


Abstract
Methylating agents, a widely used class of anticancer drugs, induce DNA methylation adducts, the most biologically significant being O6-methylguanine. The efficacy of these drugs depends on the interplay of three DNA repair systems: base excision repair (BER), methyl-directed mismatch repair (MMR) and direct damage reversal by O6-methylguanine-DNA methyltransferase (MGMT). An MGMT-inducible, MMR- and BER-proficient HeLa cell line was treated with different concentrations of N-methyl-N-nitrosourea (MNU), a model SN1 methylating agent, analogous to widely used methylating cancer chemotherapeutic drugs, under different expression levels of the repair enzyme (MGMT). MNU induced MGMT-dependent apoptotic cell death. In this particular cellular context, the induction of apoptosis was accompanied by modifications of the RNA binding protein poly(A)polymerase and significant down-regulation of the heterogeneous nuclear ribonucleoprotein (hnRNP) C1/C2. These results implicate alterations of the above mentioned RNA binding proteins in SN1 methylating agent-induced cell death and apoptosis, providing a possible perspective regarding their use as biomarkers of tumor resistance/sensitivity to chemotherapy.
- SN1 methylating agents
- apoptosis
- O6-methylguanine-DNA methyltransferase (MGMT)
- RNA-binding proteins
- N-methyl-N-nitrosourea
Methylating agents of the SN1 type are highly cytotoxic, mutagenic, recombinogenic and clastogenic; among the lesions induced, O6-methylguanine (O6-meG), although induced in small amounts (maximally 8% of total methylation products), is believed to be the mostly biologically significant (1, 2). A significant proportion of apoptotic death induced by SN1 methylating agents is attributed to O6-meG. As O6-meG mispairs with thymine during DNA replication, the O6-meG:T pairs are recognized by the mismatch repair (MMR) system, which removes the thymine. Repair resynthesis of DNA leads to reinsertion of thymine opposite O6-meG and reinitiation of futile mismatch repair. The DNA double-strand breaks (DSBs) resulting from this process are thought to initiate a series of events, including prolonged G2 arrest, finally leading to cell death and apoptosis (3-5).
A subgroup of these DNA-damaging agents, mainly hydrazine and triazine derivatives such as procarbazine, dacarbazine and temozolomide, are widely used in the chemotherapy of ovarian cancer, malignant melanoma, various hematological and brain tumors (6-8). The efficacy of these drugs is determined by the interplay of three DNA repair systems, base excision repair (BER), MMR and direct damage reversal by O6-methylguanine-DNA methyltransferase (MGMT) (9-10). MGMT, stoichiometrically and selectively removes methyl lesions from the O6 position of guanine, returning the DNA to its pre-lesioned state (11) and seems to be a critical factor influencing the cellular response to O6-meG-generating agents; its expression efficiently protects against their mutagenic, cytotoxic, clastogenic and carcinogenic activity (12).
The elucidation of the molecular mechanism implicated in cell death following DNA damage induced by anticancer genotoxic agents is crucial for the improvement of chemotherapy effectiveness (12). On the other hand, there is growing evidence supporting the implication of RNA-binding proteins, heterogeneous nuclear ribonucleoproteins (hnRNPs) in particular, in the different events of cellular regulation and tumor development including cell death.
The RNA-binding proteins, including hnRNPs, are abundant, multifunctional proteins involved in almost every aspect of RNA metabolism, having important roles in transcription, splicing, transport, translation and stability (14). Recent data provide evidence for the relation of their expression with different events during tumor development. Thus, certain hnRNP proteins can act as oncogenes (hnRNP P) or tumor suppressor genes (hnRNP E4 and G), promoting or inhibiting cancer (15-18). HnRNP proteins can also act as positive or negative regulators of apoptosis: hnRNP C1/C2 is an inhibitor controlling the cellular expression level of X-linked inhibitor of apoptosis protein (XIAP) (19-20) while hnRNP F and H enhance the synthesis of pro-apoptotic regulator Bcl-xS (21). Certain hnRNP proteins have been implicated in tumor progression based on their different expression profiles when comparing normal to cancerous tissues. HnRNP A2/B1 is the one, most extensively studied (15, 22), although reports exist for altered expression levels of hnRNP F, H, I, K and M in different types of cancer (15, 23).
