


Abstract
Background: Epigallocatechin-3-gallate (EGCG) is a polyphenol constituent present in green tea previously shown to inhibit cancer growth. However, studies on human ovarian cancer are limited. This study evaluated, the effects of EGCG as a potential anti-proliferative and pro-apoptotic agent in the human ovarian cancer line, SKOV-3. Materials and Methods: The MTS assay which measures metabolic activity of cells, bromodeoxyuridine (BrdU) incorporation assay, and flow cytometry were used for the cell proliferation studies and cell morphology, DNA fragmentation analysis and the TUNEL assay for apoptotic effects. Results: The EGCG treated SKOV-3 cells showed inhibition of cell viability and proliferation in a dose-dependent manner. Furthermore, EGCG-mediated SKOV-3 cell growth inhibition was associated with apoptotic changes as evident by cell cycle arrest and accumulation of cells in the apoptotic phase, cell morphological changes, DNA fragmentation and TUNEL-positive apoptotic cells. Conclusion: In SKOV-3 cells, EGCG inhibits cell proliferation via DNA synthesis reduction and induces apoptotic cell death via DNA damage, thus elucidating a novel, plausible mechanism of EGCG anti-tumorigenic property.
Tea derived from the leaves of the plant Camellia sinensis is the second most widely consumed beverage in the world, second only to water. Tea has been cultivated and consumed for thousands of years (1). Among all the teas consumed in the world, green tea remains the best studied for potential health benefits, including antitumor effects as evident from human epidemiological, in vitro cell culture and animal model studies (1-6). Most of the antitumor effects are mediated by polyphenols (catechins) which are naturally present in green tea.
Epigallocatechin-3-gallate (EGCG) (Figure 1) is the most abundant catechin in green tea and accounts for 50-80% of its polyphenol content, representing 200-300 mg in a brewed cup of green tea (3). EGCG offers several advantages as a putative anticancer agent given its ubiquitous distribution in nature, corresponding low cost and minimal toxicity. It is well documented that EGCG has antitumor effects on a number of carcinomas and human cancer cell lines (1, 3, 4, 7, 8). Hence, EGCG is a potential therapeutic agent for cancer therapy because of its anti-proliferative and prodifferentiation properties and ability to cause apoptosis. However, studies on human ovarian carcinomas are limited (9-14).
Ovarian cancer is the leading cause of death from all gynecological carcinomas (9). The prevalence and mortality of ovarian cancer has risen over the past century (15-16). The pathogenesis of this disease remains to be elucidated and novel therapeutic agents are constantly being explored. At present, the available experimental/human evidence on green tea intake and risk of ovarian cancer is limited. In a case−control and a 3-year prospective cohort study, involving 254 patients in China, Zhang et al. (9-10) concluded that increasing the consumption of green tea post-diagnosis enhanced ovarian cancer patients' survival and increasing the frequency and duration of tea consumption, especially of green tea, reduced the risk of developing ovarian cancer. Therefore, additional effort is needed to better elucidate the possible antitumor effects of EGCG on human ovarian cancer.
In this study, the anti-proliferative and pro-apoptotic potential of EGCG on cell survival, DNA synthesis, the cell cycle, cell morphology, DNA fragmentation and TUNEL-positive apoptotic cells was examined in the human ovarian cancer cell line, SKOV-3.
Materials and Methods
Cell culture. The SKOV-3 cells were obtained from the American Type Culture Collection (Rockville, MD, USA). Tissue culture media and fetal bovine serum (FBS) were purchased from Gibco (Carlsbad, CA, USA). EGCG and all the other reagents, except where indicated, were purchased from Sigma Chemical Company (St. Louis, MO, USA). The EGCG was dissolved in PBS and stored at −20°C. The SK0V-3 cell line was maintained in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% FBS, penicillin (100 units/ml), streptomycin (100 μg/ml) and 1mM glutamine. The cells were cultured at 37°C in a humidified atmosphere under 5% CO2. The cell culture medium was changed every 48-72 h. For all the experiments, unless otherwise stated, 1×103 cells were seeded in 96-well plates and grown to confluence for 24 h, prior to being treated with EGCG or the carrier solvent PBS. The final concentrations of EGCG used for all the experiments were prepared by diluting the stock with cell culture media. The cells were then treated with either the vehicle (PBS) or EGCG at various concentrations for 2 days.
