


Abstract
Background: High-dose chemotherapy (HDC) followed by autologous stem cell transplantation (ASCT) is used for the treatment of hemato-oncologic malignancies. In this study, we measured the effect of HDC/ASCT on plasma concentrations of antiangiogenic soluble vascular endothelial growth factor receptor 1 (sVEGFR1) and of leukapheresis products (LP) and patient serum on chick chorioallantoic (CAM) angiogenesis. Materials and Methods: VEGFR1- and CD34-expressing cells of leukapheresis products were analyzed by flow cytometry. Alternatively spliced isoforms of VEGFR1 mRNA were quantified using reverse transcription PCR. Results: Plasma concentrations of sVEGFR1 decreased after HDC, but significantly increased after ASCT. In the CAM assay, sera of patients elicited a proangiogenic effect before and after HDC, but a strong antiangiogenic response after ASCT, comparable to that of bevacizumab at therapeutic concentrations. LP contains high concentrations of sVEGFR1, and high density of VEGFR1+ neutrophilic granulocytes, in which mRNA expression is shifted toward the soluble VEGFR1 isoform. Conclusion: Neutrophil-derived antiangiogenic sVEGFR1 within the LP may contribute to the therapeutic efficacy of ASCT.
- Alternative splicing
- angiogenesis
- autologous stem cell transplantation
- chick chorioallantoic membrane assay
- high-dose chemotherapy
- placental growth factor
- vascular endothelial growth factor
- VEGFR1
Autologous stem cell transplantation (ASCT) after high-dose chemotherapy (HDC) emerged as a treatment option for hematologic and oncologic malignancies more than fifty years ago (1) and represents the cornerstone for the first-line treatment of younger patients with multiple myeloma (2). It is an accepted therapeutic option in refractory or relapsed lymphoma (3), relapsed testicular cancer (4), and some forms of leukemia (5).
ASCT has also been used for therapeutic angiogenesis in non-oncologic disease, such as ischemic cardiac (6-8) and limb (9) disease, and connective tissue disorders (10-12) in order to improve microcirculation, although randomized controlled trials in ischemic cardiovascular disease reported conflicting data on its beneficial effects (6-8). However, there is considerable heterogeneity in the methods of mobilization, collection, and storage of autologous stem cells.
While there is evidence that autologous bone marrow cell transplantation has proangiogenic effects in the treatment of ischemic disease (10, 13-15), only little is known about the angiogenic potential of ASCT after HDC in the treatment of hematologic and oncologic malignancies (16-19).
Physiological and pathological angiogenesis, including tumor angiogenesis, is regulated by a balance of pro- and antiangiogenic molecules. When the balance shifts in favor of angiogenesis inducers, an angiogenic switch activates the normally quiescent vasculature to form new blood vessels (20). Even a short-lived, transient angiogenic switch can prime progressive tumor growth (21). A prominent proangiogenic signaling circuit involves members of the vascular endothelial growth factor (VEGF) family, in particular VEGFA, and its receptors, VEGFR2 and VEGFR3 (22). VEGFR1, a receptor of VEGFA and placental growth factor (PGF), has strong antiangiogenic activity and its genetic ablation has been shown to be associated with excessive angiogenesis (23). PGF has been shown to act as an amplifier of VEGF-driven angiogenesis (24). Besides being expressed on endothelial cells (25), VEGFR1 is also expressed on a variety of other cell types, including neutrophils (26, 27). Two different isoforms of VEGFR1 arise through alternative splicing: a full-length transmembrane protein containing an intracellular tyrosine kinase domain, and a truncated soluble form, lacking the transmembrane and tyrosine kinase domains (sVEGFR1) (25). Proteolytic cleavage of VEGFR1 may also be involved in the generation of sVEGFR-1 (28). VEGFR1 and sVEGFR1 both exert antiangiogenic effects by sequestration of VEGF, and through a dominant negative effect by forming heterodimers with VEGFR1 and -2 (29).
Patient characteristics.
The aim of this study was to determine the effect of HDC and ASCT on plasma concentrations of sVEGFR1, VEGF, and PGF and on angiogenesis.
