


Abstract
Aim: To assess radiosensitzing potential of huachansu (HCS) and delineate the underlying mechanisms. Materials and Methods: Lung cancer cell lines were exposed to HCS, radiation or both and subjected to survival assays, Western blots, apoptosis assay and immunocytochemical analysis. Results: HCS suppressed the viability of all three lung lines tested and enhanced radiosensitivity of H460 and A549 (wild-type p53) only with no effect on H1299 (p53 null) cells. HCS prolonged the presence of radiation-induced γH2AX foci and increased radiation-induced apoptosis. Western blots showed that HCS increased cleaved caspase-3 and cleaved poly-(ADP-ribose) polymerase (PARP) levels, as well as reducing BCL-2 and p53 protein levels in H460 cells. Conclusion: HCS-enhanced radiosensitivity of human lung cancer lines appeared to be p53-dependent. Inhibition of DNA repair and increase in radiation-induced apoptosis may have served as underlying mechanisms. These data suggest that HCS may have potential to improve the efficacy of radiotherapy.
Radiotherapy has traditionally been the treatment of choice for locoregionally advanced, unresectable cancer, including lung cancer. Technologic advancements in radiation planning and delivery methods, the use of modified radiation fractionation schedules, and the combination of radiotherapy with chemotherapeutic drugs have significantly improved local tumor control and patient survival over the past two decades (1, 2). Chemoradiation is the most fully developed treatment approach, particularly concurrent chemoradiotherapy, in which chemotherapeutic drugs in addition to their own antitumor actions render tumor cell clonogens more susceptible to being killed by ionizing radiation. Although recent clinical trials have shown that concurrent chemoradiotherapy is superior to radiotherapy alone for controlling locoregional disease and for improving survival among patients with lung cancer, the overall effectiveness of therapy is nevertheless rather limited, with 5-year survival rates of only about 15% to 20% (3-5). The therapeutic improvements have been achieved by using standard chemotherapeutic agents such as cisplatinum and taxanes, which have been selected for combined treatment based primarily on their own antitumor activity as single agents. Unfortunately, concurrent chemoradiotherapy using these agents is commonly associated with considerable toxicity to normal tissues, which limits the radiation dose that can be safely delivered. Thus, new strategies are needed to improve the therapeutic ratio of combined chemoradiotherapy for this purpose.
Recent research has focused on targeting diverse molecular signaling networks and processes in cancer cells that are responsible for radioresistance or chemoresistance. Some of these approaches, such as inhibition of the epidermal growth factor receptor signaling pathway (6), have already shown potential therapeutic efficacy in clinical trials when combined with radiotherapy. Another approach is to search for antitumor drugs that increase the radiosensitivity of tumor cells, or which are less toxic to normal tissues than are the standard chemotherapeutic agents that commonly used in combination with radiotherapy. Huachansu (HCS) may be such an agent.
Chansu, the dried secretion from the skin glands of Bufo bufo gargarizans Cantor or B. melanostictus Schneider, has long been used for cancer treatment in China and other Asian countries. HCS, an injectable form of chansu, is a sterilized hot-water extract of dried toad skin that has been widely used as an antitumor agent in traditional Chinese medicine for various types of cancer, especially liver and pancreatic cancer (7-12). A phase I clinical study involving patients with advanced hepatocellular carcinoma, non-small cell lung cancer, or pancreatic cancer, jointly conducted by Fudan University Cancer Hospital, in Shanghai and The University of Texas MD Anderson Cancer Center in Houston, showed that HCS was well tolerated and had encouraging antitumor efficacy (13). The extract contains several biologically active substances, primarily indole alkaloids (bufotenine, bufotenidine, and cinobufotenine) and steroidal cardiac glycosides (bufalin, resibufogenin, cinobufagin, cinobufotalin, marinobufagin, and bufotalin) (14). Recent studies showed that the antitumor activity of HCS can be attributed mainly to the cardiac glycosides it contains, including bufalin, resibufogenin, and cinobufagin (15-18).
