


Abstract
It has been reported that curcumin inhibited various types of cancer cells in vitro and in vivo. However, mechanisms of curcumin-inhibited cell growth and -induced apoptosis in human non-small cell lung cancer cells (NCI-H460) still remain unclear. In this study, NCI-H460 cells were treated with curcumin to determine its anticancer activity. Different concentrations of curcumin were used for different durations in NCI-H460 cells and the subsequent changes in the cell morphology, viability, cell cycle, mRNA and protein expressions were determined. Curcumin induced apoptotic morphologic changes in NCI-H460 cells in a dose-dependent manner. After curcumin treatment, BAX and BAD were up-regulated, BCL-2, BCL-XL and XIAP were down-regulated. In addition, reactive oxygen species (ROS), intracellular Ca2+ and endoplasmic reticulum (ER) stress were increased in NCI-H460 cells after exposure to curcumin. These signals led to a loss of mitochondrial membrane potential (ΔΨm) and culminated in caspase-3 activation. Curcumin-induced apoptosis was also stimulated through the FAS/caspase-8 (extrinsic) pathway and ER stress proteins, growth arrest- and DNA damage-inducible gene 153 (GADD153) and glucose-regulated protein 78 (GRP78) were activated in the NCI-H460 cells. Apoptotic cell death induced by curcumin was significantly reversed by pretreatment with ROS scavenger or caspase-8 inhibitor. Furthermore, the NCI-H460 cells tended to be arrested at the G2/M cell cycle stage after curcumin treatment and down-regulation of cyclin-dependent kinase 1 (CDK1) may be involved. In summary, curcumin exerts its anticancer effects on lung cancer NCI-H460 cells through apoptosis or cell cycle arrest.
- Curcumin
- apoptosis
- G2/M arrest
- caspase cascade
- endoplasmic reticulum stress
- lung neoplasms
- herbal medicine
Lung cancer is the most common cause of cancer deaths in the United States and other parts of the world (1). In spite of advances in chemotherapy, the prognosis of lung cancer remains poor. Natural products were the main source of health care products in ancient times. In modern medicine, they are still major sources of new drug development (2).
Curcumin (diferuloylmethane), a phenolic compound isolated from the plant Curcuma longa, has been used in traditional medicines of China and India for thousands of years. Curcumin exhibits anticancer effects in various types of cancer cell line in vitro, including these of hematologic malignancies, head and neck, genitourinary, gastrointestinal, breast, ovarian and neurologic cancer, melanoma and sarcoma (3). These anticancer activities are partly attributed to its effects on many molecular targets involved in cell cycle, apoptosis, transformation, proliferation, survival, invasion, angiogenesis and metastasis of tumor cells (4). In animal models, curcumin given orally, intravenously, intraperitoneally or by gavage has been reported to have anticancer effects in different types of cancer (5). Many clinical trials on curucmin with cancer patients are ongoing (6). The safety of curcumin was demonstrated in a phase I study, which showed oral ingestion of curcumin up to 8000 mg/day for 3 months did not produce any treatment-related toxicity (7). Some biologic activities were observed in a phase II trial on pancreatic cancer patients taking curcumin daily (8).
Significant amounts of curcumin were found in lung tissue 2 minutes after injection with radiolabelled curcumin into tail veins of imprinting control region (ICR) mice (9). This pharmacokinetic information has made curcumin treatment for lung cancer feasible. Curcumin inhibits activator protein-1 (AP-1) and mediastinal lymph node metastasis of Lewis lung carcinoma cells (10) and induces cell death in A549 cells (human lung adenocarcinoma cell line) through apoptosis (11-12). Human lung adenocarcinoma cells (CL1-5) treated with curcumin showed a significant reduction in tumor migration, invasion and metastatic ability (13). Curcumin reduced N-bis(2-hydroxypropyl) nitrosamine-induced lung tumorigenesis in BALB/c mice in vivo (14). The NCI-H460 cell line is derived from human large cell lung cancer, which is one of the major types of non-small cell lung carcinoma. Direct cytotoxic effects of curcumin on NCI-H460 cells have never been reported and this is the main purpose of this study.
Materials and Methods
Chemicals and reagents. Curcumin, propidium iodide (PI), Triton X-100 and N-acetylcysteine (NAC) were obtained from Sigma-Aldrich Chemical Co. (St. Louis, MO, USA). Culture medium RPMI-1640, fetal bovine serum (FBS), penicillin-streptomycin and L-glutamine were purchased from GIBCO BRL (Invitrogen, Grand Island, NY, USA). 2,7-Dichlorodihydrofluorescein diacetate (DCFH-DA), 3,3′-dihexyloxacarbocyanine iodide (DiOC6) and N-[4-[6-[(acetyloxy) methoxy]-2,7-dichloro-3-oxo-3H-xanthen-9-yl]-2-[2-[2-[bis[2[(acetyloxy)methoxy]-2-oxyethyl] amino]-5-methyl-phenoxy] ethoxy]phenyl-N-[2-[(acetyloxy) methoxy]-2-oxyethyl]-, (acetyloxy) methyl ester (Fluo-3/AM) were purchased from Molecular Probes (Invitrogen, Eugene, OR, USA). Caspase-8 inhibitor (Z-IETD-FMK) was from R&D systems (Minneapolis, MN, USA).