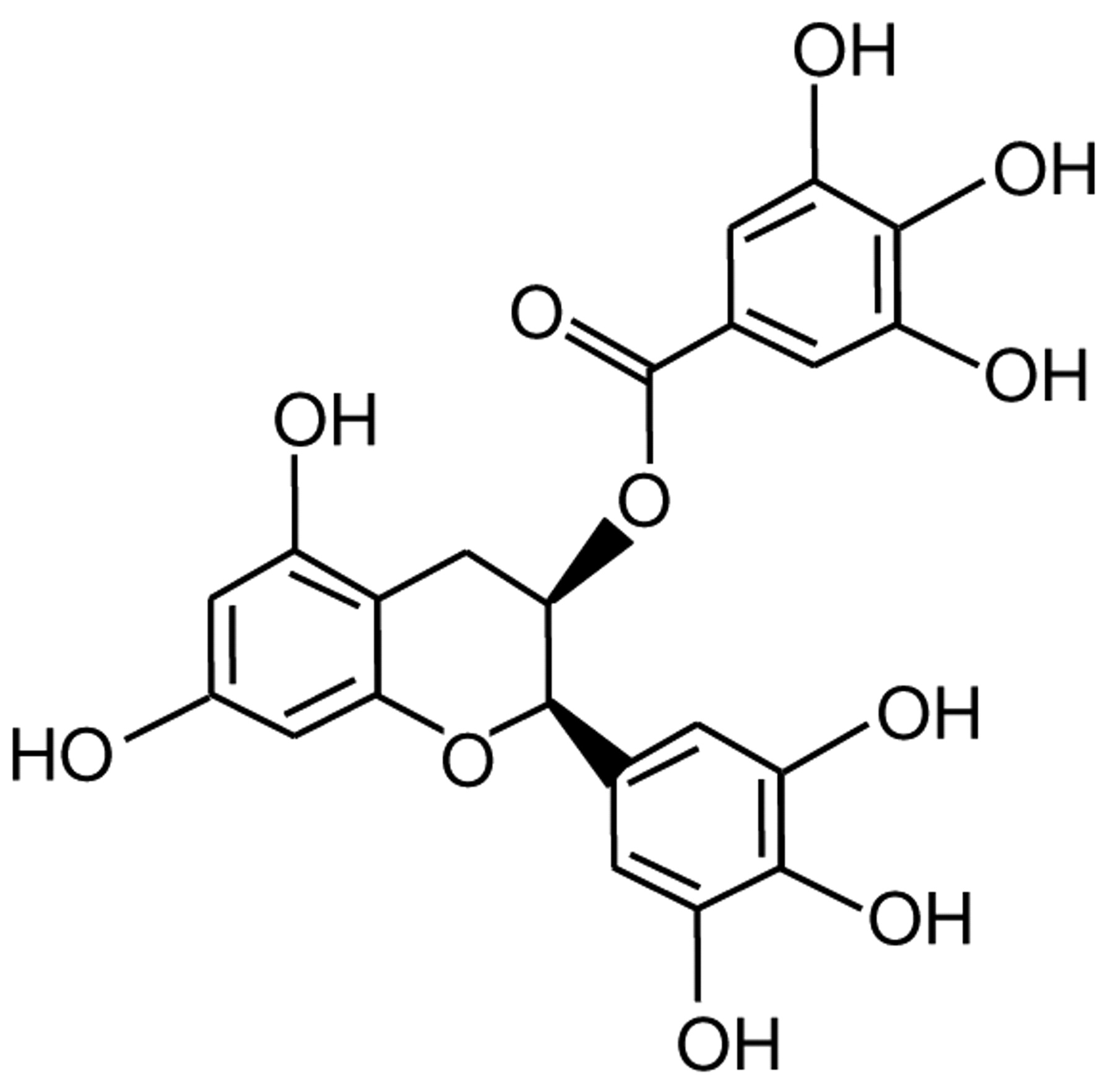
Structure of epigallocatechin-3-gallate (EGCG).