Among the cardiac glycosides derived from chansu, the bufadienolides, including bufalin, cinobufagin, and epoxybufanolides, have been found to inhibit tumor cell proliferation and induce apoptosis in several types of human cancer cell lines, including leukemia HL-60 and U937 cells, prostate cancer PC3 and DU145 cells, and human epidermoid carcinoma KB cells (17, 19-21). These glycosides induce differentiation and apoptosis in several human leukemia cell lines through alteration of expression of c-MYC and BCL-2. They also increase the activity of caspase-3 in DU145 and PC3 cells, and caspase-9 in LNCaP prostate cancer cells (17). In addition, bufalin was found to inhibit proliferation of human leukemia U927 cells by activating mitogen-activated protein kinase (MAPK) via a signaling pathway that included RAS, RAF-1, and MAPK-1 (22). Other reports suggest that bufalin induces changes in cell cycle distribution by reducing levels of topoisomerase I and II (20, 21, 23, 24) and by down-regulating cyclin A, BCL-2, and BCL-xL and increasing the expression of p21 and BAX (18, 24, 25).
Cardiac glycosides such as digoxin, oleandrin, and bufalin have also shown significant antitumor effects in preclinical in vivo tumor models (26). For example, bufalin was highly effective in reducing tumor size and prolonging the lifespan of mice bearing orthotopic human hepatocellular carcinoma (BEL-7402) xenografts (18). The treatment induced massive apoptosis in tumors but importantly was not associated with adverse morphologic changes in myocardial, hepatic, or renal tissues, suggesting a selective antitumor response. Similarly, several other studies showed that cardiac glycosides such as oleandrin and bufalin may selectively induce apoptosis in cancer cells (27-30).
In addition to being selectively cytotoxic for tumor cells, cardiac glycosides such as ouabain, bufalin and oleandrin have been shown to enhance the response of cancer cells to the cytotoxic actions of ionizing radiation (31-34). Ouabain was reported to enhance the in vitro radiosensitivity of several tumor cell types, including lung squamous cell carcinoma (31, 32), colon adenocarcinoma (33, 35), and melanoma (35). Lawrence (33) reported that ouabain-induced radiosensitization was selective for cancer cells; specifically, A549 human lung adenocarcinoma cells were radiosensitized but human lung fibroblasts were not. The mechanisms of radiosensitization have not been fully elucidated, but inhibition of repair of sublethal radiation damage and increased radiation-induced apoptosis have been reported as possibilities. Bufalin reportedly enhanced the radiosensitivity of Chinese hamster ovary cells by inhibiting DNA repair (36). A previous report from our group (34) showed that oleandrin enhanced the radiosensitivity of PC-3 human prostate carcinoma cells, in part by sensitizing them to radiation-induced apoptosis. The present study was undertaken to investigate whether HCS enhanced radiosensitization of human lung tumor cells grown in vitro and to define the underlying mechanisms associated with its radiosensitization.
Materials and Methods
Cell cultures. The human lung cancer cell lines H460 (p53 wild-type [wt]), A549 (p53 wt), and H1299 (p53 null) were obtained from the American Type Culture Collection (Manassas, VA, USA) and maintained in RPMI-1640 medium supplemented with 10% fetal calf serum, 10,000 U/ml of penicillin–streptomycin, and 2 mM L-glutamine. Cells were grown as monolayers in 75-cm2 flasks and maintained in a humidified 5% CO2/95% air atmosphere at 37°C.
Cell viability assay for HCS cytotoxicity. H460, A549, and H1299 cells were plated in 96-well plates and treated with different concentrations (0.05-50 mg/ml) of HCS (Anhui JinChan Biochemical Sharing Inc., Anhui, P.R. China) for various periods (24, 48, or 72 h). Cells were then stained with 3-[4,5-dimethylthiozol-2-yl]-2,5-diphenyltetrazolium bromide (MTT, 200 μg/ml) and incubated for 4 h at 37°C, after which the cells were lysed in 150 μl of ethanol/dimethyl sulfoxide mixture (1:1), and the absorbance was read at 540 nm using a 96-well plate reader.