Cell culture. Human lung carcinoma cell line NCI-H460 was obtained from the Food Industry Research and Development Institute (Hsinchu, Taiwan, R.O.C.). It was maintained in RPMI-1640 medium plus 10% FBS, 2 mM L-glutamine, 100 Units/ml penicillin and 100 μg/ml streptomycin. The cells were kept in an incubator at 37°C under 5% CO2 and 95% air. Stocked cells were plated to a culture flask or a 6-well plate at a density of 2-5×105 cells per ml prior to each experiment.
Morphologic changes and cell viability determinations. NCI-H460 cells were plated onto 12-well plates at a density of 2×105 cells/well and curcumin was added at final concentrations of 0, 5, 10, 20, 30, 40 or 50 μM. In control wells, only dimethyl sulfoxide (DMSO; solvent) was added. The cells were exposed for 24 h and a phase-contrast microscope was used to observe morphologic changes in examined cells at the end of this time. Cell viability was estimated through flow cytometric methods described elsewhere (15-16).
Cell cycle and apoptotic cell determinations. Approximately 2×105 cells/well of NCI-H460 cells in 12-well plates were incubated with 5, 10, 20, 30 or 40 μM of curcumin for 24 h. The cells were harvested by centrifugation, washed with PBS and fixed in 70% ethanol at −20°C overnight. The cells were then re-suspended in PBS containing 40 μg/ml of PI, 0.1 mg/ml RNase and 0.1% Triton X-100 in a dark room for 30 min and were subsequently analyzed by a flow cytometer (FACSCalibur, Becton-Dickinson, San Jose, CA, USA) (17, 18). The cell cycle distribution and sub-G1 groups (apoptosis) were calculated and analyzed by CellQuest (Becton-Dickinson) and ModFit LT software (Verity Software House Inc., USA).
Caspase-3, -8 and -9 activity assays. NCI-H460 cells were plated onto 12-well plates at a density of 2×105 cells/well with 25 μM curcumin. The cells were then incubated for different periods of time and harvested, washed and re-suspended in 50 μl of 10 μM substrate solution (PhiPhiLux and CaspaLux kit, OncoImmunin, Inc. Gaithersburg, MD, USA) before being incubated at 37°C for 60 min. The cells were washed again by PBS and were analyzed by flow cytometry (16, 18, 19).
Flow cytometric detection of reactive oxygen species (ROS), mitochondrial membrane potential (ΔΨm) and intracellular Ca2+ levels. Approximately 2×105 cells/well of NCI-H460 cells into 12-well plates were incubated with 25 μM curcumin for 3, 6, 12 and 24 h. The cells were harvested and washed twice, re-suspended in 500 μl of DCFH-DA (10 μM) for ROS, DiOC6 (4 μmol/l) for ΔΨm and Fluo-3/AM (3 μg/ml) for intracellular Ca2+. The cells were incubated with these specific fluorochromes at 37°C for 30 min and the differential expressions were analyzed by flow cytometry (20-22).
Effects on viability after pre-treatment with caspase-8 inhibitor before curcumin treatment. Cultured NCI-H460 cells with or without pre-treatment with 20 μM of caspase-8 inhibitor (Z-IETD-FMK) for 3 h prior to the addition of 25 μM of curcumin were then analyzed for caspase-8 activity and cell viability. These methods have described previously (16, 23).
Effect of ROS scavenger (NAC) on curcumin-treated NCI-H460 cells. Approximately 2×105 cells/well of NCI-H460 cells were plated into 12-well plates with or without pre-treatment with 15 mM of NAC for 3 h before the addition of 25 μM of curcumin and the levels of cell viability and ΔΨm were determined. The methods of determination were described previously (20-21).
Primers used in real-time PCR in this study. The DNA sequence was evaluated using Primer Express software.
Immunostaining and confocal laser microscopy examinations for mitochondrial release and ER stress related protein levels. NCI-H460 cells (5×104 cells/well) plated onto 4-well chamber slides were treated with 25 μM curcumin for 24 h. The cells were fixed in 4% formaldehyde for 15 min, permeabilized with 0.2% Triton-X 100 in PBS for 1 h with blocking of non-specific binding sites using 2% bovine serum albumin (BSA). The slides were then incubated with anti-cytochrome c, -endonuclease G (Endo G), -GADD153 and -GRP78 primary antibodies (1:100 dilution; Santa Cruz Inc., CA, USA) overnight and exposed to secondary antibodies (FITC-conjugated goat anti-mouse IgG at 1:100 dilution; Santa Cruz Inc.) at 37°C for 2 h. Mitochondria and nuclei were then counterstained with MitoTracker® Red CMXRos (Molecular Probes/Invitrogen) and PI. Photomicrographs were obtained using a Leica TCS SP2 Confocal Spectral Microscope (24-25).
Effects of curcumin on apoptosis-associated proteins by Western blotting assay. Total proteins were collected from the NCI-H460 cells after treatment with 25 μM of curcumin for 0, 6, 12, 24 and 48 h. The amounts of proteins (B cell lymphoma-2 (BCL-2), Bcl extra long (BCL-XL), BAX, BAD, X-linked inhibitor of apoptosis protein (XIAP), FAS/CD95, cyclin A, cyclin D, cyclin E, cyclin dependent kinase 2 (CDK2), CDK4, CDK6, CDK1, cyclin B, CDC25c) were determined by sodium dodecylsulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and immunoblotting as previously described (26-28).