Cell viability. The MTS assay which measures the metabolic activity of cells was used to determine the survival of EGCG-treated cells compared to controls using a Cell Titre 96 Aqueous One Solution Proliferation Assay kit (Promega Corporation, Madison, WI, USA). Cell viability was determined spectrophotometrically by measuring the conversion of the MTS substrate to the formazon product which is directly proportional to the number of viable cells. In brief, MTS reagent (10 μl/well) was added to both the vehicle and EGCG treated cells and the OD at 490 nm was recorded after 4 h using a microplate reader. Cell viability was expressed as the percent of viable cells.
Cell proliferation. Bromodeoxyuridine (BrdU), a thyamine, DNA precursor analog incorporation assay kit (Roche Diagnostics, Indianapolis, IN, USA), which measures BrdU incorporation in place of thymidine during DNA synthesis before cell division, was used to determine the proliferation of EGCG-treated cells compared to controls. In brief, both the vehicle and EGCG-treated cells were labeled with BrdU (10 μM) for 6 h prior to incubation with anti-BrdU-peroxidase (POD) (10 μl/well) for 2 h. The immune complex was detected following the addition of trimethyl benzidine substrate and measured at 450 nm using an ELISA reader. The number of proliferating cells is represented by the level of BrdU incorporation which directly correlates to the color intensity and the absorbance values. Cell proliferation was expressed as the % BrdU incorporation.
Cell cycle analysis. Flow cytometry was used for cell cycle analysis. In brief, 3×105 cells were cultured in 6-well plates and treated with EGCG (10 μg/ml) or vehicle (PBS) for 2 days. Both the floating and adherent cells were collected and washed twice with PBS prior to staining using propidium iodide (PI). A FACScan flow cytometer and Modfit software (BD Biosciences, Mountain View, CA, USA) were used for data acquisition and analyis. Ten thousand events were analyzed for each sample.

Effect of EGCG on SKOV-3 cell viability. After 2 days of treatment, cell viability was determined by MTS assay. *p<0.01 Compared to control.

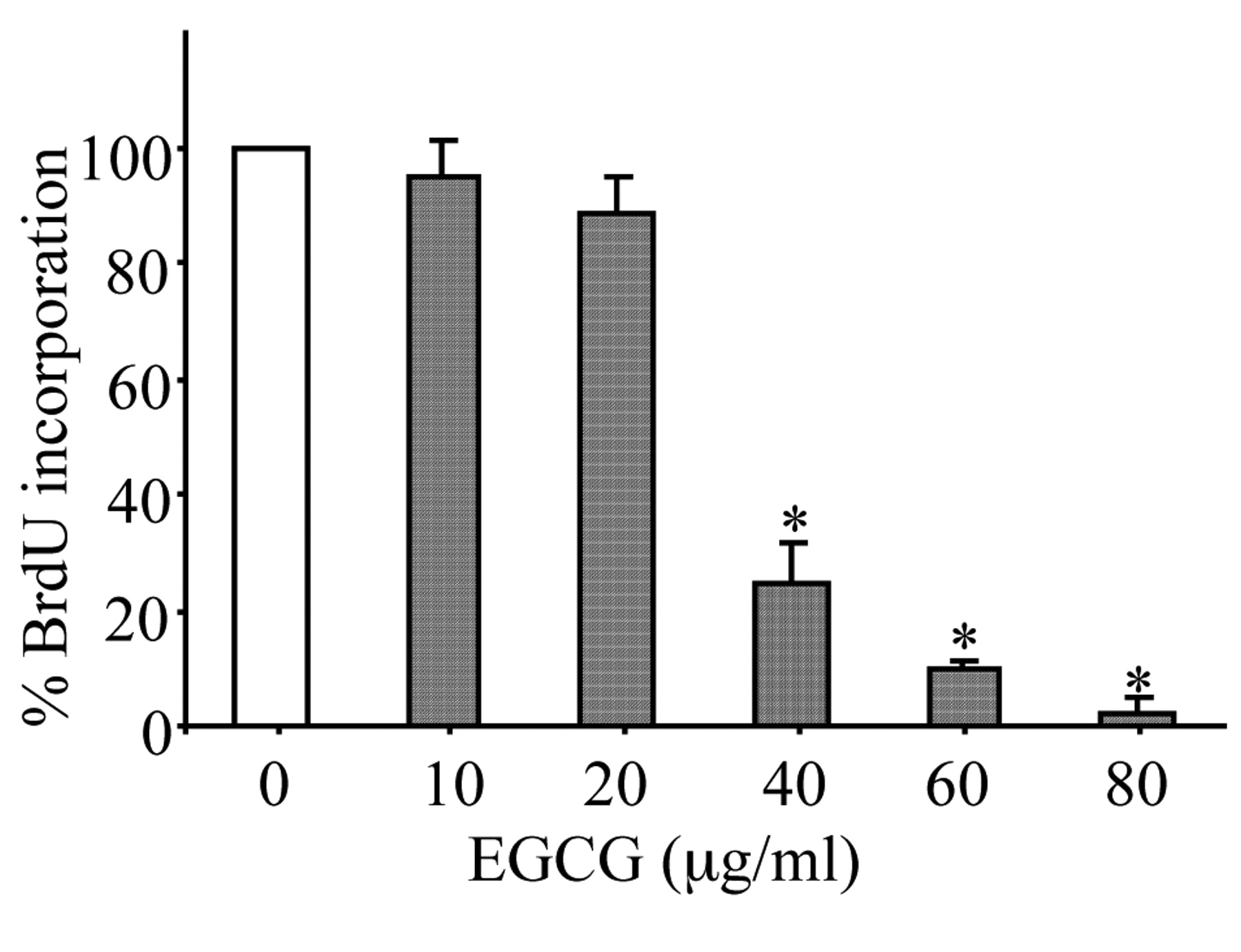
Effect of EGCG on SKOV-3 cell proliferation. After 2 days of treatment, DNA synthesis was measured by BrdU incorporation assay. *p<0.01 Compared to control.