Materials and Methods
Patients, blood samples, leukapheresis products, and peripheral blood mononuclear cells. Patients undergoing HDC and ASCT for the treatment of various cancer types were included in the study. The study protocol was approved by the local Ethics Commission (University of Zurich approval number EK-1363) and written informed consent was obtained from all patients. Patient characteristics are shown in Table I. Serum and EDTA plasma samples were taken 6 h before HDC, 4 h before ASCT, 15 min after ASCT, and at the time of hematological reconstitution (neutrophil count ≥500/μl). Following stem cell mobilizing chemotherapy and stimulation with granulocyte-colony stimulating factor (G-CSF), leukapheresis products were collected, supplemented with 10% dimethyl sulfoxide (DMSO) and 33% RPMI cell culture medium and stored in liquid nitrogen until ASCT. For analysis of the leukapheresis products, leukocytes were separated from the leukapheresis product by Ficoll gradient centrifugation, washed three times with phosphate buffered saline (PBS)/5 mM EDTA and filtered through a 100 μm cell strainer. Peripheral blood mononuclear cells (PBMCs) were collected from the blood of cancer patients by Ficoll gradient centrifugation.
Quantification of VEGFR1, VEGF, and PGF protein expression by enzyme-linked immunosorbent assay (ELISA). ELISA for the quantification of VEGFR1, VEGF, and PGF were performed using immunoassays from R&D Systems (Abingdon, UK), and concentrations in plasma and supernatants/medium expressed as pg/ml, or as pg/mg total protein for VEGFR1 concentrations in cell lysates.
Chick chorioallantoic membrane (CAM) assay for the analysis of angiogenesis. The CAM from a shell-less chick embryo culture was used as described earlier (30). In brief, chicken egg contents with intact yolk and embryo were incubated at 37°C and 70% humidity. Methylcellulose disks (usually about 4-7 per CAM) as carriers for test samples were placed on the CAM of day 8.5 chick embryos in duplicates and inoculated with 10 μl of serum (± bevacizumab), leukapheresis product, or freezing medium containing DMSO. Bevacizumab (BV) was used at a final concentration of 500 μg/ml. Angiogenesis was compared by stereomicroscopy between noninoculated CAM areas and areas inoculated with the appropriate control probes after 48 h, following the application of 20% intralipid (Fresenius Kabi, Uppsala, Sweden) underneath the intact CAM to enhance the vascular contrast, and scored by two independent investigators blinded to the experiments: 0, neutral; (+), weak stimulation; +, marked stimulation; ++, strong stimulation; (−), weak inhibition; −, marked inhibition; − −, strong inhibition of vasculogenesis (31). Sera from healthy volunteers were used for comparison.
Magnetic activated cell sorting (MACS) for the separation of CD34- and VEGFR1- positive and negative cells from leukapheresis products. Leukocytes from leukapheresis products were positively selected for CD34+ cells using the CD34 MicroBead Kit according to the manufacturer's protocol (Miltenyi Biotech, Bergisch Gladbach, Germany) and for VEGFR1+ cells using an allophycocyanin (APC)-labeled monoclonal mouse-anti-human VEGFR1 antibody (R&D Systems) and anti-APC beads (Miltenyi Biotech).
Reverse transcription quantitative PCR (RT-qPCR) for quantification of the alternatively spliced isoforms of VEGFR1 mRNA. RT-qPCR was performed for the quantification of the alternatively spliced mRNA isoforms of sVEGFR1 and transmembrane VEGFR1 from snap frozen cells after removal of genomic DNA in an ABI PRISM 7700 Sequence Detector (Applied Biosystems Inc., Foster City, CA, USA) for 40 cycles. Sequences of primers and fluorescent VIC-labeled probes were: sVEGFR1, 5’ TGTTGCAGTGCTCACCTCTGATTGTAATTTCTT 3’ (probe), 5’ GCAAGATTCAGGCAC CTATGC 3’ (sense), 5’ AGATCCGAGAG AAAACAGCCTTT 3’ (antisense); VEGFR1, 5’ TGCTTCCTGATCTCTGATTGTAATTTCTTTCTTCTG 3’ (probe), 5’ GCAAG ATTCAGGCACCTATGC 3’ (sense), 5’ CTGAGGTTTCGCA GGAGGTATG 3’ (antisense).

