


Abstract
Etomidate is an important tool in the arsenal of the emergency physician, and it has been used in a variety of scenarios for both intubation and procedural sedation. In the present study, we investigated the cytotoxicity of etomidate including induction of apoptosis, and levels of protein and gene expressions associated with apoptotic cell death in murine leukemia RAW264.7 cells in vitro. Cytotoxic and apoptotic responses to etomidate of RAW264.7 cells, including cell morphological changes and cell viability were examined and measured by phase-contrast microscopy and flow cytometric assay, respectively. Results indicated that etomidate increased apoptotic cell morphological changes and reduced cell viability in RAW264.7 cells. 4’,6-Diamidino-2-phenylindole (DAPI) staining also showed that etomidate induced the formation of apoptotic bodies, a characteristic of apoptosis. Results from Western blotting indicated that etomidate enhanced the levels of cytochrome c, apoptosis-inducing factor (AIF), endonuclease G (Endo G), caspase-9, caspase-3 active form and Bax proteins, but it inhibited the expression of Bcl-xl, leading to apoptosis. DNA microarray assay indicated that etomidate increased the expression of 17 genes (LOC676175; Gm14636; 2810021G02Rik; Iltifb; Olfr1167; Ttc30b; Olfr766; Gas5; Rgs1; LOC280487; V1rd4; Hist1h2bc; V1rj3; Gm10366; Olfr192; Gm10002 and Cspp1) and reduced the expression of 15 genes: (Gm10152; Gm5334; Olfr216; Lcn9; Gm10683; Gm5100; Tdgf1; Cypt2; Gm5595; 1700018F24Rik; Gm10417; Maml2; Olfr591; Trdn and Apol7c). In conclusion, etomidate induced cytotoxic and apoptotic effects the in murine leukemia RAW264.7 cells in vitro.
In 1972, etomidate (R-(2-ethyl 1-(phenylethyl)-1H-imidazole-5-carboxylate)) was first introduced into clinical practice in Europe, and it was approved for use in the United States in 1983 (1). Etomidate has been used in a variety of scenarios for both intubation and procedural sedation. Etomidate has been deemed an appropriate agent for procedural sedation in the emergency department (2). Etomidate does have side-effects. However, for example, used as a continuous infusion for sedation, etomidate significantly increased the mortality rates of ICU trauma patients (3). Other adverse effects include apnea, emesis, hemoglobin desaturation, myoclonus, bradycardia, and emergency anxiety (4-7).
Although many studies have been reported on the mechanism of action of etomidate for clinical use, there is no information on the etomidate-induced cytotoxic effects. Thus, in the present study, we investigated the effects of etomidate on murine leukemia RAW264.7 cells in vitro by measuring cytotoxic effects, gene expression and apoptosis associated proteins in vitro.
Materials and Methods
Reagents and chemicals. Etomidate was purchased from Lipuro, B. Braun Co. Ltd. (Melsungen, Germany). Etomidate was prepared in phosphate-buffered saline (PBS), and an equal volume of PBS (0.1%) was added to the control cells as vehicle. Propidium iodide (PI) and RNase A were from Sigma-Aldrich Corp. (St. Louis, MO, USA). 4, 6-Diamidino-2-phenylindole (DAPI) was purchased from Molecular Probes/Invitrogen Life Technologies (Eugene, OR, USA). RPMI-1640 medium, fetal calf serum (FCS), penicillin and streptomycin antibiotic mixture were purchased from Invitrogen Life Technologies (Carlsbad, CA, USA). The primary antibodies to Fas, cytochrome c, apoptosis-inducing factor (AIF), endonuclease G (Endo G), caspase-9, caspase-3, Bax and Bcl-2 were obtained from Santa Cruz Biotechnology Inc. (Santa Cruz, CA, USA).
RAW264.7 cell line. The RAW264.7 murine leukemia cell line was obtained from the Food Industry Research and Development Institute (Hsinchu, Taiwan, ROC). The cells were maintained in RPMI-1640 medium with 2 mM L-glutamine, supplemented with 10% heat-inactivated FCS and 1% antibiotic/antimycotic and incubated at 5% CO2 and 37°C.