Real-time PCR assay for mRNA levels of relative apoptosis gene expressions. Total RNA was extracted from the NCI-H460 cells after treatment with 25 μM curcumin for 24 and 48 h, using Qiagen RNeasy Mini Kit (Qiagen, Valencia, CA, USA) as described previously (27). The RNA samples were reverse-transcribed according to the standard protocol of the supplier (Applied Biosystems, Foster City, CA, USA). Quantitative PCR was performed using 2× SYBR Green PCR Master Mix (Applied Biosystems) and 200 nM of forward (F) and reverse (R) primers (Table I). Each assay was run on an Applied Biosystems 7300 Real-Time PCR system in triplicate and expression fold-changes were derived using the comparative CT method (12, 29).
Statistical analysis. All data are expressed as the mean±standard deviation (S.D.). Student's t-test was used to compare data of different groups. A p-value less than 0.05 was considered significant.

Curcumin affected the morphology and percentage of viable NCI-H460 cells. The cells were treated with or without various doses of curcumin for 24 h. The cells were examined and photographed (×200) under phase-contrast microscopy (A) and the percentage of viable cells (B) was determined as described in the Materials and Methods. The asterisks indicate significant differences from the control at *p<0.05 and ***p<0.001 by Student's t-test.
Results
Morphologic changes and decreased cell viability after curcumin treatment of NCI-H460 cells. After curcumin treatment for 24 h, the cells underwent significant morphologic changes, including membrane bleeding and apoptotic bodies were observed in NCI-H460 cells which were examined and photographed under a microscope as shown in Figure 1A. In addition, the viability of NCI-H460 cells decreased in a concentration-dependent manner after treatment with curcumin for 24 h (Figure 1B).