Clonogenic cell survival assay. Cells in culture were exposed to HCS (20 mg/ml) for 24 h and then irradiated with 2 Gy, 4 Gy, or 6 Gy of γ-rays from a 137Cs source (3.7 Gy/min). Cells were then assayed for colony-forming ability by replating them in specified numbers into 100-mm dishes containing drug-free medium. After 12-14 days of incubation, cells were stained with 0.25% crystal violet in absolute ethanol, and colonies with more than 50 cells were counted. Radiation survival curves were plotted after normalizing for cytotoxicity induced by HCS alone. Clonogenic survival curves were constructed from at least three independent experiments. The average survival levels were fitted by least-squares regression using a linear quadratic model (37).
Quantification of γH2AX-foci formation. Cells were plated on coverslips (Becton Dickinson, Franklin Lakes, NJ, USA) placed in a 35 mm dish. The next day cells were irradiated with or without HCS pre-treatment (20 mg/ml) for 24 h. At various time points after irradiation, the cells were fixed for immunofluorescence analysis. After blocking with bovine serum albumin, cells were incubated with γH2AX antibody (Trevigen, Inc., Gaithersburg, MD, USA) (1:400) overnight at 4°C. Positive foci were visualized by incubation with a 1:300 dilution of fluorescein isothiocyanate (FITC)-conjugated donkey anti-rabbit IgG (Jackson ImmunoResearch, West Grove, PA, USA) for 30 minutes. Coverslips were mounted in Vectashield/4,6 diamidino-2-phenylindole (Vector Laboratories, Peterborough, UK). For each data point, 300-500 nuclei were evaluated and averaged to yield number of foci/cell.
Flow cytometry analysis for cell cycle distribution and apoptosis. The terminal deoxynucleotidyltransferase (TdT) dUTP nick end-labeling (TUNEL) assay (Apo-BrdU kit from BD Biosciences, Franklin Lakes, NJ, USA) was performed according to the manufacturer's instructions. Briefly, cells (2×106) were fixed in 1% paraformaldehyde, washed in phosphate-buffered saline, suspended in 70% ethanol, and stored at −20°C until use. Re-suspended cells were stained in a solution containing TdT and FITC-dUTP and incubated overnight at room temperature in dark. They were then rinsed and re-suspended in 0.5 ml propidium iodide/RNase A solution and analyzed by flow cytometry.
Western blot analyses. After experimental treatment, cells were lysed in a buffer containing 50 mM Tris–HCl (pH 8), 450 mM NaCl, 1% Igepal, 5 mM ethylenediaminetetraacetic acid, 1% (v/v) of protease inhibitor cocktail, and 1% (v/v) phosphatase inhibitor cocktails I and II (Sigma, St. Louis, MO, USA). Proteins (80 μg per lane) were separated by SDS-PAGE and transferred to a polyvinylidene difluoride membrane (Bio-Rad Laboratories, Hercules, CA, USA). The membrane was blocked by 5% non-fat dry milk in Tris-buffered saline and 0.1% Tween-20 (TBS-T) before incubation with the designated primary antibodies (cleaved caspase-3 and p53, Cell Signaling Technology, Inc. Danvers, MA, USA; poly (ADP-ribose) polymerase (PARP) and BCL-2, Santa Cruz Biotechnology, Inc. Santa Cruz, CA, USA). After the membrane was washed with TBS-T, it was incubated with secondary antibody (GE Healthcare, Fairfield, CT, USA) and the immune reaction was visualized using an ECL plus kit (Amersham Corp., Arlington Heights, IL, USA). The intensity ratios of bands compared to control bands were quantified using ImageQuant 5.2 software (GE Healthcare, Fairfield, CT, USA).
Statistical analyses. Student's t-tests were used to determine statistical differences between the various experimental groups; p<0.05 was considered to be significant.
Results
HCS enhanced radiosensitivity of lung cancer cells. As a first step, we determined if HCS itself affected cell survival. H460, A549, and H1299 tumor cells were incubated in the presence of 0.05-50 mg/ml of HCS for 24, 48, or 72 h, after which cell survival was determined with an MTT assay. HCS caused dose-dependent reductions in cell survival of all three cell lines tested (Figure 1). HCS at 2.5 mg/ml or less did not affect survival of any cells tested. At 5 mg/ml, HCS reduced survival of H460 cells by 33.9%±9.3% (p=0.017), A549 cells by 34.1%±2.84% (p<0.001), and H1299 cells by 16.8%±3.41% (p=0.023); at 50 mg/ml, it reduced survival of H460 cells by 88.3%±0.70% (p<0.001), A549 cells by 85.5%±0.59% (p<0.001), and H1299 cells by 70.5%±0.66% (p<0.001). Extending the incubation time beyond 24 h resulted in no further reductions in cell survival (data not shown). The IC50, defined as the concentration at which 50% of the cells were killed, was 20 mg/ml, and this dose was used in subsequent experiments that combined HCS and ionizing radiation.