In the present study, we investigated the DNA repair-dependent molecular mechanism of cell death induced by the SN1 methylating agents in cell lines of human origin, along with the concomitant alterations in the expression level of RNA binding protein, in the context of their possible use as markers of tumor resistance/sensitivity to chemotherapy. To this end, a pair of HeLa Tet-On cell lines, differentially expressing the repair enzyme MGMT were treated with equimolar cytotoxic doses of N-methyl-N-nitrosourea (MNU), a model SN1 methylating agent analogous to widely used methylating cancer chemotherapeutic drugs. Cell death was assessed by cytofluorometry of DNA content, microscopy and immunodetection of specific markers.
Materials and Methods
Cell culture and methylating agent treatment. A Hela Tet-On cell line (MGMT deficient, Mer-, MMR proficient) was used to construct an MGMT-inducible clone (HeLa Tet-On/hMGMT or HeLa Tet-On 18 cell line) where the expression of the human MGMT gene is gradually turned on by doxycycline (Dox) (Sigma-Aldrich, St. Luis, Missouri, USA) (Clontech Tet-On™ Gene Expression Systems and Cell Lines, Mountain View, CA, USA). The clonal cell line possesses a background MGMT activity equal to ~3-6 fmol/μg DNA due to Tet-On system background leakage; MGMT activity sharply increases within the range of 1-100 ng Dox /ml culture medium and reaches a plateau at concentrations of 100-1000 ng Dox/ml (>30 fmol/μg DNA). Both cell lines were grown according to the Clontech's User Manual.
MNU (Sigma-Aldrich) was prepared, as previously described (24), by dissolving the agent in dimethyl sulphoxide (DMSO), (Sigma-Aldrich) at a concentration of 20 mg/ml (stock solution) and working solutions were prepared in fresh medium without Fetal Bovine Serum (FBS) (GIBCO, Invitrogen, Carlsbad, CA, USA) wherein the DMSO concentration did not exceed 0.1% v/v. Then the cells were treated for 1 h after which the medium was replaced by fresh medium containing 10% FBS. Cells were harvested at 24, 48, 72 and 96 h after the end of the MNU treatment. Non treated cells were used in all cases as control. All cells were harvested using 0.25% trypsin solution in phosphate buffer saline (PBS) (GIBCO). Equimolar cytotoxic doses were determined by clonogenic cell survival assay as previously described (24). All assays were carried out in triplicate.
Determination of MGMT activity. MGMT activity was determined in the HeLa and HeLa Tet-On cell lines by a standard previously described assay (25) based on reaction with DNA which had been methylated with N-[3H] MNU (21.0 Ci/mmol, Amersham Biosciences, Little Chalfont Buckinghamshire, UK) and subsequently heated to 80°C for 16 h to remove N-methylpurines.
Flow cytometry. Cells were plated at a density of approximately 1.5×105 cells per well of a 6-well plate and allowed to grow overnight. After 24 h, cells were treated with different doses of MNU corresponding to Lethal Dose (LD)50 and LD100 for each cell line (10-200 μg/ml) for 1 h at 37°C. After treatment, at 24, 48, 72 and 96 h, cells were trypsinized and fixed in ice-cold 100% ethanol overnight at 4 °C. Cells were washed with PBS, stained with propidium iodide for 2 h and subjected to Fluoresence –activated cell sorting (FACS) analysis on a FACScan flow cytometer (BD PharMingen, San Diego, California, USA). Data were analyzed using the CellQuest (BD Biosciences, San Jose, CA, USA) and ModFit (Verity Software, Topsham, ME, USA).