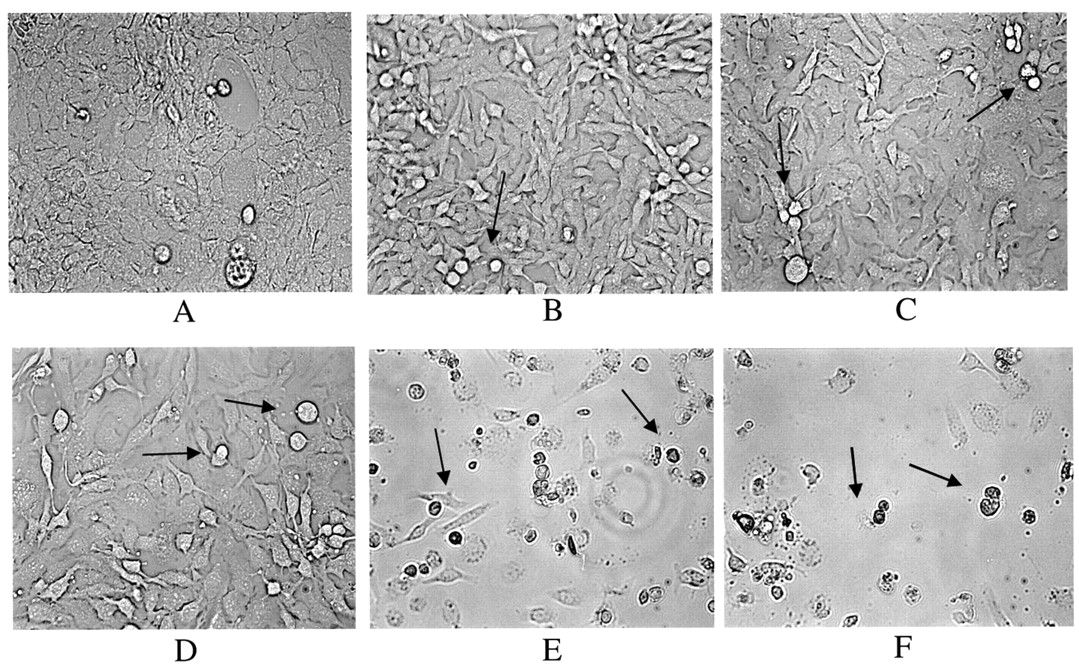
Cell morphology. The effect of EGCG on cell morphology was monitored as follows: the cells (5×106) were seeded in 48-well plates and treated with either vehicle (PBS) or EGCG (0-80 μg/ml) for 2 days. The cells were photographed (magnification ×10) with a Digital Camera attached to a phase-contrast light microscope (Nikon Eclipse TE2000-E; Tokyo, Japan).
DNA fragmentation analysis. An apoptotic DNA ladder kit (Roche Diagnostics) was used to determine nuclear DNA fragments in EGCG treated cells compared to controls. In brief, 1×106 cells were seeded in T-25 flasks and treated with vehicle (PBS) or EGCG (40 μg/ml for 2 days. Post treatment, both floating and adherent cells were collected, lysed in lysis buffer prior to DNA extraction using a glass fiber