Effect of HCS on cell viability. H460 (triangles), A549 (circles), and H1299 (squares) human lung cancer cells were exposed to HCS at the indicated concentrations for 24 h. Cells were then stained with MTT, lysed, and the absorbency was read at 540 nm. Viable cells were calculated as percentages of control cells. Data shown are means±SE from three independent experiments.
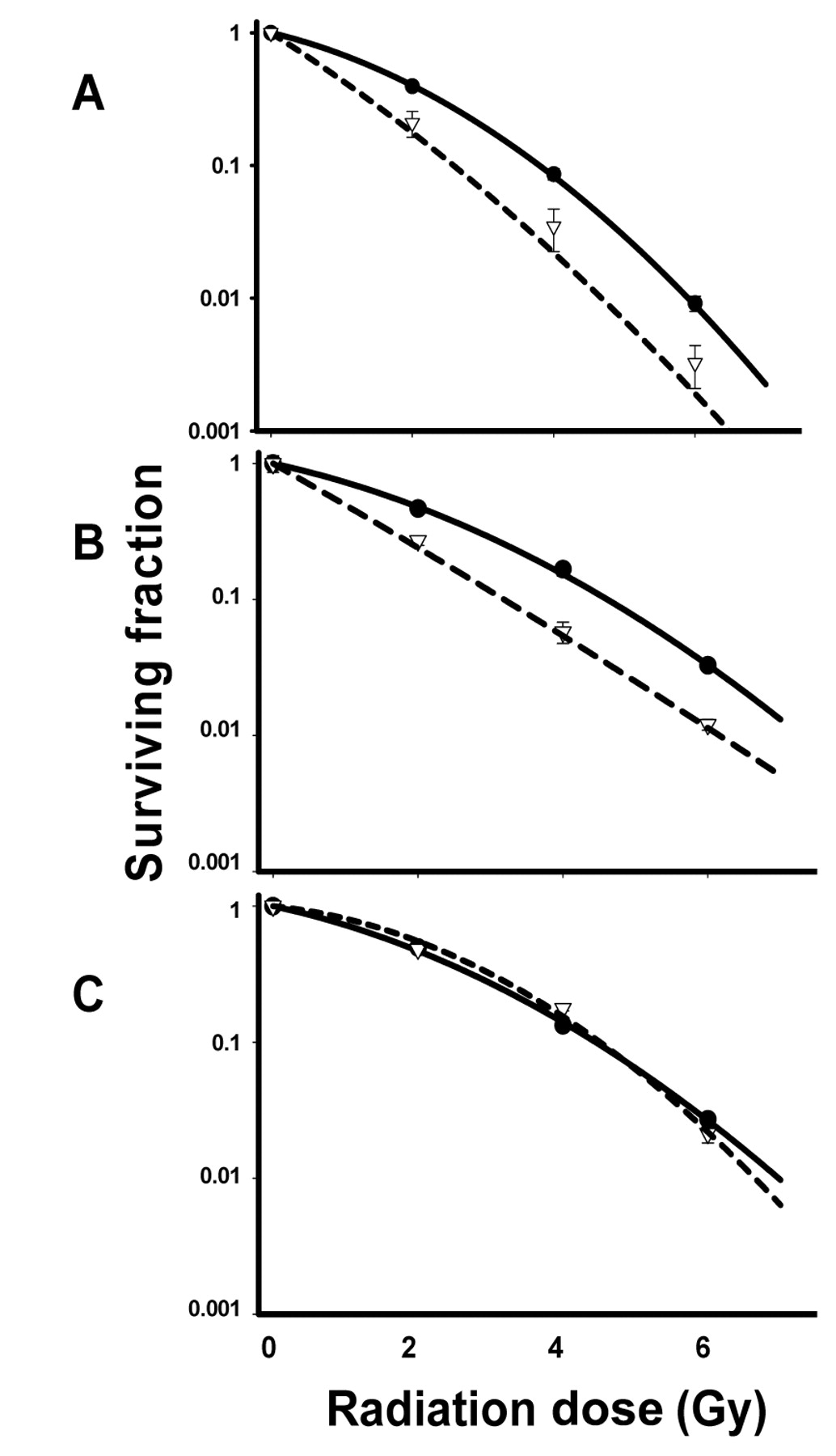
In order to determine if HCS enhanced cell radiosensitivity, we exposed H460, A549 and H1299 cells growing in monolayer cultures to 20 mg/ml HCS for 24 h and then irradiated the cells with 2 Gy, 4 Gy, or 6 Gy of γ-rays. The cells were then plated and incubated for 12 to 14 days and assessed for colony formation. Radiation dose-response curves for HCS plus radiation, normalized to account for the cytotoxicity of HCS alone, are shown in Figure 2. HCS reduced the colony-forming efficiency of all three cell lines: H460 cells from 97.5%±0.03% to 66.0%±0.04% (p<0.001), A549 cells from 60.0%±0.08% to 28.0%±0.14% (p<0.001), and H1299 cells from 46.7%±0.04% to 36.7%±0.01% (p=0.008). When combined with radiation, HCS strongly increased the radiosensitivity of H460 and A549 cells (both are p53 wt) (Figure 2 A and B), but not that of H1299 cells (p53 null) (Figure 2 C). At cell survival fraction of 0.1, the enhancement factors were 1.50 for H460 cells 1.44 for A549 cells. To determine if the drug-radiation sequence is important for increasing cell radioresponsiveness, additional experiments were performed in which the cells were first irradiated with 2 Gy, 4 Gy, or 6 Gy and then treated with 20 mg/ml HCS either immediately thereafter or 24 h later and incubated for 24 h. This treatment did not enhance cell radioresponse (data not shown). Thus, enhancement of radiosensitivity required that cells be treated with HCS before radiation delivery.
