 exerts a potent inhibitory effect not only on HGF/c-Met-dependent angiogenesis, but also on the HGF/c-Met-independent angiogenesis induced by bFGF and VEGF (11). Thus, we consider NK4 as a promising molecular target drug for colorectal cancer therapy.

Regulation of Erk1/2 and Akt activation by NK4 gene expression. To examine the phosphorylation of Akt at serine-473 and Erk1/2 at threonine-185/tyrosine-187, lysates were prepared and analyzed with each phospho-/total protein by ELISA. The quantity of phospho protein was normalized to that of each total protein. Each error bar denotes the standard deviation.
We next investigated apoptosis of CT26 cells induced by the combination of 5-FU and NK4 gene expression from various aspects. The findings of activation of caspase-3 and -7, the increase in the proportion of cells in the sub-G1 phase in the cell cycle study, and the increase in the rate of TUNEL-positive cells all convinced us that NK4 gene expression would enhance cell apoptosis induced by 5-FU. Additionally, the increase in S phase in the cell cycle study indicates that NK4 gene expression exhibits additive action on cytotoxicity induced by 5-FU because 5-FU acts as a cytostatic agent by arresting cells in the G2 phase (18, 19). Concerning this combination effect, we developed two hypotheses to reveal its mechanism. First, NK4 gene expression enhances 5-FU-induced apoptosis through the influence on 5-FU metabolism. Second, in contrast, 5-FU affects the inhibition of HGF/c-Met signaling by NK4. We have already revealed that NK4 gene expression inhibits HGF-activated phosphoinositide 3-kinase (PI3K)/Akt, Erk1/2 downstream of the HGF/c-Met pathway in CT26 transfectants (23).
To test the influence of NK4 gene expression on 5-FU metabolism, we examined the activity of Tyms, which acts as a key metabolic enzyme of 5-FU. The results of qRT-PCR revealed that the combination effects are not due to the changes of 5-FU metabolism at least. To further investigate the mechanism behind the effects of NK4 gene expression and 5-FU chemotherapy, we next examined the intracellular signaling downstream of the HGF/c-Met pathway. The difference of phosphrylations of Akt and Erk1/2 between CT26/NK4 and CT26/NEO was thought to be due to NK4 gene expression because the difference was observed even at 0 μM of 5-FU. In addition, 5-FU dose-dependently suppressed these two phosphorylations in both transfectants, indicating that 5-FU also has suppressive effect on the phosphrylations of Akt and Erk1/2. The PI3K/Akt pathway is known to be involved in the regulation of cell survival, apoptosis, and growth (20, 21). Furthermore, PI3K/Akt and/or Erk1/2 are known to be associated with chemosensitivity or chemoresistance of 5-FU in colon cancer (29-31). Thus, we concluded that the combination effect was mediated through the suppression of intracellular signaling downstream of HGF/c-Met by 5-FU without changes of 5-FU metabolism.

Change in thymidylate synthase activity by NK4 gene expression. RNA of CT26/NK4 and CT26/NEO cells was isolated after 12 h treatment with 5-FU. Then total PCR was performed for thymidylate synthase and GAPDH gene. Date ware normalized by control values at 0 μM 5-FU. Each error bar denotes the standard deviation.
We have emphasized that NK4 gene therapy alone exerts potent antitumor activity by different mechanisms in addition to HGF antagonism and antiangiogenesis in a mouse model (14, 22, 23). However, the majority of human malignancies are more complex, and the inhibition of a single target alone is likely to be therapeutically ineffective. As such, some studies recently addressed the possible efficacy of NK4 in combination with conventional chemotherapeutic agents or with other signal transduction inhibitors. Chemotherapeutic agents such as 5-FU and gemcitabine significantly improve the efficiency of adenovirus-mediated gene transfer in pancreatic cancer, resulting in the promotion of antitumor activity by adenovirus expressing NK4 (32, 33). In addition, NK4 expression suppressed gefitinib (EGFR-tyrosine kinase inhibitor) resistance induced by the interaction between gastric cancer and fibroblasts, and eventually, the combination of NK4 with gefitinib synergistically decelerated tumor progression by inhibiting proliferative, angiogenic and antiapoptotic effects (34). In the present study, we demonstrated the apparent synergistic efficacy of NK4 gene therapy in combination with 5-FU in colon cancer.
Inhibition of HGF/c-Met signaling represents a promising strategy for colorectal cancer treatment either alone or as part of a combination therapy. NK4 gene therapy is thought to be the most reasonable way to maintain a high concentration of NK4. While many problems still remain in terms of gene delivery systems, the development of appropriate gene transduction methods could enable great progress in NK4 gene therapy and offer a new therapeutic option for colorectal cancer patients.
Acknowledgements
This study was supported by a Grant-in-Aid for Young Scientists and a Grant-in-Aid for Scientific Research from the Ministry of Education, Culture, Sports, Science and Technology of Japan.
- Received February 27, 2011.
- Revision received April 29, 2011.
- Accepted May 3, 2011.
- Copyright© 2011 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.