spin column. A 2% agarose gel was used to analyze the extracted DNA. DNA bands were visualized under UV light after ethidium bromide staining and photographed with a Chemi-Doc XRS instrument and Quantity One software (Bio-Rad Laboratories, Hercules, CA, USA).
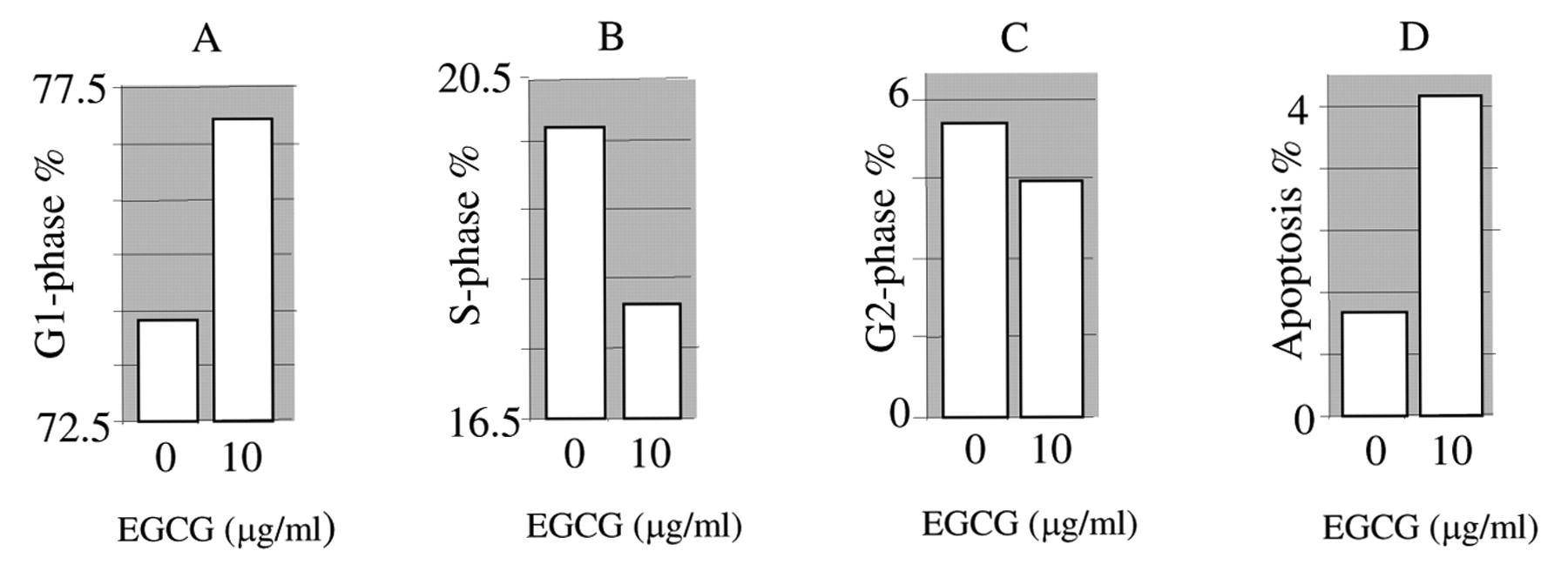
Effect of EGCG on cell cycle progression. After 2 days of treatment, the cell cycle phase distribution was determined by FACS analysis.
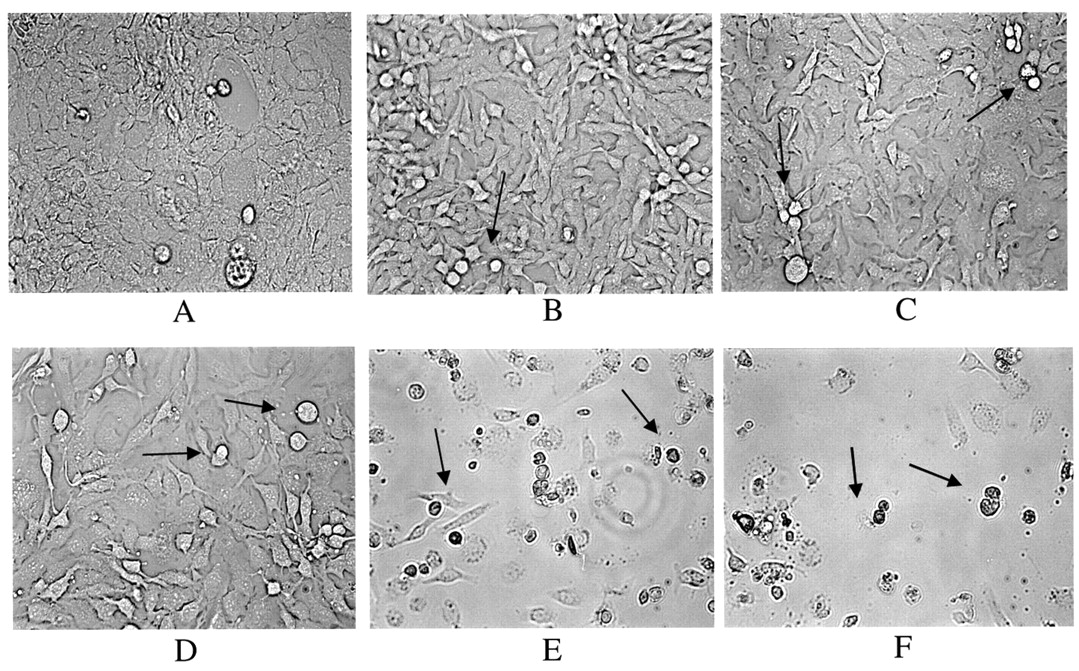
Effect of EGCG on cellular morphology of SKOV-3 cells. The cells were treated with EGCG for 2 days. A: Cells treated with vehicle (PBS) control; B-F: cells treated with EGCG at B: 10 μg/ml; C: 20 μg/ml; D: 40 μg/mL; E: 60 μg/ml; and F: 80 μg/ml EGCG. The arrows indicate cell death characterized by cellular shrinkage.
