


Abstract
Background: Carcinoid tumors are associated with the carcinoid syndrome, a set of symptoms resulting from the peptide and amine products, including serotonin, secreted from the cancer cells. The purpose of this study was to investigate the relationship between the phosphatidylinositol-3-kinase/protein kinase B (PI3K/Akt) inhibitor PTEN (phosphatase and tensin homolog deleted on chromosome ten) and serotonin synthesis and secretion in the carcinoid cancer cell line BON. Materials and Methods: PTEN was inhibited by pharmacological and molecular approaches, and the resultant secretion of serotonin and expression of tryptophan hydroxylase 1 (TPH1), the rate-limiting enzyme in serotonin synthesis, was assessed. Results: Inhibition of PTEN in vitro, with concomitant increased Akt signaling, resulted in decreased secretion of serotonin, as well as decreased serotonin synthesis, as confirmed by reduced expression of TPH1. Inhibition of PTEN in BON cells in an animal model resulted in decreased serum serotonin. Conclusion: By inhibiting signaling through Akt, PTEN indirectly promotes serotonin synthesis and secretion.
Carcinoid tumors are slow growing neuroendocrine neoplasms that have been increasing in incidence since the 1970s (1). These tumors arise from enterochromaffin cells lining the gut and secrete amine and peptide products, including chromogranin A (CgA) and serotonin (2). Derived from the amino acid tryptophan, serotonin is a neurotransmitter that enhances bowel motility and, in excess, can lead to severe diarrhea (3). The rate-limiting enzyme in serotonin synthesis is tryptophan hydroxylase (TPH), which converts tryptophan to 5-hydroxy-L-tryptophan (5-HTP) (4). Two isoforms of TPH exist: TPH1 is normally only expressed in the pineal gland, spleen, thymus and enterochromaffin cells of the gut, while TPH2 is generally confined to the brainstem (4). Serotonin is hypothesized to play a significant role in the development of carcinoid syndrome, and many treatments for the symptoms inhibit the release of serotonin and other hormones from carcinoid tumors. Unfortunately, due to their indolent nature, many carcinoids are not detected until they have reached an advanced stage (5) and generally do not respond well to standard chemotherapeutic agents (6).
The development of novel treatments for carcinoid tumors has focused on targeting growth factor receptors and oncogenic cellular signaling pathways (6). Phosphatidylinositol-3-kinase/protein kinase B (PI3K/Akt) signaling is overactivated in many types of cancer, including carcinoid tumors (7, 8), through either constitutive activation of PI3K, which indirectly promotes Akt phosphorylation and activation, or down-regulation or mutation of phosphatase and tensin homolog deleted on chromosome ten (PTEN), the endogenous inhibitor of PI3K/Akt signaling. PTEN converts the membrane phospholipid phosphatidylinositol-3,4,5-triphosphate to phosphatidylinositol-4,5-bisphosphate, thereby removing the binding site for Akt, where it is phosphorylated and activated (9, 10). While the role of Akt signaling in tumor progression and metastasis is well established, the role of the PI3K/Akt pathway in carcinoid secretion or the carcinoid syndrome (a set of symptoms resulting from carcinoid peptide and amine products) has not been investigated.
Our laboratory established and characterized the BON cell line, which was derived from a lymph node metastasis of a pancreatic carcinoid tumor (11, 12). BON cells synthesize and secrete neurotensin, pancreastatin, CgA and serotonin (11, 13-15). Various growth and cell signaling inhibitors are noted to inhibit BON cell growth in vitro and in vivo (12). Recently, we have developed an animal model in which splenic injection of BON cells into athymic nude mice leads to the development of primary and metastatic tumors and produces symptoms consistent with carcinoid syndrome including increased serum serotonin concentration (16). Additionally, we previously demonstrated that BON cells with stable reduction of PTEN have a higher metastatic potential using an in vivo model (17). The purpose of the current study was to assess the relationship between PI3K/Akt/PTEN signaling and serotonin secretion and synthesis in the BON carcinoid cell line.