Analysis of protein extract. Preparation of total protein extracts, electrophoretic analysis and western blotting were as previously described (24). Primary antibodies against the following proteins were used at the indicated dilutions: PARP (sc-7150; Santa Cruz Biotechnology, Santa Cruz, CA, USA), 1:500; BCL-2 (sc-509; Santa Cruz Biotechnology), 1:500; caspase-3 (sc-7150; Santa Cruz Biotechnology), 1:500; caspase-7 (9494; Cell Signaling Technology, Danvers, MA, USA) 1:500, poly(A)polymerase (T-20) (sc-20188; Santa Cruz Biotechnology) 1:500, β-actin (MAB1501; Chemicon, Temecula, CA, USA), 1:4.000; hnRNPC1/C2(4F4) (sc-32308; SantaCruz Biotechnology), 1:1.000., CHK1 (ab63345; Abcam, Cambridge, UK) 1:500, Phospho-CHK1 (ab2834; Abcam).
Results
Cell death characterization at the cellular level. The morphological changes accompanying cell death induced by MNU were assessed through cytofluorometric analysis of DNA content performed in parallel in both the parental and the clonal cell line after applying MNU doses ranging between the lethal dose (LD)50 to LD100 as assessed by the clonogenic cell survival assay. Cells were harvested and analysed at 24, 48, 72 and 96 h post treatment.
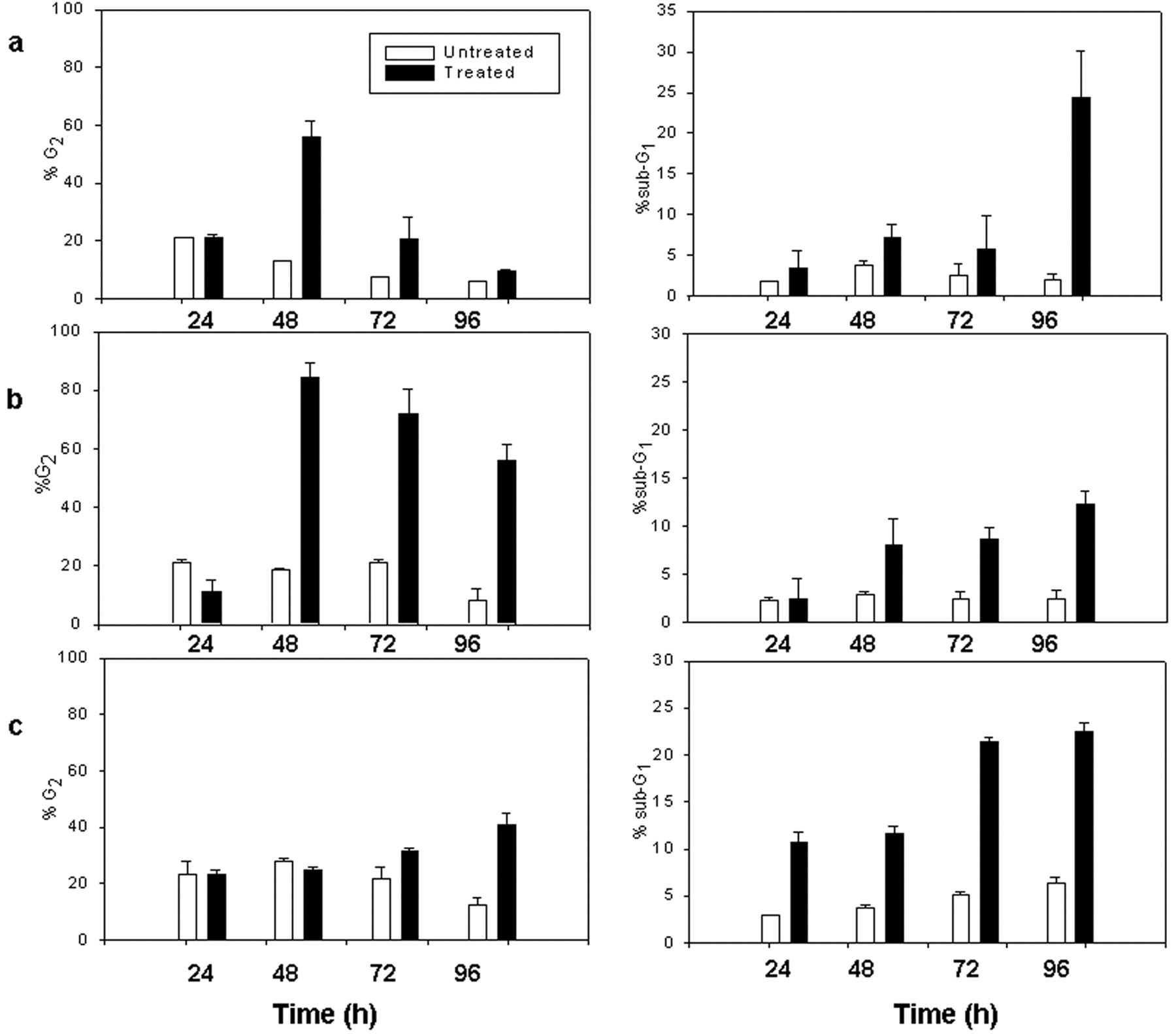
In the HeLa Tet-On cells lacking functional MGMT, the results revealed a G2/M arrest at 48 h and dose-dependent cell cycle alterations. A transient G2/M arrest was observed 48 h after treatment with MNU at LD50 (5 μg/ml), followed by an increase in the sub-G1 fraction at 96 h after treatment (Figure 1a). The increase in the sub-G1 fraction, corresponding to subdiploid cells, is indicative of apoptotic death. When cells were treated with MNU at LD100 (10 μg/ml), a prolonged G2/M arrest, starting at 48 h, was observed, also accompanied by subG1 increase at the same time points (Figure 1b). In the course of parallel cytofluorometric analysis of DNA content in the clonal cell line HeLa Tet-On 18 (MGMT activity: 3-6 fmol/μg DNA), an increase in the G2 fraction, followed by subG1 increase, became evident 72 h post treatment only with MNU at LD100 (200 μg/ml) (Figure 1c).