Cell cycle arrest and apoptosis of NCI-H460 cells after treatment with curcumin. The cell cycle distribution of NCI-H460 cells after treatment with 0, 5 and 10 μM of curcumin for 24 h is depicted in Figure 2A. The cell cycle tended to be arrested at the G2/M stage after exposure to curcumin. The sub-G1 peaks, indicating the proportions of apoptosis, increased in a dose-dependent manner when the concentration of curcumin added increased (Figure 2B).

Curcumin promoted G2/M phase arrest and caused apoptosis of NCI-H460 cells. The cells were treated with different concentrations of curcumin for 24 h. The distribution of cell cycle and sub-G1 (apoptosis) peaks were examined by flow cytometric assay. A: Bar chart showing the cell cycle distribution; B: apoptosis (%). The asterisks indicate significant differences from the control at *p<0.05 and ***p<0.001 by Student's t-test.
Curcumin stimulated the caspase-3, -8 and -9 activity of NCI-H460 cells. After treatment with curcumin for different durations, the NCI-H460 cells expressed increased caspase-3 activity. The caspase-3 activity reached its maximum when the duration of treatment was 12 h. The caspase-8 and -9 activities of NCI-H460 cells were also increased after 25 μM of curcumin treatment (Figure 3).
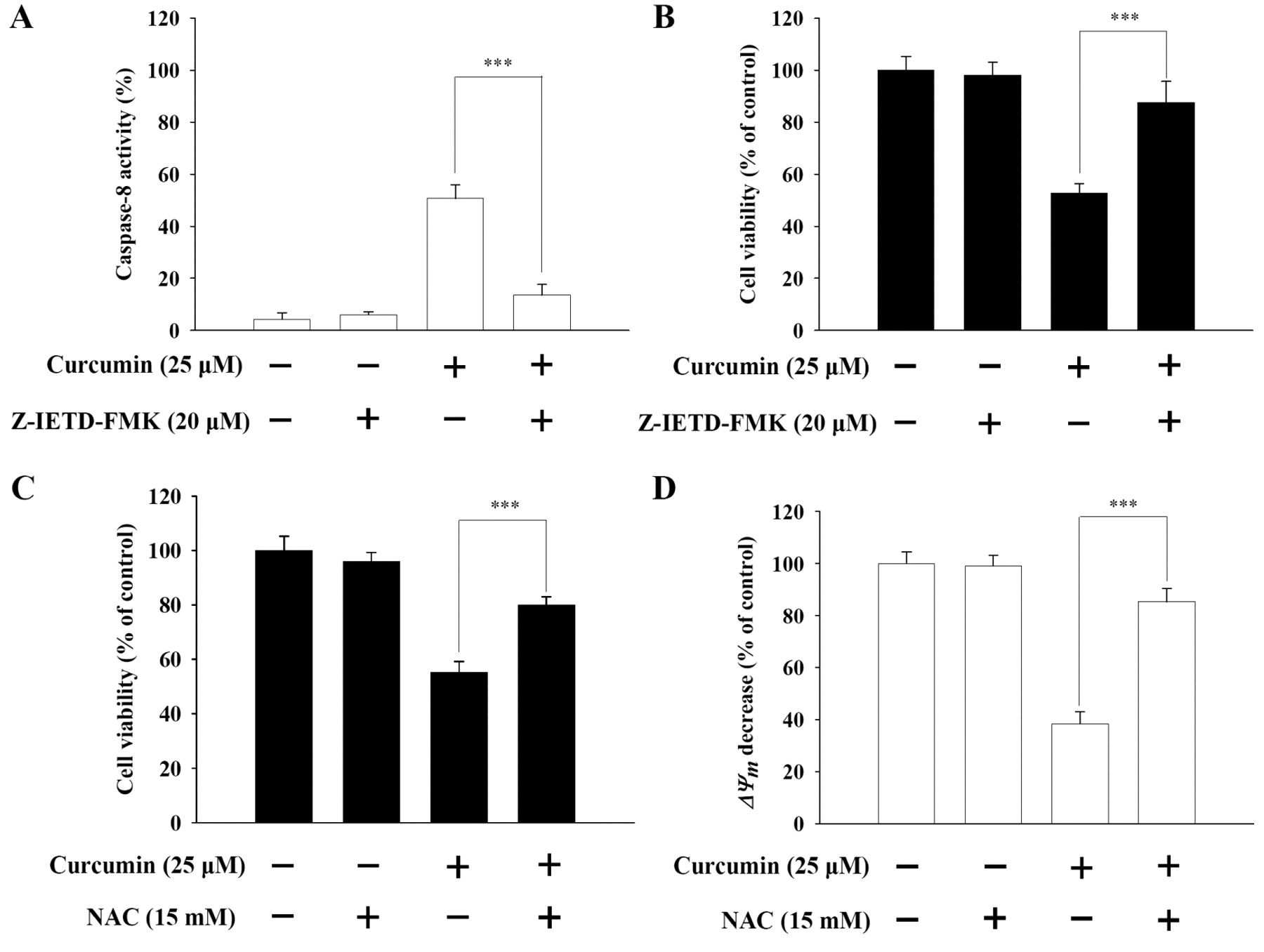
Effects of caspase-8 inhibitor on curcumin-treated NCI-H460 cells. Caspase-8 inhibitor (Z-IETD-FMK) significantly suppressed the increased caspase-8 activity after exposure to curcumin (Figure 4A). Curcumin-induced apoptotic cell death in NCI-H460 cells was also significantly reversed by prior to Z-IETD-FMK (Figure 4B).
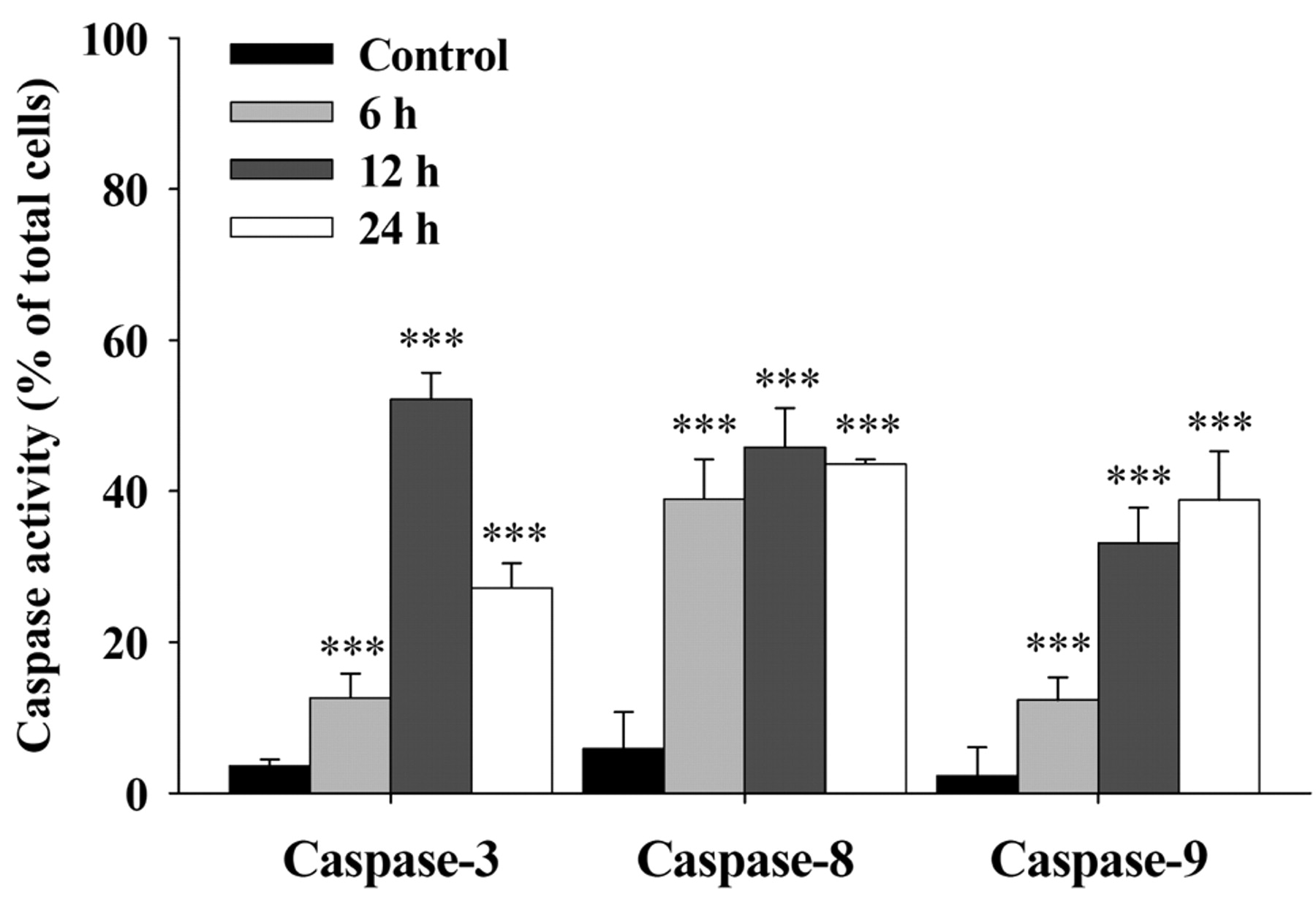
Curcumin stimulated the activities of caspase-3, -8 and -9 in NCI-H460 cells. The cells were treated with 25 μM curcumin for 0, 6, 12 and 24 h then the activities of caspase-3, -8 and -9 were determined by flow cytomtric assay as described in the Materials and Methods. The asterisks indicate significant differences from the control at p<0.001 by Student's t-test.