Enzyme-linked immunosorbent assays for the analysis of plasma concentrations of vascular endothelial growth factor receptor-1 (VEGFR1) (a, percentage change; b, plasma concentration), vascular endothelial growth factor (VEGF) (c), and placental growth factor (PGF) (d) in seven patients. *p<0.05.
Analysis of full blood counts and leukocyte subfractions with blood cell analyzer and cytologic staining. Full blood counts from EDTA plasma and leukocyte subfractions from leukapheresis products, and CD34 and VEGFR1 positively and negatively selected cells were analyzed using an automated blood cell analyzer (ADVIA 120; Siemens, Erlangen, Germany) and by cytologic staining of cytospins with May-Grünwald-Giemsa.
Flow cytometric analysis for the analysis of CD34+ and VEGFR1+ cells. Leukapheresis products, and CD34 and VEGFR1 positively and negatively selected cells of leukapheresis products were stained with a fluorescein isothiocyanate (FITC)-labeled monoclonal mouse-anti-human CD34 antibody (Miltenyi Biotech) and an APC-labeled monoclonal mouse-anti-human VEGFR1 antibody (R&D Systems) and subjected to flow cytometric analysis using a FACSCalibur® flow cytometer, CellQuestPro (Becton Dickinson Biosciences, Franklin Lakes, NJ, USA), and WinMDI Version 2.9 software (Joseph Trotter, Scripps Research Institute, La Jolla, CA, USA).

Chick chorioallantoic membrane (CAM) assays (magnification ×12.5) with semi-quantitative angiogenesis scores at the bottom of each photograph (a) and bar charts with semi-quantitative angiogenesis scores of CAM duplicates, based on the number of bifurcations which are compared between the center and outside of the methylcellulose disks (white dotted line) (b). A-C: Sera at the designated time points; D: sera incubated in vitro with bevacizumab (500 μg/ml final concentration); E: supernatant of leukapheresis products; F: freezing medium including 10% DMSO and 33% RPMI cell culture medium.
Statistics. Kruskal-Wallis and Mann-Whitney U-tests were applied to assess statistical significance (P) of the difference between multiple and two groups, respectively. Fisher's r to Z test was performed for the calculation of significance P of correlations. P<0.05 was considered significant.
Results
Plasma VEGFR1 decreased after HDC, but significantly increased after ASCT. Plasma PGF, but not VEGF, increased after HDC. Median plasma VEGFR1 levels decreased by 13% after HDC, but increased by 33% immediately after ASCT. This increase was still apparent after hematological reconstitution 11 days after ASCT (Figure 1a and b). Changes in VEGF concentrations were not significant at any time point (Figure 1c). In contrast, PGF concentrations significantly increased after HDC, but did not further increase after ASCT (Figure 1d).
Sera of patients before HDC and ASCT stimulate angiogenesis, while sera after ASCT and leukapheresis products inhibit angiogenesis. Sera from patients undergoing HDC and ASCT induced increased capillary growth and numbers of bifurcations in the CAM assay before HDC, and after HDC/before ASCT. After ASCT, sera exhibited less ability to induce capillary growth and bifurcations, which was sustained through the time of hematological reconstitution 11 days later. Overall, the inhibitory effect of ASCT on angiogenesis in the CAM assay was comparable to the effect of bevacizumab, a humanized mouse monoclonal antibody targeting VEGF, when added to serum obtained before ASCT in a concentration in the upper range measured in human serum during clinical trials with weekly administration of bevacizumab at therapeutical doses of 10 mg/kg body weight. Supernatants of leukapheresis products had strong antiangiogenic effects, whereas sera of healthy volunteers, or freezing medium only, containing 10% DMSO, did not have any measurable effect on CAM angiogenesis (Figure 2a and b). Intra-individual increases in plasma VEGFR1 concentrations were negatively associated with angiogenesis, whereas decreases were positively associated. This correlation was statistically significant (Figure 3).