Effect of N-Methyl-N-Nitrosourea (MNU) treatment on the cell cycle. Untreated and MNU-treated (LD50 or LD100) HeLa Tet-On and HeLa Tet-On 18 cells harvested at 24, 48, 72 and 96 h, were stained with propidium iodide and subjected to cytofluorometric analysis of DNA content. a: In HeLa Tet-On cells, transient G2/M arrest was observed at 48 h after treatment with MNU at LD50, followed by induction of apoptosis (sub-G1 increase) 96 h after treatment. b: When HeLa Tet-On cells were treated with MNU LD100 a prolonged G2/M arrest was observed along with sub-G1 increase starting at 48 h. c: In HeLa Tet-On 18 cells, an increase in the G2 fraction, followed by sub-G1 increase, became evident 72 h post treatment with MNU at LD100. The results are derived from at least three experiments per cell line/time point.
Biochemical analysis of cell death. DNA damage signaling and G2/M arrest: The molecular analysis confirmed the cell cycle analysis, shown in Figure 1, since phosphorylation of CHK1, indicating prolonged G2/M arrest (26), was detected only in the HeLa Tet-On cells at the high MNU dose (LD100) evident 48 h post treatment (Figure 2a). The lack of Chk1 kinase phosphorylation in the HeLa Tet-On cell line with MNU at LD50 (Figure 2a) and the clonal cell line (Figure 2b) is in agreement with the results obtained from cytofluorometry (Figure 1).
Immunodetection of apoptosis through western Blotting: Poly (ADP-ribose) polymerase (PARP) is a nuclear protein implicated in DNA repair one of the earliest proteins targeted for specific cleavage to a 89-kDα fragment during apoptosis. Cleavage of PARP is considered as a sensitive apoptotic index since it can be detected even when very few cells in the total polulation are undergoing apoptosis (27). When MNU doses equivalent to LD50 were applied to parental and clonal HeLa Tet-On cells, with or without Dox induction, PARP cleavage was detectable by immunobloting as early as 48 h and was prominent at 72 h only in HeLa Tet-On cells lacking MGMT activity (Figure 3a).