Flow cytometric analysis of ROS, intracellular Ca2+ and ΔΨm levels in NCI-H460 cells with or without curcumin treatment.
Effects of curcumin on ROS production, ΔΨm and intracellular Ca2+ levels in NCI-H460 cells. After curcumin treatment for different time intervals, the ROS and intracellular Ca2+ increased significantly as compared with the control group. In addition, there was significant loss of ΔΨm after treatment with 25 μM of curcumin (Table II).
Effects of ROS scavenger on curcumin-treated NCI-H460 cells. In addition to reducing ROS production, NAC (ROS scavenger) significantly reversed the curcumin-induced apoptotic cell death in NCI-H460 cells (Figure 4C). Moreover, in cells pretreated with NAC before treatment with curcumin, there was a significant loss of ΔΨm (Figure 4D).
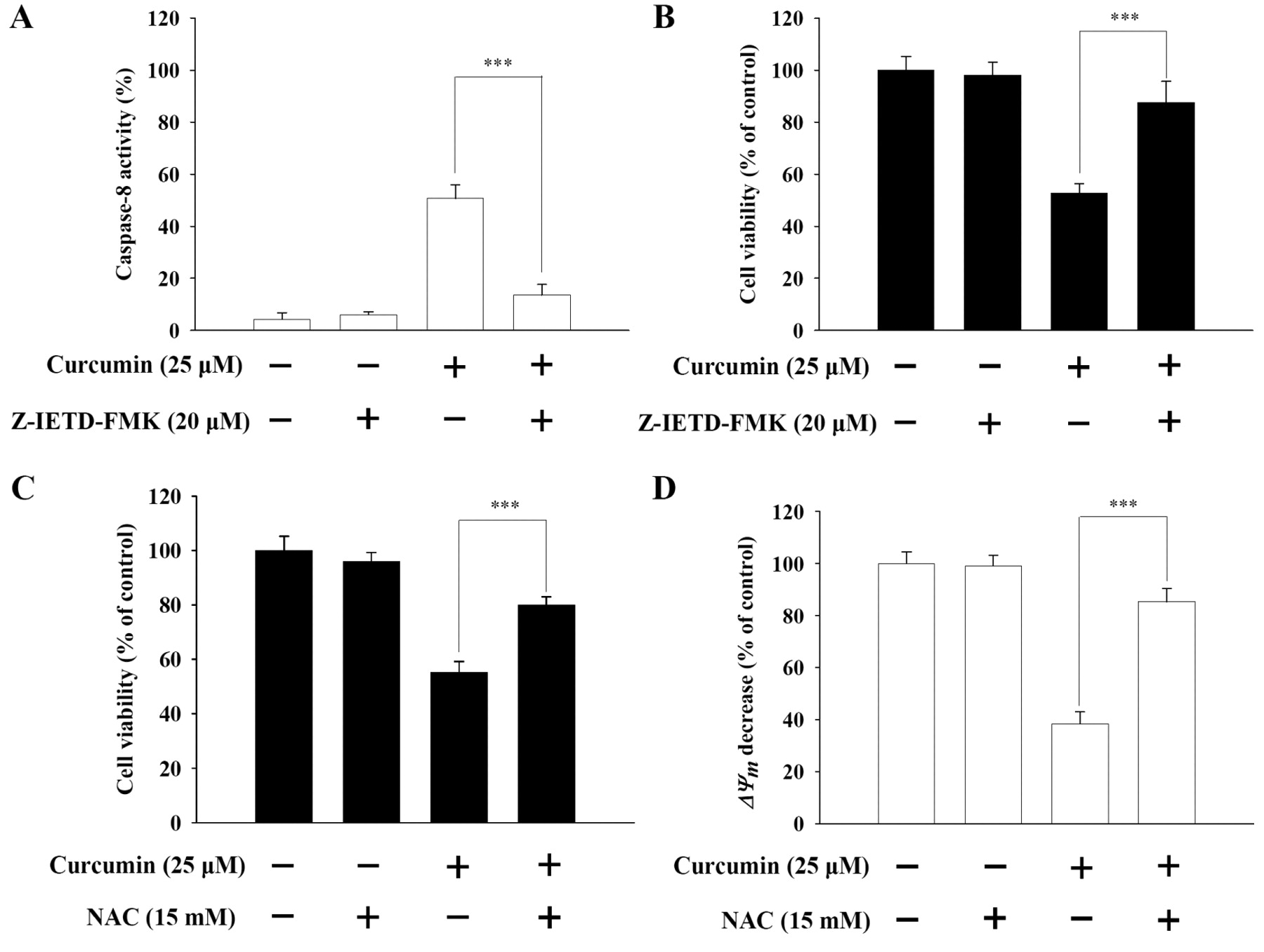
Caspase-8 inhibitor (Z-IETD-FMK) and ROS scavenger (NAC) affected curcumin-induced caspase-8 activity, the percentage of viable cells and loss of ΔΨm in NCI-H460 cells. The cells were pre-treated with Z-IETD-FMK or NAC, exposed to curcumin for 24 h and then harvested for determining the caspase-8 activity (A), the percentage of viable cells (B and C) and loss of ΔΨm (D) as described in the Materials and Methods. The asterisks indicate significant differences from the control at p<0.001 by Student's t-test.