Materials and Methods
Cell culture. The BON cell line was last authenticated in October 2009 at the Johns Hopkins Genetic Resources Core Facility with short tandem repeat analysis using an Identifiler identification kit (Applied Biosystems, Carlsbad, CA, USA). The BON cells were maintained in DMEM/F12K (50/50) medium (Mediatech Inc., Herndon, VA, USA) supplemented with 5% fetal bovine serum (Atlanta Biologicals, Lawrenceville, GA, USA) in 5% CO2 at 37°C.
PTEN and Akt inhibition. To reduce PTEN expression, BON cells were transfected with a pGIPZ™ lentiviral vector (Open Biosystems, Huntsville, AL, USA) including short-hairpin (sh)RNA to PTEN and markers for green fluorescent protein (GFP) and puromycin resistance. These cells were designated BON shPTEN. BON cells transfected with a pGIPZ™ lentiviral vector encoding scrambled shRNA (BON shControl) were used for comparison. Briefly, BON cells were transfected at 60-80% confluence. Transfection complexes were made by mixing 10 μg Arrest-In™ transfection reagent (Open Biosystems) diluted in 50 μl serum-free DMEM/F12K medium with 2 μg shRNA construct diluted in 50 μl serum-free DMEM/F12K medium. Complexes were incubated at room temperature for 10 min and then added to cells plated in 6-well tissue culture dishes in 2 ml serum-free DMEM/F12K medium. Cells were incubated with transfection complexes for 6 h at 37°C in 5% CO2. Medium was aspirated and replaced with 5% serum-supplemented medium. After 48 h, cells that incorporated the shRNA constructs were selected in medium containing puromycin (2 μg/ml). Alternatively, the phosphatase activity of PTEN was inhibited with the 3-hydroxypicolinate vanadium (IV) complex VO-OHpic trihydrate (18) purchased from Sigma (St. Louis, MO, USA). The VO-OHpic trihydrate stock was dissolved in ddH2O and used at a concentration of 500 nM.
The isoform-selective Akt Inhibitor VIII and Akt Inhibitor XII were obtained from Calbiochem® (San Diego, CA, USA). Of the three Akt isoforms, Akt Inhibitor VIII has the highest selectivity for Akt1, while Akt Inhibitor XII is most selective for Akt2. Stock solutions of the Akt Inhibitor VIII and Akt Inhibitor XII were dissolved in DMSO and ddH2O, respectively, and used at concentrations of 500 nM and 1 μM, respectively.
Western blot analysis. Total protein was resolved on NuPAGE® 4-12% Bis-Tris gels (Invitrogen, Carlsbad, CA, USA) and transferred to Sequi-Blot™ PVDF membranes (Bio-Rad, Hercules, CA, USA). The membranes were incubated with specific primary antibodies against CgA, PTEN, pAkt, Akt and β-actin (loading control) (Cell Signaling Technology Inc., Danvers, MA, USA). Antibody against TPH1 was obtained from Abcam Inc. (Cambridge, MA, USA). Following incubation with a horseradish peroxidase (HRP)-conjugated secondary antibody (Santa Cruz Biotechnology Inc., Santa Cruz, CA, USA), proteins were visualized using an enhanced chemiluminescence (ECL™) detection system (GE Healthcare, Pittsburg, PA, USA).
Serotonin and 5-HIAA ELISA. To quantify the serotonin produced by the BON cells, a serotonin ELISA (Immuno-Biological Laboratories Inc., Minneapolis, MN, USA) was performed with BON cell conditioned media according to the manufacturer's directions. BON shPTEN and shControl cells were plated at a density of 5×104 cells/cm2 and cultured for 24 h. Then the medium was removed, and cells were washed and serum starved in serum-free McCoy's 5A medium for 24 h. McCoy's 5A medium contains ascorbic acid, which protects serotonin from oxidative degradation. To assess the effect of pharmacological inhibition of PTEN and Akt on serotonin secretion and synthesis, parental BON cells were cultured and serum-starved under the same conditions, with the addition of VO-OHpic trihydrate, Akt Inhibitor VIII, or Akt Inhibitor XII at the beginning of serum starvation in McCoy's 5A medium. After the medium was collected, the cells in each flask were counted and the serotonin concentration in the medium was normalized to the number of cells. From these values, the serotonin levels in the conditioned media relative to the control groups were calculated. Using the above ELISA kit, serotonin levels were also assessed in serum collected from mice. To account for differences in the number of physiological and pathological serotonin secreting cells in each mouse, serotonin concentration was normalized to total body weight. A 5-hydroxyindoleacetic acid (5-HIAA) ELISA kit from DRG Diagnostics (Mountainside, NJ, USA) was used to evaluate the 5-HIAA (serotonin metabolite) in the urine. The ELISA with the BON cell supernatant was performed in quadruplicate, while the ELISAs with the animal samples were performed in duplicate.