In HeLa Tet-On cells caspase-3 activation was not detected during the induction of apoptosis by MNU, in accordance with the lack of DNA fragmentation and DNA fragmentation factor 45 (DFF45) proteolysis in this cellular system (data not shown). Instead, activation of caspase-7 was detected (Figure 3b). Caspase-7 is another executioner caspase which compensates for non functional caspase-3 but does not lead to apoptotic DNA fragmentation via proteolysis of DFF45, since caspase-3 seems to be the primary regulator of this process (28). BCL-2 decline was also observed, suggesting activation of the mitochondrial pathway (Figure 3c) (29, 30).

Effect of N-Methyl-N-Nitrosourea (MNU) treatment on the cell cycle checkpoint effector kinase CHK1. Immunodetection of CHK1 phosphorylation in protein extracts of HeLa Tet-On (a) and HeLa Tet-On 18 (b) cells 24, 48 and 72 h after MNU (LD50 and LD100) treatment. Cell extracts from untreated (untr.) cells at the same time points were used as controls. All extracts (40 μg) were immunoblotted with a-phosho-CHK1 antibody detecting the phosphorylated form of CHK1 (82 kDa). The relative abundance of total protein applied was measured by using the amount of β-actin (45 kDa) as control.
Modifications of RNA binding proteins: To evaluate a potential association between the expression levels of hnRNPs with the observed effects of MNU treatment, total cell extracts from both the parental and the clonal cell line collected at selected time points of both were analyzed by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and the respective western blots were probed with available antibodies against PARP and several hnRNP proteins. In parallel, anti-β- actin was used as a marker of protein loading. By immunoblotting, the expression of hnRNP A1, A2, A3, H, K/J and L at 24, 48 and 72 h post MNU treatment displayed no changes, neither during the incubation period nor in comparison with the respective untreated samples (data not shown). However, the analysis revealed a significant decrease in the expression level mainly of the C2 isoform of hnRNP C1/C2 detected in the HeLa Tet-On cell line starting at 24 h post MNU treatment, expression was dramatically reduced to almost zero at 72 h (Figure 4a). It is worth mentioning that this decrease was evident only in the HeLa Tet-On MNU-treated cells, while the treated HeLa Tet-On 18 cells displayed no changes in the expression of hnRNP C1/C2 at the same time points with isotoxic MNU doses (Figure 4a).
Under the described experimental conditions, partial cleavage of PAP was again detected only in the HeLa Tet-On cells lacking MGMT activity starting at 48 h post MNU treatment (Figure 4b). The enzyme PAP is involved in mRNA endoribonucleolytic cleavage of the pre-mRNA at the polyadenylation site followed by synthesis of a poly A tail at the 3’ end (31).
Discussion
In the context of the present study, a pair of HeLa Tet-On cell lines, differentially expressing the repair enzyme MGMT, were treated with equicytotoxic doses of MNU, a model SN1 methylating agent, prototypical of several drugs used for cancer chemotherapy. Apoptotic cell death was assessed and confirmed by cytofluorometry of DNA content, microscopy and immunodetection of specific markers. Alterations occurring in RNA-binding proteins, in particular of hnRNPs, during MNU-induced apoptosis were examined in an effort to assess their possible use as markers of tumor resistance/sensitivity to chemotherapy.