Mechanisms underlying enhanced cell radiosensitivity. We investigated several mechanisms that could be responsible for the enhanced radiosensitivity of cells treated with HCS. These included the ability of the drug to change the cell cycle distribution at the time of radiation delivery, to render DNA more vulnerable to radiation-induced DNA breaks, to slow DNA repair, or to sensitize cells to radiation-induced apoptosis. Flow cytometric analysis after 24 h of incubation with 20 mg/ml revealed that HCS did not affect the cell cycle distribution (data not shown), thus excluding cell cycle redistribution as a mechanism for the observed radiosensitization.
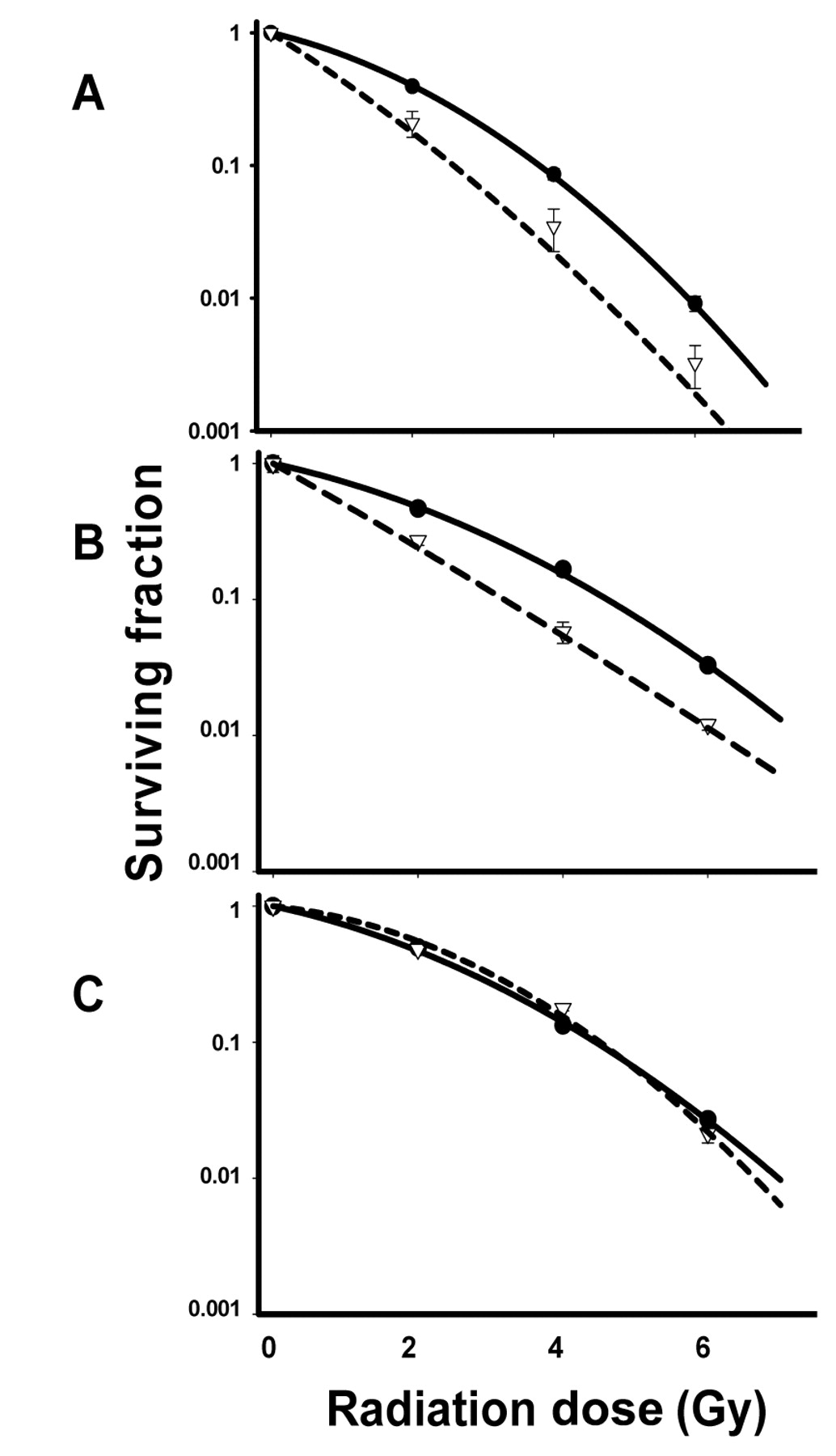
Effect of HCS on radiation sensitization. H460 (A), A549 (B), and H1299 (C) cells were treated with 20 mg/ml HCS for 24 h and exposed to a single radiation dose of 2, 4, or 6 Gy. Cells were then trypsinized and plated in specified numbers in drug-free medium, and 12 to 14 days later, colonies were stained and counted. Percentages of surviving cell colonies were normalized against the cytotoxicity induced by HCS alone. Clonogenic cell survival assay of radiation only (filled circle with solid line), HCS and radiation combination (open triangle with dash line) were plotted in logarithmic scale. Values shown are the means±SE from three independent experiments. Cell survival fraction of 0.1, the enhancement factors were 1.50 for H460 cells 1.44 for A549 cells.