Immunohistochemistry. Serial tissue sections were incubated overnight in TPH1 or CgA primary antibodies diluted in antibody diluent, as described previously (19). Briefly, the sections were stained, and the intensity of TPH1 staining relative to CgA expression was assessed in a blinded fashion by an experienced pathologist. Protein staining was performed using an Envision+® System-HRP (DAB) kit (Dako, Carpinteria, CA, USA); the samples were counterstained with hematoxylin.
Animal studies. Male athymic nude mice (4-6 weeks) were purchased from Harlan-Sprague-Dawley (Frederick, MD, USA). The mice were anesthetized with isoflurane and BON shPTEN or BON shControl or parental BON cells (1×107 per 100 μl) were injected into the pancreas or spleen with a 27-gauge needle as described previously (16, 17). The mice were sacrificed 10 weeks following injection. Trunk blood was collected from the mice following sacrifice, centrifuged, and the top layer (serum) was collected for analysis. Urine was collected from the mice immediately prior to sacrifice. The primary and metastatic tumors were excised from the mice and paraffin embedded. Additionally, cells from liver metastases (designated BON shPTEN metastasis) were harvested and cultured before being assessed for serotonin secretion. All studies were approved by the Institutional Animal Care and Use Committee of the University of Kentucky.
Statistical analysis. Descriptive statistics including means and standard deviations were calculated and displayed in bar graphs to summarize the ELISA measurements across cell culture and animal treatment and control groups. Two-sample t-test or analysis of variance was performed for comparison of two or multiple groups, respectively. Assumptions on data normality and equality of variance were verified to determine validity of the statistical tests. The null hypothesis was rejected when p<0.05. Data analysis was conducted using SAS® Release 9.2 statistical software (SAS Institute Inc., Cary, NC, USA).

Effect of reduction in PTEN expression and activity on serotonin synthesis and secretion. A: Relative secretion of serotonin in BON cells with stable reduction of PTEN (PTEN shRNA) compared to control shRNA BON cells (*p=0.002); BON shPTEN metastatic (PTEN shRNA met) cells harvested from liver metastases following pancreatic injection of BON shPTEN cells compared to control shRNA BON cells (*p<0.0001; (p): number of passages after being harvested from a liver metastasis). B: Western blot analysis of BON cells with stable reduction of PTEN and BON shPTEN metastatic cells compared to control. C: Serotonin secretion following inhibition of PTEN activity with VO-OHpic trihydrate (*p=0.016, 0.026 and <0.0001 after treatment for 6 h, 24 h and 48 h, respectively vs. treatment with vehicle control) and corresponding Western blots.
Results
Effect of PTEN knockdown on serotonin secretion and synthesis. We examined the relationship between PTEN and serotonin secretion and synthesis and demonstrated with a serotonin ELISA that the BON cells with stable reduction of PTEN (BON shPTEN) secreted approximately half the level of serotonin compared to the BON shControl cells (Figure 1A; p=0.002). The secretion of serotonin from BON shPTEN metastasis cells from two different passages was approximately 100 times less than that of the BON shControl cells (Figure 1A; p<0.0001).

Effect of reduction in PTEN expression in an in vivo model. Control shRNA or PTEN shRNA BON cells were injected into the pancreas of athymic nude mice (n=10) and samples collected at sacrifice after 10 weeks. A: Serum serotonin concentration relative to total body weight (*p=0.012). B: Urine concentration of the serotonin metabolite 5-HIAA relative to total body weight (*p=0.0059). Immunohistochemically stained representative tumor sections after BON shPTEN or BON shControl cells were injected into the pancreas (×100) (C) or parental BON cells were injected into the spleen (×200) (D).