Determination of cell viability and morphology. RAW264.7 cells (2×105 cells/ml) maintained in 12-well plates were treated with 0, 2.5, 5, 10, 20 and 40 μg/ml etomidate for 48 h. At the end of incubation, the cells were examined and photographed by a phase-contrast microscope at ×200 or were harvested for measuring the total percentage of viable cells as previously described (8, 9). For trypan blue dye exclusion assay, cells from each treatment were collected and re-suspended in PBS, and then were mixed with 0.4% Trypan blue stain. Cells that excluded blue dye (live cells) and cells that contained blue dye (dead cells) were counted using a hemocytometer under a light microscope (10, 11). The obtained values were compared with PBS only (control) and the quantitative analysis of cell viability was carried out.
Determination of apoptotic bodies by DAPI nuclear staining. The presence of apoptotic bodies and nuclei morphology were examined by DAPI staining. RAW264.7 cells (2×105 cells/well) in 12-well plates were exposed to 40 μg/ml etomidate for 24 h, then cells were fixed in 4% paraformaldehyde-PBS solution for 15 min and were stained with DAPI (300 nmol/l) for 30 min at room temperature. Cells were examined for apoptotic bodies and nuclear morphology and photographed under fluorescence microscopy. Apoptotic cells were recognized and determined based on characteristic observations including the presence of condensed, fragmented, and degraded nuclei (8, 9).
Determination of apoptosis-associated proteins by Western blot assay. RAW264.7 cells (5×105 cells/ml) were placed in 6-well plates with RPMI-1640 medium with 10% FCS for 24 h. Cells were treated with or without 18 μg/ml etomidate for 48 and 72 h. Cells were individually collected from each treatment and total protein was extracted into the PRO-PREP™ protein extraction solution (iNtRON Biotechnology, Seongnam, Gyeonggi-Do, Korea), and centrifuged at 12,000 rpm for 10 min at 4°C as previously described (12, 13). The total protein of each sample was determined by Bradford assay and all cell lysates were treated with reducing sample buffer, and boiled for 5 min at 100°C. Proteins were resolved on an SDS polyacrylamide gel via electrophoresis as described previously (14, 15). The gel was then transferred to PVDF membrane in Western transfer buffer. The membrane was stained by primary antibodies (anti-Fas, -cytochrome c, -AIF, -Endo G, -caspase-9, -caspase-3, -Bax and -Bcl-2) overnight, then washed and stained by appropriate horseradish peroxidase-conjugated secondary antibodies (GE Healthcare, South San Francisco, CA, USA). The intensity of immunoreactive bands was determined using a densitometer (Molecular Dynamics, Sunnyvale, CA, USA) equipped with Image QuaNTsoftware (8, 9).
Determination of apoptosis-associated gene expression by microarray assay. RAW264.7 cells (5×105 cells/ml) were maintained in 12-well plates in RPMI-1640 medium with 10% FCS for 24 h. Cells were treated with 0 and 18 μg/ml etomidate for 48 h. The total RNA was extracted by using Qiagen RNeasy Mini Kit (Qiagen, Inc, Valencia, CA, USA) (9). The isolated total RNA was used for cDNA synthesis and labeling and microarray hybridization, which was then followed by fluor-labeled cDNA hybridized complements on the chip (Affymetrix GeneChip Human Gene 1.0 ST array; Affymetrix, Santa Clara, CA, USA) (16). The resulting localized concentrations of fluorescent molecules were detected, quantitated (Asia BioInnovations Corporation, Taipei, Taiwan, ROC) and analysed by using Expression Console software (Affymetrix) with default RMA parameters. Genes regulated by etomidate were determined to be these showing a 2-fold change in expression (16-18)
Statistical analysis. Student's t-test was used to analyze differences between exposure to etomidate and the untreated (control) group. All data are presented as the means±S.D. of three experiments and a p-value less than 0.05 was considered significant.
Results
Etomidate reduced the cell viability and induced morphological changes of RAW264.7 cells. Cell morphological changes of RAW264.7 cells exposed to 18 μg/ml etomidate, included condensation both in cytoplasm and nucleus thereby resulting in granulation in the majority of RAW264.7 cells (Figure 1). Etomidate-treated cells exhibited morphological changes and also floated on the well compared to the control cells. After cells were exposed to different concentrations of etomidate for 48 h, cell viability was assessed (Figure 2). Etomidate reduced cell viability, at concentrations of 20 and 40 μg/ml etomidate, the cell viability was 34% and 32%, respectively, and the concentration required to reduce viability by 50% (EC50) was 16.34 μg/ml.