Next we investigated whether HCS affected the extent of radiation-induced DNA double-strand breaks, measured by γH2AX foci formation, and the ability of the cells to repair this damage. Cells were treated with 20 mg/ml for 24 h, irradiated with 2 Gy, and analyzed for nuclear γH2AX foci formation at 0.5, 4, 16, or 24 h later. Figure 3 illustrates the average numbers of γH2AX foci per nucleus as a function of time after radiation only (control) and after HCS plus radiation. At 30 minutes after 2 Gy of radiation only (without HCS), the number of foci per nucleus was 10.3±1.23 in H460 cells, 10.1±0.35 in A549 cells, and 12.7±0.66 in H1299 cells. The numbers of foci declined over time to a similar extent in all three cell lines. HCS by itself did not induce nuclear γH2AX foci formation and had no effect on the number of foci at 30 minutes after the cells were treated with radiation (Figure 3). However, HCS significantly delayed the reduction in the number of radiation-induced γH2AX foci in H460 and A549 cells, but not in H1299 cells. At 4 h, mean number of foci in H460 cells was 4.9±0.31 after irradiation only and 9.3±0.74 after HCS plus irradiation (p<0.001), and in A549 cells it was 3.1±0.27 after irradiation only and 9.2±1.20 (p<0.001) after HCS plus irradiation. Nuclear fluorescence stains of γH2AX foci in H460 cells at 30 minutes and 4 h after the various treatments are shown in Figure 3. At 24 h after irradiation, the difference between irradiation only and HCS plus irradiation was significant for H460 cells (p=0.001) and was not significant for A549 cells (p=0.066). These results suggest that at least in H460 cells inhibition of DNA repair may be one of the mechanisms by which HCS radiosensitized tumor cells.