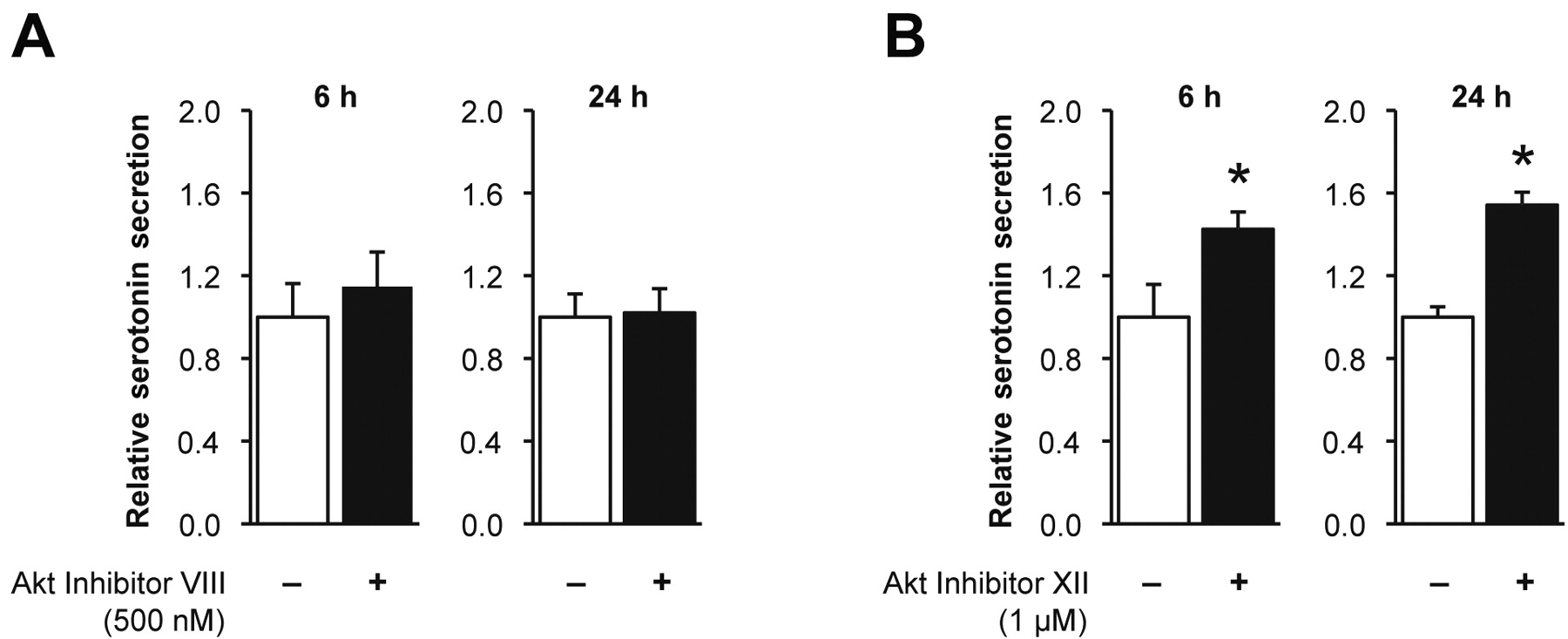
Effect of Akt1 and Akt2 on serotonin secretion. A: BON cells treated with Akt1 selective inhibitor Akt Inhibitor VIII. B: Selective inhibition of Akt2 with Akt Inhibitor XII compared to treatment with vehicle control (*p=0.003 at 6 h and p<0.0001 at 24 h).
Western blot analysis demonstrated that PTEN reduction, associated with a concomitant induction of phospho-Akt, resulted in decreased TPH1 production (Figure 1B). Consistent with the serotonin ELISA results (Figure 1A), the expression of TPH1 was minimal to non-existent in BON shPTEN metastasis cells.