Etomidate induced apoptotic body formation (chromatin condensation) in RAW264.7 cells. Cell morphological changes and a reduction of viable cells showed that etomidate induced cytotoxic effects on RAW264.7 cells. To confirm whether or not etomidate induced apoptosis, cells were stained by DAPI after exposure to etomidate and the results are shown in Figure 3. The results indicated that untreated cells were round and had homogeneous nuclei, whereas etomidate-treated cells showed condensed nuclei and apoptotic bodies (Figure. 3).
Etomidate affected the levels of apoptosis proteins in RAW264.7 cells. In order to confirm that etomidate-induced apoptosis was achieved through the changes of associated proteins in RAW264.7 cells, cells were exposure to 18 μg/ml of etomidate for 24 and 48 h, and then the levels of apoptosis-associated proteins were analyzed by Western blotting. As shown in Figure 4, etomidate increased the levels of Fas, cytochrome c, AIF, Endo G, active caspase-9 and -3, and Bax (pro-apoptotic protein), but it reduced the level of Bcl-xl (anti-apoptotic protein).
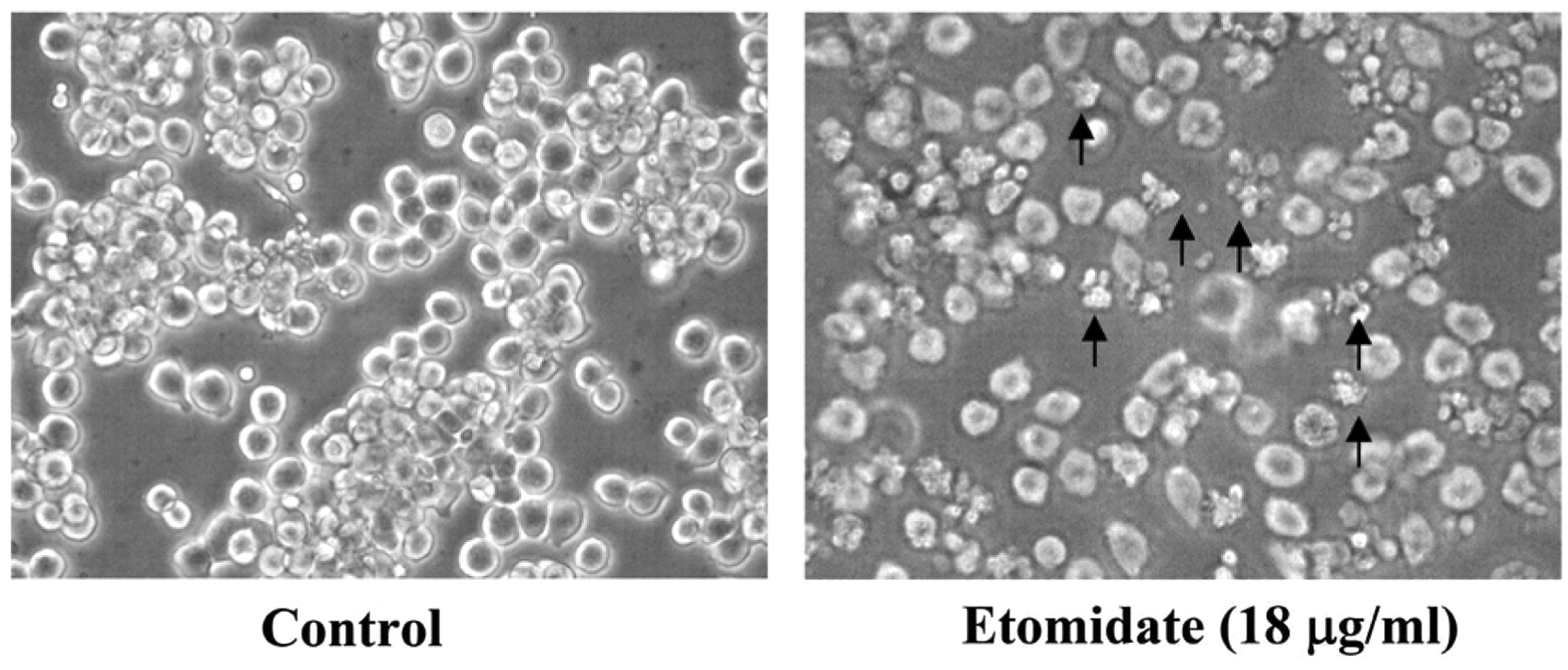
Etomidate-induced morphological changes in RAW264.7 cells. Cells were cultured with 18 μg/ml etomidate for 48 h and then examined for morphological changes and photographed by using a phase-contrast microscope as described in the Materials and Methods.