Immunofluorescence staining for the release of mitochondrial and ER stress-associated proteins. The results indicated that curcumin promoted the release of cytochrome c and Endo G from mitochondria, with their translocation to cytosol and nuclei when compared to the untreated control. ER stress marker proteins (GADD153 and GRP78) were also expressed after exposure to curcumin in NCI-H460 cells compared to control. All results are shown in Figure 5.
Effects of curcumin on apoptosis associated proteins in NCI-H460 cells. Western blotting analysis was used for determining proteins involved in apoptosis and cell cycle regulation. After 25 μM of curcumin treatment, the anti-apoptotic proteins: BCL-2, BCL-XL and XIAP were down-regulated in a time-dependent manner; the pro-apoptotic proteins: BAX, BAD and FAS/CD95 were up-regulated (Figure 6A). Cell cycle regulatory proteins: cyclin D and E were initially up-regulated followed by down-regulation. CDK1, CDK2, CDK4 and CDK6 were down-regulated after curcumin treatment (Figure 6B).
Curcumin promoted caspase-8 and Endo G mRNA gene expressions in NCI-H460 cells. The apoptotic cell death-associated gene expressions of NCI-H460 cells were assessed by real-time PCR method after treatment with 25 μM curcumin for 0, 24 and 48 h. The expression of caspase-8 mRNA increased after 24 and 48 h of curcumin treatment in a time-dependent manner. Endo G mRNA increased only after 24 h of curcumin treatment when compared with the untreated control (Figure 6C).
Discussion
In this study, we demonstrated that curcumin was cytotoxic towards human lung cancer NCI-H460 cells in a dose-dependent manner. When the concentration of curcumin was escalated to over 20 μM, the viability of NCI-H460 cells decreased precipitously. If 30 μM of curcumin was added, nearly 95% of NCI-H460 cells died (Figure 1B). The cell death coincided with the emergence of significant apoptosis, represented by the sub-G1 peak base on the flow cytometric results (Figure 2B). It was reported that 40-50 μM of curcumin was required to induce 50% of human lung cancer A549 cells to die (11, 12). NCI-H460 cells, therefore, are more sensitive to the cytotoxic effect of curcumin than A549 cells. Because the bioavailability of curcumin is low, the experimental concentrations used in cell cultures are hard to achieve in vivo. Intravenous injection of 10 mg/kg of curcumin into rats produced a maximal serum level of 0.36±0.05 μg/ml (0.98±0.14 μM). The level achieved was even lower when a 50-fold dose of curcumin was given orally to these rats (30). In human beings, one hour after oral ingestion of 4-8 g of curcumin, the plasma concentration of curcumin was only 0.41-1.75 μM (7). Despite the low bioavailability, therapeutic efficacy of curcumin against various diseases has been documented. Such a disparity between bioavailability and efficacy has not been well explained (6). In addition, there are many ways to improve the bioavailability of curcumin, for example using an adjuvant such as piperine or liposomal curcumin (31).

Curcumin promoted the release of cytochrome c and Endo G from mitochondria in NCI-H460 cells. ER stress hallmark proteins (GADD153 and GRP78) were also expressed in examined cells as compared with untreated control cells. The cells were exposed to 25 μM of curcumin for 24 h, then were stained and examined and photographed by confocal microscopy as described in the Materials and Methods. A/E: Cytochrome c; B/F: Endo G; C/G: GADD153; D/H: GRP78. Mitochondria and nuclei were counterstained with MitoTracker® Red CMXRos and PI (red color) respectively. Scale bar, 20 μm.