Effect of pharmacological inhibition of PTEN. A single treatment with VO-OHpic trihydrate decreased the BON cell serotonin secretion to approximately 74%, 78% and 90% of the vehicle-treated cells after treatment times of 6, 24 and 48 h, respectively (Figure 1C; p=0.016, 0.026 and <0.0001 at 6 h, 24 h and 48 h, respectively). While the pharmacological inhibition of PTEN, with concurrent enhancement of Akt signaling, significantly reduced serotonin secretion, it did not alter the secretion of serotonin to the same extent as with inhibition of PTEN using shRNA (Figure 1A). Additionally, the expression of TPH1 was not altered following the treatment with VO-OHpic trihydrate (Figure 1C; bottom). Taken together these results suggest that transient PTEN inhibition, with concomitant Akt induction, reduces serotonin secretion, while stable reduction in PTEN expression decreases serotonin secretion and synthesis.
Effect of PTEN reduction in vivo. Since we noted a positive relationship between PTEN expression and activity on serotonin secretion, we assessed the effect of PTEN in an in vivo carcinoid model. The average serum serotonin concentration relative to total body weight in the mice injected with the BON shPTEN cells was significantly less than in the mice injected with the BON shControl cells (Figure 2A, p=0.012). Consistent with this observation, the concentration of 5-HIAA in the urine relative to total body weight was also significantly lower in the mice that received injections of the BON shPTEN cells compared to those injected with the BON shControl cells (Figure 2B, p=0.0059). Immunohistochemical analysis of tissue sections demonstrated that relative to CgA expression, the expression of TPH1 was similar in the primary tumors derived from the BON shControl cells and BON shPTEN cells (Figure 2C). However, the metastatic tumors derived from the BON shPTEN cells demonstrated decreased expression of TPH1, compared to the primary tumors, which is consistent with the results in Figure 1B. To further evaluate differences in TPH1 expression in primary and metastatic tumors, parental BON cells were injected into the spleen of athymic nude mice. Liver metastases derived from the parental BON cells demonstrated decreased TPH1 expression compared to the corresponding primary tumors (Figure 2D).
Effect of inhibition of Akt on serotonin secretion. We assessed the effect of Akt1 and Akt2 on BON cell serotonin secretion by isoform-selective inhibition of Akt. At 6 h and 24 h following a single treatment with 500 nM Akt Inhibitor VIII (Akt1 inhibitor), there was no difference in serotonin secretion compared to vehicle treatment alone (Figure 3A, p=0.237 and 0.812, respectively). However, treatment with 1 μM Akt Inhibitor XII (Akt2 inhibitor) significantly increased serotonin secretion after 6 h (1.4 times, p=0.003) and 24 h (1.5 times, p<0.0001) (Figure 3B) compared to vehicle treatment alone.
Discussion
An inverse relationship between PI3K/Akt signaling and serotonin synthesis and secretion from the BON carcinoid cell line was demonstrated, and the in vivo studies further suggested that the secretion of serotonin decreased in more aggressive carcinoid tumors. A possible explanation for these observations is that BON cells may lose their endocrine properties as they become more metastatic.
One of the central hypotheses of cancer biology is that cellular dedifferentiation is concomitant with tumor progression (20), and deletion of Pten has been demonstrated to induce the dedifferentiation of cells into a ‘cancer stem cell’ phenotype (21). By inhibiting the tumorigenic activity of PI3K/Akt signaling, PTEN may also contribute to the maintenance of the differentiated, secretory state of BON cells. One of the markers of the differentiated carcinoid phenotype is TPH1, the rate-limiting enzyme of serotonin synthesis, and overactivation of the Notch signaling pathway, known to inhibit endocrine differentiation, represses TPH1 expression in BON cells (22). Furthermore, TPH expression is restricted to cells whose distinct function is serotonin synthesis, and in serotonergic neurons, its expression is regulated by several transcription factors (23, 24). Gata3, a transcription factor that regulates TPH expression, is down-regulated, while PI3K/Akt signaling is enhanced, in metastatic breast carcinomas (25). Interestingly, the inhibition of Akt1 had no effect on serotonin secretion in the present study, while inhibition of Akt2 increased serotonin secretion from the BON cells. While the role of Akt2 in cancer progression and metastasis is well-established, and we have previously determined that Akt2 promotes colon cancer metastasis (26), its role in secretion is less well understood, and further investigation is warranted. As with the present Akt2-mediated inhibition of BON cell serotonin secretion, Akt2 has also recently been shown to be a suppressor of gastric acid secretion (27, 28).