Immunodetection of apoptosis in the HeLa Tet-On cell line: cleavage of Poly(ADP-ribose) polymerase (PARP), caspase-7 proteolytic activation and BCL-2 decline. HeLa Tet-On cells were treated with 5 μg/ml of N-Methyl-N-Nitrosourea (MNU) (LD50) and maintained in culture for the indicated length of time. Cell extracts (40 μg) were immunoblotted with respective antibodies. The relative abundance of total protein applied was measured by using the amount of β-actin as assessed on the same blot as control a: PARP antibody detecting both the intact 116 kDa PARP) and the cleaved 89 kDa PARP fragment. The arrows depict the expected position of the intact 116 kDa PARP and the 89 kDa cleaved PARP fragment and of β-actin. b: Caspase-7 antibody detecting both the intact 38 kDa procaspase-7 and the 20 kDa cleaved fragment. The arrows depict the expected position of the intact 38 kDa procaspase-7 and the 20 kDa cleaved fragment resulting from the proteolytic activation of procaspase-7, and of β-actin. c: BCL-2 antibody detecting the intact 26kDa BCL-2. The arrows depict the expected position of the intact 26 kDa BCL-2 and the position of β-actin.
Since the repair of O6-meG by MGMT is stoichiometrical, returning the DNA to its prelesional state, the enzyme level in tumors is a major factor determining the efficacy of methylating anticancer drugs (12, 32). In dividing cells, DNA damage caused by genotoxic insults results in the activation of cell cycle checkpoints followed by DNA repair to ensure the integrity of the transcribed genome. Checkpoint regulations have evolved to coordinate the response to different types of DNA damage and are mediated through activation of two major protein kinase pathways: the ataxia-telangiectasia mutated (ATM) through checkpoint kinase 2 (CHK2) and the ataxia telangiectasia and Rad-3-related (ATR) through CHK1 (33). SN1 DNA methylating agents are known to elicit a G2/M checkpoint response via an MMR system-dependent mechanism (34). As expected, MNU induced dose- and MGMT-dependent cell cycle alterations. Only in cells lacking functional MGMT (HeLa Tet-On) MNU did induce prolonged G2/M arrest, followed by CHK1 phosphorylation and late apoptotic cell death confirmed by PARP cleavage, caspase-7 proteolytic activation and BCL-2 decline (Figure 3c), suggesting activation of the mitochondrial pathway in this cellular system (29, 30), and RNA-binding protein modifications; significant down-regulation of hnRNP C1/C2 and modifications of the RNA-binding protein PAP were detected. Our results, to our knowledge for the first time, implicate alterations of these RNA-binding proteins in SN1 methylating agent-induced cell death and apoptosis.
The RNA-binding proteins constitute an abundant superfamily of multifunctional proteins that are considered to be important gene regulators. Among them, the hnRNPs have been implicated in a variety of biological functions involving almost every aspect of RNA metabolism, as well as in tumorigenesis (15) and apoptosis. Given these facts, in the present study we searched for deregulated expression of selected members of the hnRNP protein family accompanying the observed effects of MNU treatment in the examined cell cultures. Using an immunochemical technique, we were unable to detect differences in the expression levels of the hnRNP A1, A2/B1, A3, H, K/J and M during the experimental time course in the treated versus the control cells in both cell lines. In contrast, the expression level of hnRNP C1/C2 was progressively reduced to an almost undetectable level at 72 h, but only in the HeLa Tet-On cells lacking MGMT activity, treated with the high MNU dose, while the expression level in the treated HeLa Tet-On 18 cells remained unchanged.