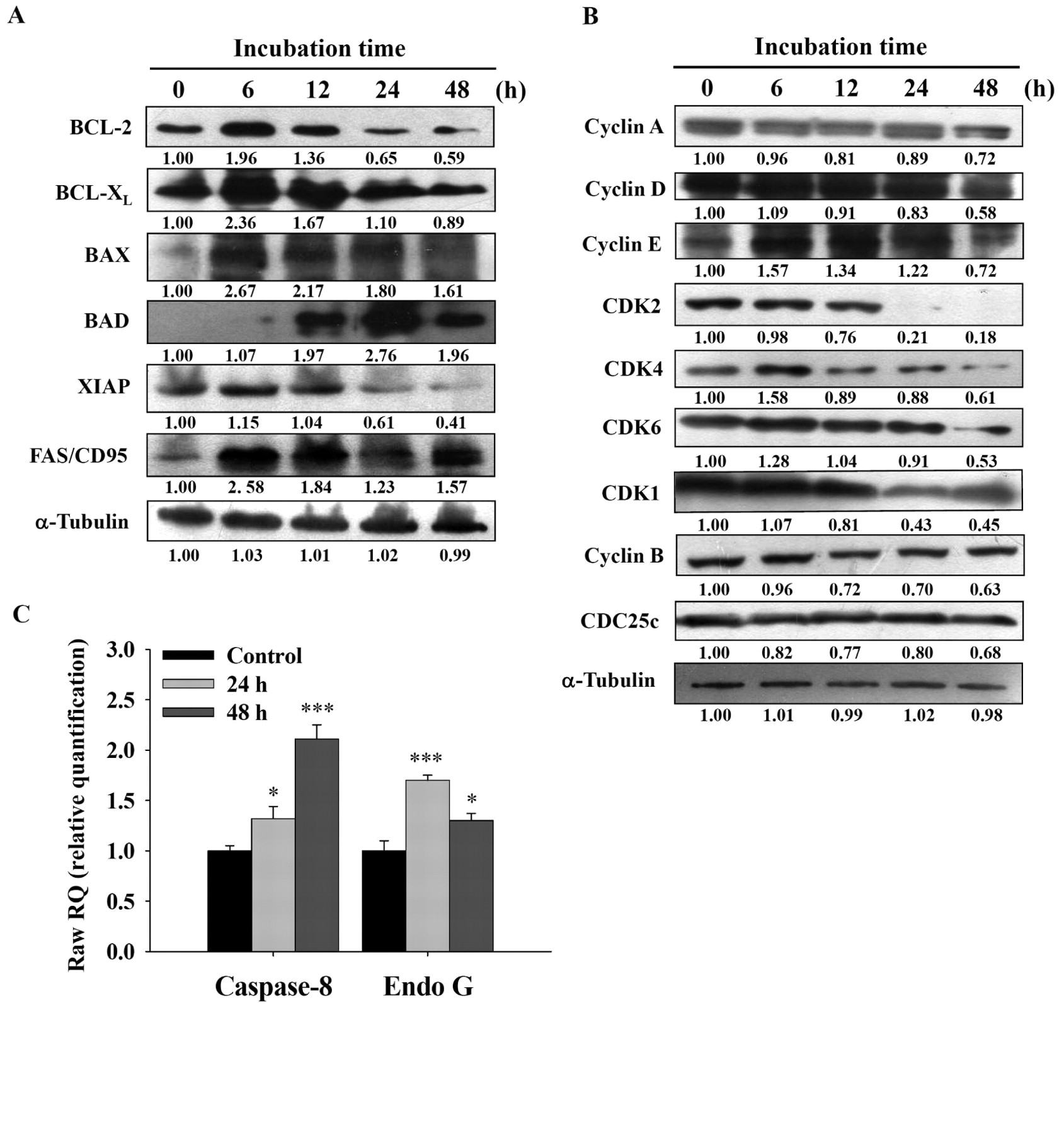
Representative Western blotting (A-B) showing changes in the levels of apoptosis and cell cycle arrest-associated proteins levels in NCI-H460 cells after treatment with curcumin. The cells (1×106 cells/ml) were treated with 25 μM curcumin for 0, 6, 12, 24 and 48 h then the total proteins were evaluated as described in the Materials and Methods. Effects of curcumin on the mRNA expressions of apoptosis-related genes in NCI-H460 cells by real-time PCR amplification (C). The asterisks indicate significant differences from the control at *p<0.05 and ***p<0.001 by Student's t-test.
Western blotting analysis showed increased FAS/CD95 expression in NCI-H460 cells after curcumin treatment (Figure 6A), combined with increased caspase-8 activity and mRNA (Figure 3 and 6C). We assumed the extrinsic apoptotic pathway was activated after exposure to curcumin. The assumption was confirmed by a significant increase in cell viability after pretreatment of curcumin-treated NCI-H460 cells with caspase-8 inhibitor (Figure 4A and B). These findings are in accordance with other studies, which showed curcumin induced caspase-8 activation in melanoma cells (32), gastric cancer cells, colon cancer cells (33), prostate cancer cells (34) and leukemia cells (35). FAS/CD95 is a membrane receptor and is usually activated by binding with FAS ligand (FASL). Bush et al. explicitly demonstrated curcumin was able to activate FAS in melanoma cells independently of FASL (32). Other anticancer drugs also triggered such FASL-independent FAS/CD95 activation (36-37).
After curcumin treatment of NCI-H460 cells, BAX and BAD were up-regulated, whereas BCL-2 and BCL-XL were down-regulated (Figure 6A). These changes displaced the balance between pro- and anti-apoptotic BCL-2 family members on mitochondrial outer membranes towards apoptosis (38), which caused a loss of ΔΨm (Table II) and the leakage of cytochrome c into the cytosol (Figure 5A). Cytosolic cytochrome c activated caspase-9, which trans-activated caspase-3 (Figure 3) and apoptotic cell death ensued. The Endo G released from mitochondria was also translocated into nuclei (Figure 5B), where it may mediate caspase-independent apoptosis (39). Due to these effects, chromatin condensation and DNA degradation (characteristic of apoptosis) result. The above events are comprised the mitochondria-dependent apoptotic pathway. Curcumin induced apoptosis in A549 cells only through ER stress pathway and mitochondria-dependent pathways (12); however, curcumin induced apoptosis in NCI-H460 cells through death receptor, ER stress and mitochondria-dependent signaling pathways. This is our novel finding in NCI-H460 cells after exposure to curcumin. Moreover, after curcumin treatment, the BCL-2, BCL-XL and XIAP were down-regulated in a time-dependent manner, and the pro-apoptotic proteins such as BAX, BAD, FAS/CD95 were up-regulated. In curcumin-treated A549 cells only BCL-2 was down-regulated. Therefore, our results suggest different levels of proteins of the BCL-2 family are involved in curcumin-induced apoptotic cell death of human non-small cell lung cancer cell lines.