Finally, to determine whether HCS increased the susceptibility of cells to radiation-induced apoptosis, we treated H460 and A549 cells with 20 mg/ml HCS for 24 h, exposed them to 2 Gy or 4 Gy, and replaced the medium containing HCS with fresh medium, or combination of both. TUNEL assay was performed to determine the apoptotic index 24 h later. HCS by itself only slightly induced apoptosis in both cell lines; in combination with radiation, it enhanced radiation-induced apoptosis only in H460 cells. HCS increased the percentage of apoptosis in H460 cells from the control value of 4.2%±0.9% to 5.2%±1.2% (p=0.668); 2 Gy increased it to 5.2%±0.5% (p=0.595); and 4 Gy increased it to 10.5%±2.7% (p=0.138). When the two agents were combined, the percentage of apoptotic cells increased sharply, to 19.8%±1.7% apoptotic cells after HCS plus 2 Gy (p=0.016 vs. radiation only, p=0.028 vs. HCS only) and to 43.4%±1.1% after HCS plus 4 Gy (p=0.005 vs. radiation only; p=0.001 vs. HCS only) (Figure 4A).

Effect of HCS on the kinetics of nuclear γH2AX formation assessed by immunocytochemical analysis. Sub-confluent cells growing on a coverslip were treated with 20 mg/ml HCS for 24 h followed by a single 2-Gy dose of radiation. Cells were then incubated in drug-free medium for 0.5, 4, 16, or 24 h and then fixed and stained for γH2AX foci using antibody specific to γH2AX as described under the Materials and Methods. Top panel: Representative H460 nuclei from various treatment conditions and time points from three independent experiments are shown. Blue stain: DNA stained with DAPI. Green stain: γH2AX foci stained with FITC. Bottom panel: Quantification of γH2AX foci after exposing the cells to irradiation (black bar), irradiation plus HCS (gray bar): 300-500 cells were evaluated for each time point after treatments. Mean±SE number of foci per nucleus are shown. *p≤0.001.
These findings were further supported by Western blot analyses of some of the cellular proteins of H460 cells, involved in apoptotic signaling and DNA damage repair, namely, cleaved PARP, p53, cleaved caspase-3, and BCL-2. Radiation alone increased cleaved casepase-3 expression at 24 h later by 6.3-fold over the control level, demonstrating that tumor cells were undergoing apoptosis after radiation. HCS alone and HCS plus irradiation up-regulated cleaved caspase-3 expression to 13.0- and 10.8-fold, respectively, at that time point, indicating more cells underwent apoptosis with these treatments compared to radiation alone. BCL-2 expression was slightly decreased 24 h after exposing the cells to HCS or irradiation alone. The combination treatment with both HCS and irradiation potently suppressed BCL-2 expression to 0.3-fold 24 h after irradiation (Figure 4B). A marker of apoptosis, cleaved PARP, was slightly increased by HCS alone and strongly increased by 2.9-fold in the HCS plus irradiation-treated cells 4 h after irradiation. The expression of p53 protein was significantly increased by 4 Gy of radiation alone. Interestingly, HCS blocked the radiation-induced p53 expression in the combination treatment (Figure 4B).

Effect of HCS on radiation-induced apoptosis. A: Cells were treated with 20 mg/ml HCS and irradiated 24 h later with a single 2-Gy or 4-Gy dose. TUNEL assay by flow cytometric analysis showed that HCS increased the percentage of apoptotic cells after 4 Gy in H460 cells (**p<0.01) but not in A549 cells. B: Western blots showing the expression levels of caspase-3, BCL-2, PARP and p53, after 4 Gy, HCS, or HCS plus 4 Gy irradiation in H460 cells. Numbers shown below the blots represent the relative density ratios of bands compared with control (C). Blots shown are representative of three independent experiments.