Etomidate affected gene expression in RAW264.7 cells. Cells were exposed to 18 μg/ml etomidate for 24 h, and were then harvested to isolate RNA for DNA microarray analysis. The microarray analysis showed that expression of 32 genes was altered by at least two-fold (17 genes, up-regulated; 15 genes, down-regulated) compared with the untreated control cells. We observed that LOC676175, Gm14636, 2810021G02Rik, Iltifb, Olfr1167, Ttc30b, Olfr766, Gas5, Rgs1, LOC280487, V1rd4, Hist1h2bc, V1rj3, Gm10366, Olfr192, Gm10002 and Cspp1 mRNA were up-regulated and Gm10152, Gm5334, Olfr216, Lcn9, Gm10683, Gm5100, Tdgf1, Cypt2, Gm5595, 1700018F24Rik, Gm10417, Maml2, Olfr591, Trdn and Apol7c mRNA were down-regulated in the etomidate-treated cells (Figure. 5A). The top change in gene expression as scored by the number of pathways network from GeneGo analysis program is shown in Figure 5B. Thick blue lines indicate the fragments of canonical pathways. These genes may also be involved in the apoptosis-inducing effect of etomidate on RAW264.7 cells.
Discussion
Etomidate for procedural sedation has become a common drug used among emergency physicians because of its relatively safe adverse effect profile. It has been used in all age groups with success (1). There is no report indicating that etomidate may also affect cellular processes in cancer cells. Moreover, the effects of apoptosis and associated gene expressions in murine leukemia cells after being exposed to etomidate are unclear. In the present study, it was demonstrated for the first time that etomidate can induce cytotoxic effects, apoptosis and changes in gene expression in murine leukemia RAW264.7 cells in vitro.
The results from Western blotting indicated that etomidate increased the protein levels of Fas, cytochrome c, AIF, Endo G, caspase-9 and -3 and Bax, but it reduced the level of Bcl-xl (Figure 4). This clearly shows that the ratio of Bax/Bcl-xl increased. Furthermore, etomidate increased the levels of cytochrome c, AIF and Endo G which are known to be released from mitochondria, and then activate caspase-9 and -3 causing cell apoptosis (9). This would appear to be the case with etomidate via a mitochondria-dependent pathway (9, 13, 19-21) based on the change of the ratio of Bax/Bcl-xl which led to cytochrome c, AIF and Endo G, release from mitochondria.

Etomidate reduced the percentage of viable RAW264.7 cells. Cells were incubated with different concentrations of etomidate for 48 h and then were measured for viability. The total viable cells were counted by PI-incorporation and flow cytometric analysis as described in the Materials and Methods. Each point is the mean±S.D. of three experiments. ***p<0.001.
Furthermore, this study also investigated etomidate induced cytotoxic effects associated with the gene expression in RAW264.7 cells. The DNA microarray assay was used and we found that etomidate increased LOC676175, Gm14636, 2810021G02Rik, Iltifb, Olfr1167, Ttc30b, Olfr766, Gas5, Rgs1, LOC280487, V1rd4, Hist1h2bc, V1rj3, Gm10366, Olfr192, Gm10002 and Cspp1 mRNA gene expressions and reduced Gm10152, Gm5334, Olfr216, Lcn9, Gm10683, Gm5100, Tdgf1, Cypt2, Gm5595, 1700018F24Rik, Gm10417, Maml2, Olfr591, Trdn and Apol7c mRNA gene expression (Figure 5B) which might associated with cytotoxic effects in etomidate-treated RAW246.7 cells in vitro.