The molecular inhibition of PTEN concomitantly reduced TPH1 expression, while the pharmacological inhibition of PTEN decreased serotonin secretion, but not its synthesis. A possible explanation is that pharmacological inhibition of PTEN only transiently increases PI3K/Akt signaling, due to degradation of the inhibitor, thereby affecting secretion, but not gene transcription. Consistent with the in vitro studies, the mice with tumors derived from the BON shPTEN cells secreted less serotonin and excreted less 5-HIAA than the mice with tumors derived from the BON shControl cells. Interestingly, the expression of TPH1 in primary tumors derived from the BON shControl and BON shPTEN cells was similar. However, the metastatic tumors had decreased expression of TPH1 compared to the primary tumors, suggesting that as the carcinoid cells adopted a more metastatic phenotype a concurrent loss in the secretory phenotype occurred. This could be explained by the complexity of the regulation of serotonin synthesis and secretion in cancer cells. It has been demonstrated in breast carcinomas that serotonin has tumor suppressive and tumor promoting actions and that the expression of TPH1 has a nonlinear association with tumor stage (29). As breast tumors grow, they lose expression of TPH1, regaining TPH1 expression as the tumors become invasive (29). The hypothesis for this phenomenon is that early on in breast tumor development, TPH1 expression is reduced and tumors can evade the tumor-suppressive functions of serotonin, such as growth inhibition and apoptosis, and maintain a proliferative state. Since serotonin has also been demonstrated to promote epithelial-to-mesenchymal transition (30, 31), the increase in TPH1 expression as tumors expand is proposed to be a marker of increased metastatic capability (29). We noted previously that carcinoid tumors with reduced PTEN expression have an increased metastatic potential (17). Thus, in the more aggressive BON shPTEN primary tumors, it is possible that during the first weeks of tumor growth, the expression of TPH1, and, hence, secretion of serotonin, was much lower than at the time of sacrifice and more comparable to the in vitro results, shown in Figure 1. At the time of sacrifice, cells from the BON shPTEN tumors were becoming more invasive and, consequently, the expression of TPH1 may have increased. After tumor cells migrate to a distant site, they undergo mesenchymal-to-epithelial transition to form a micrometastasis resembling the primary tumor (32). Thus, the decreased expression of TPH1 in the carcinoid liver metastases might have been associated with the resurgence of a proliferative, non-invasive phenotype.
While carcinoid tumors generally maintain a differentiated phenotype, the carcinoid syndrome does not present until liver metastases have developed and the hormone products secreted by the cancer cells are able to bypass degradation in the liver. Paradoxically, the metastatic tumors in the liver were observed to have decreased expression of TPH1. It is possibly that the secretory products responsible for carcinoid syndrome are actually secreted from the primary tumor. Carcinoid tumors are highly vascular (33, 34) and upon angiogenesis may receive blood from the systemic, in addition to the portal circulation, enabling the secreted products to circulate throughout the body without being degraded in the liver.
In summary, while enhanced Akt signaling, through PTEN inhibition, increases the metastatic potential of BON cells, decreased PTEN expression reduces serotonin synthesis. This novel function of the complex PI3K/Akt/PTEN cell signaling pathway may be useful in the development of new treatments for carcinoid disease and the carcinoid syndrome. Furthermore, the expression of TPH1 and secretion of serotonin from carcinoid cells is directly related to the activity of PTEN. These findings suggest the need for a multi-targeted approach in treating carcinoid tumors, as targeting the PI3K/Akt pathway may inhibit the growth of the tumor, but also result in enhanced secretion of serotonin and other peptide products, which may have deleterious clinical effects.
Acknowledgements
The Authors would like to thank Donna Gilbreath, Jennifer Rogers and Nathan Vanderford for manuscript preparation. This work was supported by National Institutes of Health grants R37 AG10885, R01 DK48489, T32DK07639, and R01 CA104748.
Footnotes
-
This article is freely accessible online.
- Received January 7, 2011.
- Revision received March 1, 2011.
- Accepted March 2, 2011.
- Copyright© 2011 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved





{kind=link}
{kind=link}
{kind=link}