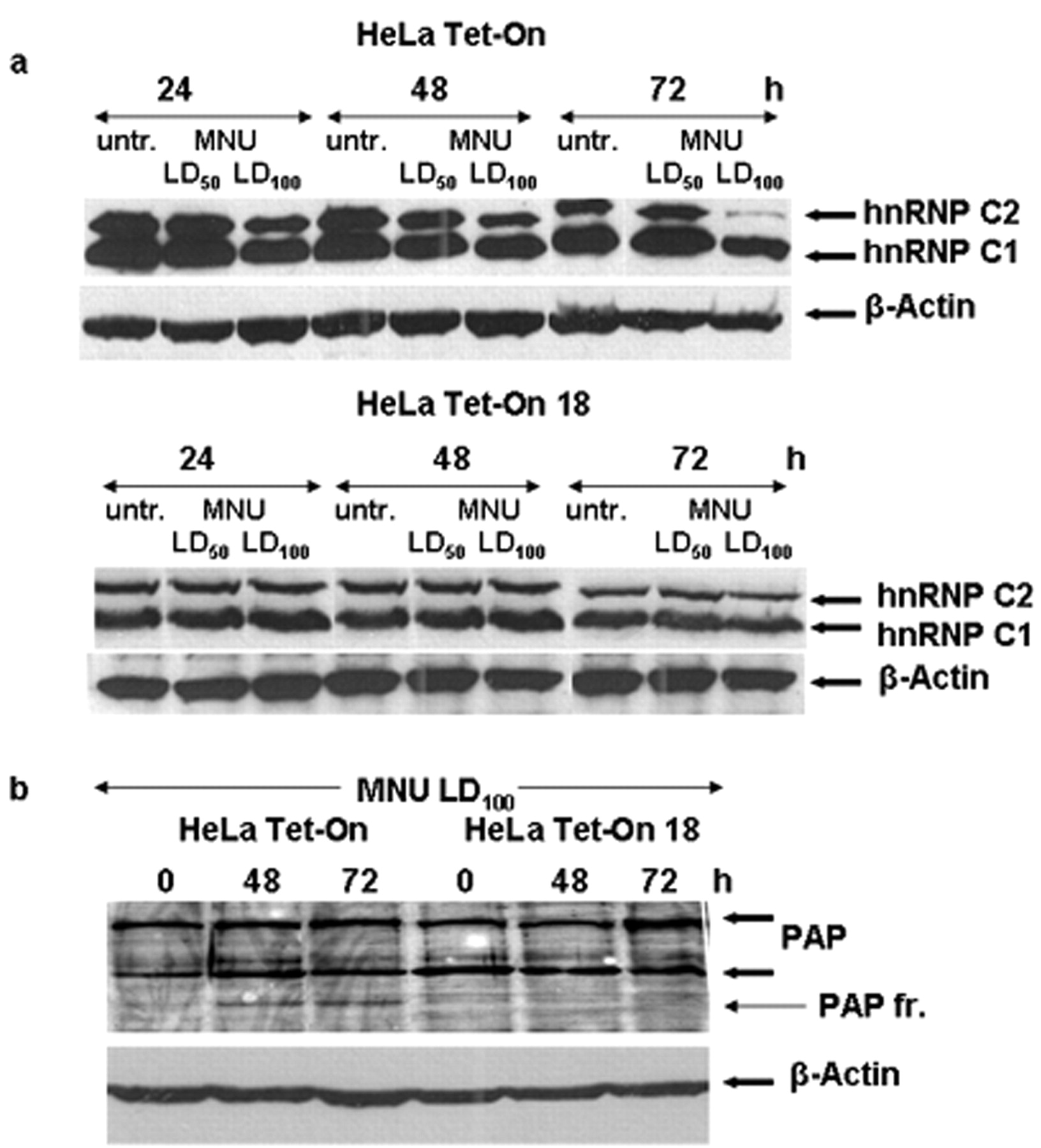
Effect of N-Methyl-N-Nitrosourea (MNU) treatment on the expression of RNA binding proteins in the HeLa Tet-On cellular system. Hela Tet-On and HeLa Tet-On 18 cells, untreated (untr) and treated with isotoxic doses of MNU, were maintained in culture for the indicated length of time after treatment. Protein extracts (40 μg) were immunoblotted with heterogeneous nuclear ribonucleoprotein (hnRNP) C1/C2 (a) and poly(A)polymerase (PAP) antibody (b). The arrows depict the expected position of the relevant proteins. The relative abundance of the total protein applied was measured by using the amount of β-actin as assessed on the same blot as control.