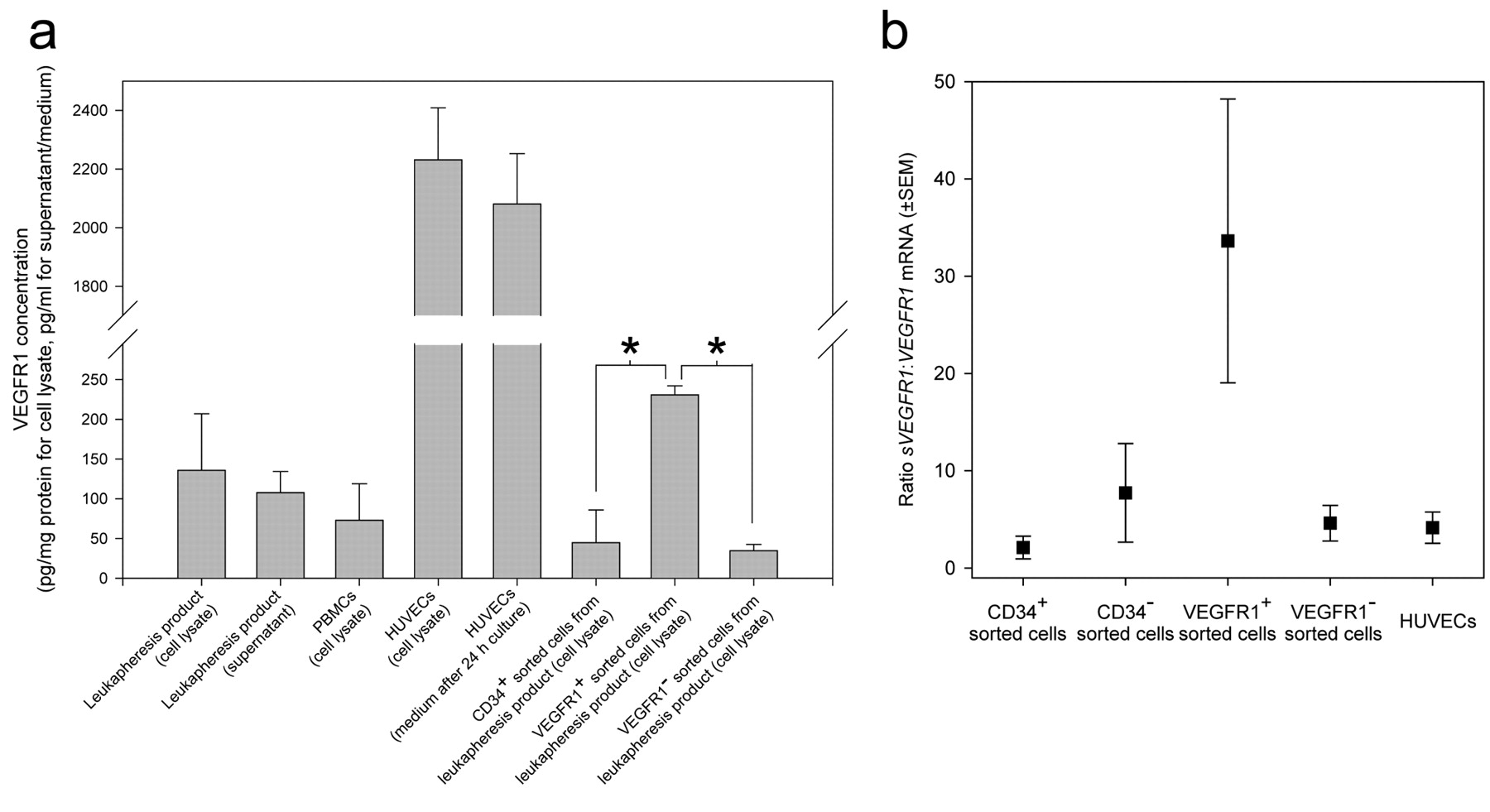
Leukapheresis products contain high concentrations of sVEGFR1 and VEGFR1 is expressed mostly on CD34− cells. To test the hypothesis that elevated levels of sVEGFR1 may be due to the sVEGFR1 present in the leukapheresis product, we analyzed sVEGFR1 concentrations in leukapheresis products. (s)VEGFR1 protein concentrations in supernatant (108 pg/ml) and cell lysate (136 pg/mg total protein) of leukapheresis products were comparable to values measured in PBMCs (73 pg/mg total protein) from other donors, but significantly lower than concentrations found in cell lysates and supernatants of human umbilical vein endothelial cells (HUVECs) in culture (2232 pg/mg total protein and 2081 pg/ml, respectively). Because CD34+ cells have been reported to express VEGFR1 (32), we sorted cells from leukapheresis products for CD34 and VEGFR1 expression by MACS. CD34+ cells expressed significantly less VEGFR1 than cells sorted for VEGFR1 expression (Figure 4a).