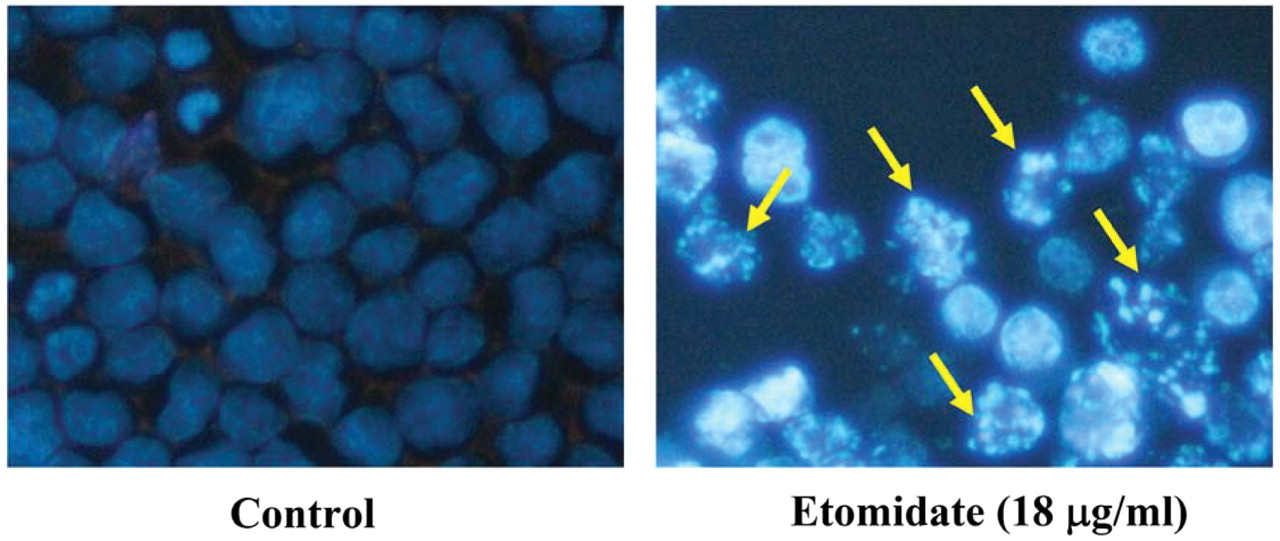
Etomidate-induced apoptotic body formation in RAW264.7 cells. Cells were treated with 18 μg/ml etomidate for 24 h and apoptotic body formation (arrows) as an indicator of apoptosis was determined by DAPI staining then photographing cells under fluorescence microscopy (×200) as described in the Materials and Methods.

Etomidate affected levels of proteins associated with apoptosis in RAW264.7 cells. Cells were treated with 18 μg/ml etomidate for 48 and 72 h then were collected and total protein determined for Western blotting as described in the Materials and Methods. The associated protein expressions (A: Fas, cytochrome c, AIF, Endo G, caspase-9 and -3; B: Bax and Bcl-xl) were estimated by Western blotting, as described in the Materials and Methods.

Etomidate-induced possible canonical pathways in RAW264.7 cells. Cells were exposed to 18 μg/ml etomidate for 24 h then were collected and total RNA extracted for DNA microarray assay. A: The top change in gene expression scored by the number of pathways from GeneGo analysis. B: Thick blue lines indicate the canonical pathways. Up-regulated genes are marked with red circles, and down-regulated ones with blue circles. The ‘checkerboard’ color indicates mixed expression for the gene between files or between multiple tags for the same gene. (ARC: activity-regulated cytoskeleton-associated protein; Arp2/3: actin-related protein 2/3 complex; CDC42: cell division cycle 42 (GTP binding protein, 25kDa); CDK1: cyclin-dependent kinase 1; CFTR: cystic fibrosis transmembrane conductance regulator; DAB1: disabled homolog 1; FAK1: ODA5-associated flagellar adenylate kinase; JAK2: Janus kinase 2; LIMK1: LIM domain kinase 1; MYOG: myogenin (myogenic factor 4); N-WASP: neuronal Wiskott-Aldrich syndrome protein; NEDD4: neural precursor cell expressed, developmentally down-regulated 4; NIK, MA3K14: serine/threonine protein kinase NIK/mitogen-activated protein 3 K 14; PAK1: p21 protein (Cdc42/Rac)-activated kinase 1; PBX1: pre-B-cell leukemia homeobox 1; PLC-beta3: phospholipase C, beta 3; PTEN: phosphatase and tensin homolog1; RAG1: recombination activating gene 1; RCC1: regulator of chromosome condensation 1; RNF7: ring finger protein 7; RGS1: regulator of G-protein signalling 1; Shc: Src homology 2 domain containing) transforming protein 1; SIX4: SIX homeobox 4).
Acknowledgements
This work was supported by a grant DOH100-TD-C-111-005 from Taiwan Department of Health, China Medical University Hospital Cancer Research Center of Excellence.
- Received February 22, 2011.
- Revision received May 4, 2011.
- Accepted May 6, 2011.
- Copyright© 2011 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}