The hnRNP C1 and C2 are isoforms arising by alternative splicing (35) and are involved in regulation of pre-mRNA processing. Their roles are absolutely vital since knockout mice for the hnRNP C1/C2 gene are lethal (36), while stem cells from such mice are viable, but present slow proliferation and lack of ability to differentiate. Moreover, reduction of hnRNP C1/C2 proteins abrogates and decelerates mitosis (37), causes a shift from G1 to G2 phase (indication of defect in G2/M transition) (38) and results in sensitization of cells to various stresses (37). All the above suggest a pleiotropic role of C1/C2 proteins in the progression of the cell cycle and globally in the maintenance of cellular homeostasis. The observed reduction of the expression level of hnRNP C1/C2 proteins in the MNU-treated HeLa Tet-On cells undergoing apoptosis (Figure 4a), is in agreement with previous reports showing either down-regulation or fragmentation of hnRNP C1/C2 proteins in apoptosis induced in vitro (39-41). More specifically, there are indications that hnRNPC1 and hnRNPC2 cleavage during apoptosis may modulate RNA metabolism, as is the case for the translation of the XIAP inhibitor of apoptosis (19, 20). The nature of hnRNP C1/C2 modulation following methylating agent-induced cell death and apoptosis seems to be influenced by the cellular background: significant down-regulation, not accompanied by apoptotic cell death, was observed in A549 non-small cell lung carcinoma cells (p53wt) while up-regulation was observed in NCI-H157 non-small cell lung carcinoma cells (p53null) undergoing apoptosis (24). In the case of the HeLa Tet-On system, it is quite interesting that down-regulation of hnRNP C1/C2, notably of C2, was evident already at 24 h after MNU treatment. This was the only biochemical marker in our study appearing before the activation of the apoptotic cascade became evident. Experiments examining general stress response pathways, have implicated a role for hnRNP C1/C2 in DNA damage response mechanisms. It can bind to Ku protein and can be phosphorylated by the catalytic subunit of the DNA-dependent protein kinase (DNA-PK) complex, suggesting a possible role for hnRNP C1/C2 in DNA DSB repair through the non-homologous end rejoining (NHEJ) repair pathway (42, 43). We believe that the early down-regulation of hnRNP C1/C2 observed in our study at the high MNU dose, where the load of N-alkylation damage is heavy, reflects its role in DNA DSB repair in addition to its involvement in apoptosis.
Upon activation of apoptosis and under the same experimental conditions (in the absence of MGMT), we also observed cleavage of the RNA-binding protein PAP (Figure 4b). Partial proteolysis of PAP occurring during apoptosis has been reported before (31, 44). Moreover, recently, a novel regulatory domain, named the Mitotically Phosphorylated, Apoptotically Cleaved (MPAC) domain has been identified in human U-box-type E4 ubiquitin ligase UFD2A and PAP, which is delineated by apoptotic protease cleavage sites and has a unique amino acid composition, highly enriched in serine, threonine and proline residues. This domain is phosphorylated during mitosis, and is specifically removed by proteolytic cleavage during programmed cell death. Hyperphosphorylation of this domain by CDK1 inhibits PAP catalytic activity (45). Thus, all the above suggest a possible involvement of PAP in cell commitment to and execution of apoptosis.
Although the involvement of the RNA-binding proteins in the apoptotic cascade has already been established, the pattern of their deregulated expression may be differentially affected by the cell type and/or the apoptotic trigger. The modifications of PAP and hnRNP C1/C2 observed in response to MNU-induced cell death and apoptosis in our study provide a promising perspective deserving further investigation. The elucidation of the RNA-binding proteins roles, in particular of those of hnRNPs, holds the promise of a better understanding of the mechanisms which promote cancer, and ultimately may contribute to the design of effective therapeutic treatments.
Aknowledgements
We thank Mrs. Irene Kolonti for excellent technical assistance. We are grateful to Dr. H. Pratsinis (Institute of Biology, Demokritos, Agia Paraskevi, Greece) for useful guidance and discussions throughout the cell cycle analysis experiments. This work was supported by the research project PENED, co-financed by the EU European Social Fund (75%) and the Greek Ministry of Development-GSRT (25%).
Authors state that there are no conflicts of interest with other people or organisations.
- Received July 13, 2011.
- Revision received November 15, 2011.
- Accepted November 16, 2011.
- Copyright© 2011 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.