Correlation between percentage changes in plasma VEGFR1 concentrations and the pro-/antiangiogenic effects in the CAM of the corresponding sera from seven patients at four different time points (n=28; note that some dots overlap and may not be distinguished well from each other).
VEGFR1+ cells in leukapheresis products predominantly express mRNA encoding for sVEGFR1. RT-qPCR for the mRNAs of soluble sVEGFR1 and transmembrane VEGFR1 in cells from leukapheresis products demonstrated that the sVEGFR1:VEGFR1 mRNA ratio in the VEGFR1+ cell fraction was 34:1, compared to a ratio of between 2:1 and 8:1 in CD34+, CD34−, and VEGFR1− cells and cultured HUVECs (Figure 4b).
Neutrophilic granulocytes are the most abundant cell population in leukapheresis products and in the VEGFR-1+ cell fraction of leukapheresis products. To determine the cell type predominantly expressing VEGFR1 in leukapheresis products, we quantified leukocyte subpopulations in leukapheresis products and VEGFR1+, VEGFR1−, CD34+, and CD34− sorted cells from leukapheresis products using an automated blood cell analyzer. Neutrophils were the predominant cells within leukapheresis products. They were not only predominantly found in the VEGFR-1+ sorted cell fraction of leukapheresis products, but were also the only cell type to show a significant difference in number compared to the VEGFR-1− sorted cell fraction (Figure 5a). These results were confirmed by Giemsa staining of the cytospin of the sorted cells, where the VEGFR1+ population mainly consisted of damaged cells appearing as shadow nuclei of the decaying fragile neutrophilic granulocytes (not shown).
Flow cytometric analysis of leukapheresis products, CD34+, CD34−, VEGFR1+, and VEGFR1− sorted cells confirmed that CD34+ cells were mainly VEGFR1−, except for a small subpopulation, possibly representing endothelial progenitor cells (33). VEGFR1+ sorted cells unsurprisingly showed the most uniform VEGFR1+ peak. However, VEGFR1+ cells were also found in the VEGFR1− sorted cell fraction (Figure 5b-f).

Enzyme-linked immunosorbent assay of (s)VEGFR1 in whole leukapheresis products (apheresate) and in CD34 and VEGFR1 positively and negatively selected cells from apheresis products, respectively, compared to peripheral blood mononuclear cells (PBMC) and human umbilical vein endothelial cells (HUVEC) (a). *p<0.05. mRNA expression of the alternatively spliced variants of VEGFR1 (i.e. the soluble sVEGFR1 and the transmembrane VEGFR1) in VEGFR1 positively selected cells from apheresis products (b). n=4 for each experiment (mean±SEM).
The neutrophilic granulocyte-gated cells showed the same pattern of VEGFR1 expression as the whole-cell fraction positively or negatively sorted for VEGFR1, further corroborating the finding that VEGFR1+ cells in leukapheresis products were predominantly neutrophilic granulocytes (Figure 5g). Although the decrease in VEGFR1 plasma concentrations was paralleled by a decrease in neutrophilic granulocytes of 37% in the blood of patients after HDC, the changes in neutrophilic granulocytes at the different time points were not statistically significant (not shown).
Discussion
The effects of HDC and ASCT on angiogenesis have not yet been studied. In this study, we demonstrate that sVEGFR1, a strong endogenous inhibitor of VEGF activity, is found at high concentrations in leukapheresis products and in serum of patients after ASCT, and changes in VEGFR1 plasma concentrations correlate with the inhibitory effect on angiogenesis on the CAM. The cells most abundantly expressing VEGFR1 in leukapheresis products are neutrophils and in these cells, VEGFR1 mRNA splicing is shifted toward the expression of the soluble isoform.