Discussion
The results of the current study demonstrated that 24-h treatment with HCS led to dose-dependent cytostatic or cytotoxic responses, with an IC50 of approximately 20 mg/ml for H460 (p53 wt), A549 (p53 wt), and H1299 (p53 null) human lung cancer cell lines, suggesting that this effect of HCS appeared to be independent of the p53 status. However, a 24-h pretreatment with HCS strongly enhanced the radiosensitivity of H460 and A549 cells, with enhancement factors of 1.50 and 1.44, respectively, at 0.1 survival fraction, with a dose range of 2-6 Gy γ-radiation. This radiosensitizing effect was stronger than the reported enhancement values of other cardiac glycosides such as ouabain (33) and oleandrin (34). HCS did not affect the radiosensitivity of H1299 cells, suggesting a role for p53 in mediating cellular response to HCS-induced radiation sensitivity. Moreover, this radiosensitization effect depended on the drug-radiation sequence. In contrast to ouabain, which can enhance radiation-induced cytotoxicity when given either before or shortly after irradiation (31-33), HCS had to be given 24 h before the irradiation to confer the radiosensitization effect.
We also investigated the mechanisms that may underlie the radiosensitization effect of HCS on H460 and A549 cells. Specifically, we investigated whether HCS acted by changing the cell cycle distribution at the time of radiation delivery, by rendering DNA more vulnerable to radiation-induced DNA breakage, by slowing the DNA repair process, or by making tumor cells more susceptible to radiation-induced apoptosis. Our results showed no change in cell cycle distribution, thus excluding cell cycle redistribution as a mechanism for the observed radiosensitization. As for DNA damage and repair, we found that pretreatment with HCS significantly delayed DNA repair indicated by prolonged presence of nuclear γH2AX foci at least in H460 cells, suggesting that HCS may have enhanced radiosensitivity by inhibiting DNA repair, as has been reported for other cardiac glycosides, including bufalin and ouabain (33, 36). Interestingly, this radioenhancement effect in our study may be dependent on the mutational status of p53, an oncogene involved in DNA repair and induction of apoptosis. Our analysis of the apoptotic index further supported these findings, although the enhancement of radiation-induced apoptosis was significant only in H460 cells. These results are consistent with our previous findings that oleandrin enhanced the radiosensitivity of PC-3 human prostate carcinoma cells by increasing the sensitivity of those cells to radiation-induced apoptosis (34). Induction of apoptosis has also been implicated in the ability of ouabain to enhance radiosensitivity in several other human tumor cell lines (32).
At the molecular level, cardiac glycosides have been found to affect the expression of cyclin A, BCL-2, BCL-xL, p21, BAX, and c-MYC (18, 24, 25). They also increase apoptosis in some cancer cells by influencing the activity of caspase-3 and caspase-9 (17). In the current study, Western blot analysis of proteins involved in apoptotic signaling showed that treating H460 cells with both HCS and radiation led to increased expression of cleaved caspase-3 and cleaved PARP, and reduced expression of BCL-2 and p53, which strengthens our conclusion that HCS increases the susceptibility of cells to radiation-induced apoptosis in at least H460 cells.
In conclusion, these findings represent the first evidence that HCS enhances the radiosensitivity of human lung cancer cell lines and to a greater extent than that reported elsewhere for other cardiac glycosides. We further found that these effects may have been related to p53 status, as the radiosensitization was not observed in p53-null cell line. The underlying mechanisms may be multiple and cell type-dependent, and may involve inhibition of DNA repair and induction of apoptosis. These findings suggest that HCS has the potential to improve the efficacy of radiotherapy and that it warrants further investigation in animal xenograft models.
Acknowledgements
These studies were supported in part by the National Institutes of Health through MD Anderson's Cancer Center Support Grant CA016672.
Footnotes
-
This article is freely accessible online.
- Received April 8, 2011.
- Revision received May 18, 2011.
- Accepted May 19, 2011.
- Copyright© 2011 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved





{kind=link}
{kind=link}
{kind=link}
{kind=link}