The proposed mechanisms of curucmin-induced apoptotic cell death in human non-small cell lung cancer NCI-H460 cells.
ROS includes free radicals such as superoxide (O −2), hydroxyl radicals (•OH) and non-radical derivatives of oxygen such as H2O2. Curcumin, although well-known for its antioxidant effect, induced ROS generation in NCI-H460 cells (Table II). Curcumin can easily pass through the plasma membrane due to its unique physicochemical properties. Crossing of the plasma membrane by curcumin causes flipping of phosphatidylserine (40), which in turn produces extracellular superoxide in the presence of NADPH oxidase (41). Such extracellular ROS can diffuse through anion channels (42) or non-specifically (43) into cells. Intracellular ROS, in the presence of transition metals, can damage DNA and other intracellular organelles and lead to apoptosis (44). In this study, the role of ROS in curcumin-induced apoptotic cell death of NCI-H460 cells was demonstrated by pre-treatment with NAC, a ROS scavenger, and the subsequent restoration of cell viability (Figure 4C).
ROS can inhibit ER calcium pumps and facilitates the dumping of the stored calcium from the ER into cytosol. Mitochondria take up the calcium and initiate apoptosis through opening of their permeability transition pores (45). In addition, depletion of ER calcium impairs proper folding of proteins in the lumen of ER and causes ER stress. The early responses to ER stress include transcriptionally induced GRP78 production. GRP78 is a chaperone which helps fold nascent unfolded proteins and relieves ER stress (46). If the ER stress persists, GADD153 will be induced transcriptionally, and is a transcription factor regulating genes involved in apoptosis. The exact mechanism of GADD153-mediated apoptosis has yet to be elucidated, but it seems to work through a BCL-2 family-regulated mitochondrial pathway (47). The complex mechanism of curcumin-induced apoptosis in NCI-H460 cells is depicted in Figure 7.
Dysregulation of the cell cycle is associated with tumorigenesis (48). We found that NCI-H460 cells were arrested at the G2/M stage after treatment with curcumin (Figure 2A). Down-regulation of CDK1 may be involved in the mechanism of this arrest (Figure 6B). According to the studies on other type of cancer, tumor cells arrested at the G2/M stage are protected from the cytotoxic effect of curcumin (49). If they go beyond the G2/M stage, they are prone to apoptosis (50). Cell cycle regulation and apoptosis are like two sides of the same coin (51). Curcumin exerts its anticancer effects through both the cell cycle arrest and apoptotic induction.
In summary, curcumin exerts cytotoxicity on NCI-H460 cells through induction of apoptosis and apoptotic cell death involves the intrinsic, extrinsic and ER stress signaling pathways (Figure 7). Curcumin also promoted G2/M phase arrest in NCI-H460 cells. Both apoptotic induction and cell cycle arrest contribute to the anticancer effects of curcumin. Taken together, the data indicate that curcumin may be a potentially effective therapy for human large cell carcinoma of the lung.
Acknowledgements
The study was supported by grant 98-CCH-IRP-08 from Changhua Christian Hospital (Changhua, Taiwan, R.O.C.).
- Received October 6, 2009.
- Revision received April 12, 2010.
- Accepted April 22, 2010.
- Copyright© 2010 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Curcumin Triggers DNA Damage and Inhibits Expression of DNA Repair Proteins in Human Lung Cancer Cells
- Translating Curcumin to the Clinic for Lung Cancer Prevention: Evaluation of the Preclinical Evidence for Its Utility in Primary, Secondary, and Tertiary Prevention Strategies
- The Molecular Basis for the Pharmacokinetics and Pharmacodynamics of Curcumin and Its Metabolites in Relation to Cancer
- Safrole Induces G0/G1 Phase Arrest via Inhibition of Cyclin E and Provokes Apoptosis through Endoplasmic Reticulum Stress and Mitochondrion-dependent Pathways in Human Leukemia HL-60 Cells
- Gallic Acid Induces G0/G1 Phase Arrest and Apoptosis in Human Leukemia HL-60 Cells through Inhibiting Cyclin D and E, and Activating Mitochondria-dependent Pathway
- Effect of Agaricus blazei Murrill Extract on HT-29 Human Colon Cancer Cells in SCID Mice In Vivo
- Etomidate Induces Cytotoxic Effects and Gene Expression in a Murine Leukemia Macrophage Cell Line (RAW264.7)
- Zanthoxylum ailanthoides Sieb and Zucc. Extract Inhibits Growth and Induces Cell Death through G2/M-phase Arrest and Activation of Apoptotic Signals in Colo 205 Human Colon Adenocarcinoma Cells
- Crude Extracts of Agaricus brasiliensis Induce Apoptosis in Human Oral Cancer CAL 27 Cells through a Mitochondria-dependent Pathway
- Phenethyl Isothiocyanate (PEITC) Promotes G2/M Phase Arrest via p53 Expression and Induces Apoptosis through Caspase- and Mitochondria-dependent Signaling Pathways in Human Prostate Cancer DU 145 Cells