While several endogenous inhibitors of angiogenesis (20) exist, VEGFR1 and its soluble isoform sVEGFR1 are among the strongest endogenous inhibitors of angiogenesis, acting directly on VEGF via sequestration of circulating VEGF and through a dominant negative effect by forming heterodimers with VEGFR1 and -2 (29). PGF has been shown to act as an amplifier of VEGF-driven angiogenesis (24). The changes in plasma VEGFR1 in our study, together with the finding that plasma VEGF is unchanged before and after HDC, and ASCT, respectively, and a lack of PGF increase after ASCT are consistent with a proangiogenic pattern after HDC, and an antiangiogenic pattern after ASCT. Tachi et al. observed an increase in VEGF serum levels over 3 months after bone-marrow mononuclear cell implantation for critical severe limb ischemia (14), while Sasaki et al. reported a decrease of proangiogenic factors in a patient with multiple myeloma following chemotherapy and peripheral ASCT (18). Interestingly, an increase in bone marrow endothelial colonies after HDC, and a significant reduction of their number after ASCT was observed in non-Hodgkin's lymphoma patients, corroborating an antiangiogenic response of ASCT also on a cellular level (16). A decrease in microvessel density after ASCT has been described in multiple myeloma (17) although this finding is not consistent (34). The source of sVEGFR1 in the blood of patients after ASCT for ischemic disease has been attributed to endothelial progenitor cells, which have been demonstrated to be increased after chemotherapy (33, 35) and ASCT (15). However, in our experience the CD34+ cell fraction (including endothelial progenitor cells) may be as low as 1-2% of all cells within the leukapheresis product, if no CD34+ enrichment procedure is performed, and one may wonder therefore if it is a major source of sVEGFR1.
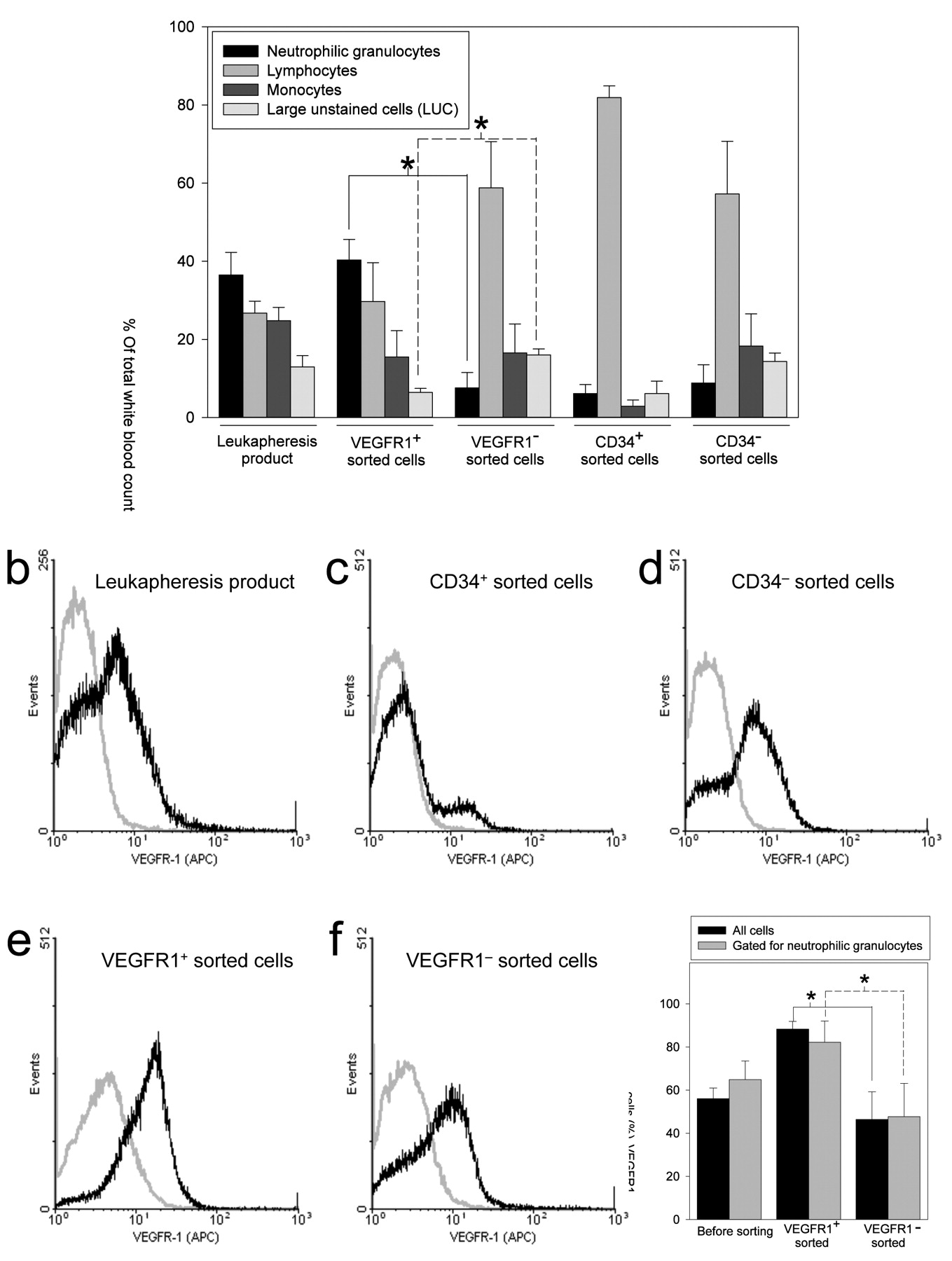
a: Automated blood cell analysis of leukocyte subfractions within leukapheresis products. n=4 (mean±SEM). *p<0.05. Flow cytometric analysis of a leukapheresis product (b), CD34+ (c), CD34− (d), VEGFR1+ (e), and VEGFR1− (f) magnetic activated sorted cells. g: Percentage of VEGFR-1+ cells in all cells vs. cells gated for neutrophilic granulocytes in leukapheresis products are shown. n=4 (mean±SEM). *p<0.05.