TUNEL assay. The TUNEL (terminal deoxynucleotidyltransferase-mediated deoxyuridine -5′-triphosphate (dUTP) nick-end labeling) assay used to determine apoptosis in EGCG treated cells compared to controls was a DeadEnd colorimetric apoptosis detection kit (Promega). In brief, 5×103 cells were seeded in a 48-well plate and treated with vehicle or EGCG (30 μg/ml) for 2 days. Post treatment, the cells were washed with PBS, fixed with 4% paraformaldehyde, permeabilized with Triton X-100 and labeled with fluorescein-12-dUTP using terminal deoxynucleotidyl transferase, followed by counterstaining with PI (5 μg/ml). The assay included negative controls (without terminal transferase). Apoptosis was detected by fluorescence microscopy (Nikon Eclipse TE2000-E fitted with a cooled charge-coupledevice camera at 10°C below ambient temperature with a digital resolution of 1.5 megapixels; Tokyo, Japan). The apoptotic cells were detected by fluorescein staining and the nuclei were detected by PI staining (magnification ×10).
Statistical analysis. The results are expressed as means±standard deviation (SD). Statistical significance was determined using Students t-test and one-way analysis of variance (ANOVA) where indicated. Statistical significance was reported if the p-value was greater than 0.05 (p<0.05).

DNA fragmentation analysis of SKOV-3 cells treated with EGCG (50 μg/m) for 2 days. Lane 1: Positive control (apoptotic U937 cells supplied with the DNA ladder kit); lane 2: DNA standard; lane 3: vehicle-treated cells; lane 4: EGCG-treated cells.
Results
Effect of EGCG on cell proliferation/survival. EGCG significantly inhibited survival in the SKOV-3 cells in a dose-dependent manner (p<0.01) (Figure 2). Furthermore, EGCG inhibited DNA synthesis in a dose-dependent manner as evident by the observed decrease in the incorporation of BrdU into DNA in the EGCG-treated cells compared to the controls (p<0.01) (Figure 3).

TUNNEL assay of SKOV-3 cells treated with EGCG (30 μg/ml) for 2 days. Representative photomicrographs of overlaid apoptotic stain (green) and nuclear stain (red). TUNEL-positive nuclei due to DNA fragmentation (bright yellow spots) are indicated by arrows. A: control; B: EGCG-treated cells.
Effect of EGCG on cell cycle. The FACS analysis demonstrated that treatment with EGCG increased the percentage of SKOV-3 cells in the G1-phase (Figure 4 A) with an associated decrease in the percentage of cells in S-phase (DNA synthesis) (Figure 4B) and G2-phase (Figure 4C) of the cell cycle when compared to the untreated controls. Furthermore, the percentage of cells in the apoptosis phase was increased when compared to the controls (Figure 4D).
Effect of EGCG on apoptosis. The EGCG-mediated SKOV-3 inhibition of cell proliferation/survival was associated with apoptotic changes. Morphological differences were observed between the EGCG-treated and the control cells (Figure 5A vs. B, C, D, E, F). At EGCG concentrations where inhibition of cell proliferation/survival was observed (10-80 μM), the cells were shrunken, rounded and detached from the cell culture substratum. These morphological changes were absent from the control cells (Figure 5A). Furthermore, DNA fragmentation/laddering (characteristic of apoptosis) was observed when the cells were treated with EGCG, indicating cell death by apoptosis in these cells while no DNA laddering was observed in the untreated cells (controls) (Figure 6). The TUNEL assay identified the nuclei by red PI staining and the green tunel positive (apoptotic) nuclei were identified by yellow spots in the PI and TUNEL overlay. Numerous yellow spots were seen in the EGCG-treated cells, when compared to control cells (Figure 7).