Stimulation of angiogenesis after HDC is not an unexpected finding, as recent studies reported changes consistent with an angiogenic switch after cytotoxic systemic chemotherapy (35, 36). There are only few clinical studies addressing the functional effects of ASCT on angiogenesis. Most studies have been performed in patients with ischemic digits or limbs due to connective tissue or vascular disease, and in patients with cardiac ischemic disease. Functional outcomes in these studies were measured by angiography, and variable rates of improved perfusion were reported, which can be considered a proangiogenic effect, or more precisely, vascular remodeling and arteriogenesis (6-8). However, the methods of autologous stem cell collection were considerably heterogenous in such studies, using only positively selected CD34+ cells from peripheral leukapheresis after recombinant human G-CSF with (13) or without (37) previous cyclophosphamide as a mobilization regimen, using mononuclear cells from bone marrow aspirate that contain only 0.5% CD34+ cells (10), or using recombinant human G-CSF-stimulated cells from direct bone marrow aspiration without further processing (11). Furthermore, there are no functional data on the potentially distinct effect of the co-infused non-cellular components of the leukapheresis product on angiogenesis. It is possible that cellular and non-cellular components of leukapheresis products have distinct and maybe even opposing effects on angiogenesis.
There are several explanations for the proangiogenic effect of serum from patients before HDC. All patients in our study received a cytotoxic chemotherapy and recombinant human G-CSF for stem cell mobilization shortly before ASCT, either as a previous chemotherapy cycle or as a separate regimen in patients with multiple myeloma. Chemotherapy itself could therefore be the cause of increased angiogenesis. However, given that all of our patients undergoing HDC/ASCT suffered from active cancer, the stimulatory effect of serum on CAM angiogenesis before HDC might also be explained, by tumors eliciting proangiogenic responses (35, 36). G-CSF has been shown to promote angiogenesis (38) via the release of VEGF from neutrophils (39). Interestingly, however, the administration of recombinant human G-CSF did not result in a proangiogenic response to serum in the CAM assay in the six out of seven patients receiving it from the day before ASCT until hematological reconstitution (40).