Discussion
Earlier studies (11, 12) and the present study showed that EGCG inhibited the viability of SKOV-3 cells. The present study had the advantage that the MTS assay was used, which measures the mitochondrial activity of cells. EGCG-treated cells lost the ability to provide and maintain energy for metabolic activity and survival indicating that EGCG caused cellular damage. DNA replication occurs before cell division/doubling, thus the measurement of DNA synthesis, is an attractive marker for cell proliferation. A decrease in BrdU-labeled DNA was noted in the EGCG treated cells, suggesting that EGCG is an inhibitor of DNA synthesis and cell proliferation in SKOV-3 cells.
Furthermore, the percentage of EGCG-treated cells in the apoptotic phase was significantly increased as compared to the untreated cells (Figure 4D), suggesting a block in cell cycle progression leading to programmed cell death. This was in agreement with previous studies which established that apoptosis is the likely mechanism of action of tea polyphenols for inhibiting cell growth in various carcinomas (1, 3, 5, 7). Thus, the activation of apoptosis is believed to play a critical role in both the chemo prevention and treatment of human carcinomas. Morphological changes associated with apoptosis were also observed in the EGCG treated cells. The biochemical hallmark of apoptosis is genomic DNA fragmentation, an irreversible event that commits the cell to die. The nuclear endonuclease enzyme cleaves DNA resulting in DNA fragments. The EGCG treatment caused DNA fragmentation (Figure 6), further confirmed by the TUNEL assay, labeling the DNA fragments and indicating that EGCG induced apoptosis in the SKOV-3 cells by causing DNA damage.
In a recent study, the combination of ECGC with lysine, proline, arginine and ascorbic acid was shown to inhibit critical steps in ovarian cancer development and spread, such as matrix metalloproteinase-2 secretion and invasion (14). In prostate cancer DU-145 cells, the combination of EGCG and the nonsteroidal anti-inflammatory drug (NSAID), ibuprofen had a synergistic effect on apoptosis (17). The synergistic effects of EGCG with NSAIDS on ovarian cancer are not known. In ovarian cancer, the postulated mechanisms of EGCG anti-tumorigenic effects include decreasing the levels of circulating estrogen and endothelin. Recent limited data have linked green tea/EGCG intake to lower circulating estrogen levels and lower ovarian cancer risk in human epidemiological and animal studies (18), however more studies are needed to further explore this relationship. Furthermore, endothelin-1 (ET-1) and its selective receptor ETA (ETAR) are overexpressed in primary and metastatic ovarian carcinoma, when compared to normal ovarian tissue. Binding of ET-1 to ETAR activates the receptor and triggers tumor growth, neoangiogenesis, invasiveness and apoptosis protection. Pretreatment of ovarian cancer cell lines HEY and OVCA 433 with green tea and EGCG inhibited ET-1 and ETAR mRNA expression and downstream signaling pathways resulting in the reduction of cell proliferation, angiogenesis and invasiveness (4, 13). EGCG appears to be a multifunctional agent recognized for antitumor, anti-angiogenic, anti-metastatic and anti-invasive properties.
In summary, the anti-proliferative and proapoptotic properties of EGCG in the human ovarian cancer cell line, SKOV-3, were supported by the observed reduction in cell survival and DNA synthesis, as well as cell cycle arrest and the accumulation of cells in the apoptotic phase caused by DNA damage.
Acknowledgements
We would like to thank NIH COBRE Grant 1-P20RR018728 for providing instrumentation support.
- Received August 24, 2010.
- Revision received May 14, 2010.
- Accepted May 14, 2010.
- Copyright© 2010 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}