Leukapheresis products contained high concentrations of sVEGFR1. To our surprise, the cells expressing the highest concentrations of VEGFR1 in leukapheresis products were not CD34+ cells. Lysates of VEGFR1 positively selected cells expressed almost 5 times more VEGFR1 than CD34 positively selected cells, while the expression of VEGFR1 was not significantly different between CD34 positively and VEGFR1 negatively selected cells. VEGFR1 is expressed on many different cell types, including neutrophils (25-27, 32, 41-44). VEGF-dependent neutrophil migration has been demonstrated to be mediated by VEGFR1, in analogy to similar findings in monocytes (26, 42). For monocytes it has been shown that GM-CSF-induced sVEGFR1 expression inhibits angiogenesis (45). Given the abundance of neutrophils in leukapheresis products in our study, it seems likely that neutrophils are a major source of VEGFR1 after ASCT. In our study, neutrophils were the most abundant cell type in leukapheresis products, and only neutrophils showed a significant difference in number when leukapheresis products were positively and negatively selected for VEGFR1-expressing cells, further corroborating that overall, this cell type is the predominant cell type expressing VEGFR1.
However, the sole expression of transmembrane VEGFR1 by neutrophils does not yet explain the increase in soluble plasma sVEGFR. We speculated that a cell type, which predominantly expresses VEGFR1 would also be a major source of sVEGFR1, if the mRNA ratio of the differentially spliced isoforms sVEGFR1:VEGFR1 was not smaller than that of other VEGFR1-expressing cell types. In our study, the mRNA ratio of the differentially spliced isoforms sVEGFR1:VEGFR1 showed a shift in favor of the expression of sVEGFR1 in VEGFR1 positively selected cells, which we have demonstrated to consist mainly of neutrophils. Indeed, a reciprocal relationship of sVEGFR1 and VEGFR1 has been proposed for the modulation of responsiveness to VEGF in monocytes and neutrophils (26, 46).
Using flow cytometric analysis, we confirmed that CD34+ sorted cells are mainly VEGFR1−, with only a small population expressing VEGFR1. As CD34+ cells have been shown to express VEGFR1 (32), the small fraction of VEGFR1+ CD34 positively sorted cells was unexpected, but might represent endothelial progenitor cells (33).
While VEGFR1+ sorted cells naturally exhibited a strong and uniform VEGFR1+ peak, VEGFR1 negatively sorted cells also exhibited a VEGFR-1+ peak, albeit smaller. This can be technically explained by the possibility that VEGFR1 may be a weaker epitope for magnetic cell sorting, resulting in contamination of the VEGFR1− flow-through. When the cells were analyzed according to their typical scatter pattern (granulocyte gate), virtually all VEGFR1+ cells proved to be granulocytes. The results of cytologic and automated cell analysis confirmed the expected distribution, with >90% of the granulocyte population being neutrophilic granulocytes.
In conclusion, the efficiency of HDC followed by ASCT is believed to result from the fact that due to the re-infusion of bone marrow stem cells, the time to hematologic recovery can be shortened and chemotherapy can be applied at higher doses than hematologic toxicity resulting in prolonged aplasia and associated morbidity and mortality would normally allow. In this study, we provide evidence that ASCT after HDC can exert an antiangiogenic effect, possibly mediated via VEGFR1 derived from neutrophils in the leukapheresis product. Although a number of other factors, which have not yet been explored, could contribute to the antiangiogenic effects observed, our findings could have important implications for manipulative strategies in ASCT, e.g. CD34+ enrichment by purging. Consequently, application of neutrophil depletion techniques might result in a diminished beneficial antiangiogenic effect. Given the differences in the methodology of stem cell collection, a better understanding of the heterogeneity regarding the composition of leukapheresis products might explain the sometimes contradictory results in therapeutic angiogenesis and could help improving the outcome of ASCT in the treatment of hematologic and oncologic malignancy and ischemic disease.
Acknowledgements
We thank our clinical colleagues for collecting the required biological samples during the study, and Prof. Dr. Gabriele Schoedon, Dora Greuter, Patrick Spielmann, and Dr. Jyrki Eloranta for their skilful technical assistance with leukocyte isolation, leukapheresis, RT, and qPCR, respectively.
Footnotes
-
↵* These authors contributed equally to this work.
-
This article is freely accessible online.
- Received July 13, 2011.
- Revision received August 16, 2011.
- Accepted August 17, 2011.
- Copyright© 2011 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}