


Abstract
Background: Obesity is associated with an increased risk of estrogen-dependent breast cancer. Recently, concerns have been raised regarding the role of zeranol (Z), a non-steroidal anabolic growth promoter with potent estrogenic activity widely used in the U.S.A. beef industry, as a possible contributor to an increased incidence of human breast cancer. This study hypothesized that obese individuals may be at greater risk of developing zeranol-induced breast cancer. Materials and Methods: The aromatase mRNA expression level of three cell types isolated from adipose tissues were assayed by RT-PCR, and the cell proliferation of primary cultured human normal breast pre-adipocytes (HNBPADs) was investigated using the CellTiter96® non-radioactive method. The effects of Z and gossypol on aromatase expression of leptin-pretreated HNBPADs were evaluated by RT-PCR. Results: HNBPADs expressed higher aromatase than primary cultured human breast epithelial cells and stromal cells. Z enhanced the mitogenic activity of leptin and increased aromatase expression in HNBPADs. Moreover, (−)-gossypol counteracted Z- and leptin-induced cell proliferation and aromatase expression. Conclusion: These results suggested that bioactive Z metabolites contained in meat produced from Z-implanted beef cattle may increase estrogen biosynthesis in obese individuals by increasing aromatase expression and estrogen production, which will promote cell sensitivity and increase breast cancer cell growth.
Breast cancer is the second leading cause of cancer-related deaths in women and is the most common cancer among women today in the U.S.A. One risk factor for breast cancer is obesity, which has been shown to increase breast cancer risk especially in postmenopausal women (1, 2). It has been found that obese women with breast cancer have reduced survival rates and an increased chance of recurrence (3). Meanwhile, obesity has increased at an alarming rate and is now a worldwide public health problem. In recent years, the relationship between obesity and breast cancer has received increasingly more attention (2).
The mechanism of obesity-induced estrogen-dependent breast cancer is remain poorly understood. One of the possible contributing factors is the increased exposure of mammary gland epithelial cells to estrogens produced locally by adipose tissue (1). Some studies have shown that the association between obesity and higher risk for breast cancer in postmenopausal women may largely be explained by increased levels of estrogens, particularly bioavailable 17β-estradiol (4, 5). Upon binding to estrogen, an estrogen receptor activates transcription of its target genes, which are responsible for cancer cell proliferation in estrogen-dependent breast tumors (6). The biosynthesis of estrogens in breast tissue is catalyzed by cytochrome P450 aromatase (P450arom). Aromatase is the product of the CYP19A1 gene, a member of the P450 superfamily of genes that catalyses the rate-limiting step in estrogen biosynthesis. The conversion of C19 androgenic steroids to the corresponding estrogen is a reaction termed aromatization, since it converts the Δ4-3-one A-ring of the androgen to the phenolic A-ring of estrogen (7). This is the major mechanism of estrogen synthesis in postmenopausal women. The enzyme aromatase is found in a number of human tissues and cells, including ovarian granulosa cells, the placental syncytiotrophoblasts, adipose and skin fibroblasts, bone and the brain (8). Furthermore, adipose tissue, where aromatase transcripts primarily reside, is the principal site of estrogen production in postmenopausal women (9).
Another mechanism explaining the association between obesity and breast cancer implicates leptin, a cytokine synthesized in adipose tissue. Leptin acts as an important signaling molecule in energy regulation and food intake, which may also play an important role in the process of initiation and progression of human cancer (10, 11). Numerous studies have shown that leptin induces the proliferation of several breast cancer cell models (12-15), which leads to leptin being considered as a novel growth factor. Leptin is not expressed in healthy breast tissue in general, but does exist in the healthy tissue adjacent to malignant tumors (16, 17). Its receptor isoforms, ObRa and ObRb, are detectable in breast cancer cells but are undetectable in normal breast epithelial cells (18-20). In clinical studies, the serum level of leptin in obese cancer patients was found to be significantly higher than that in the healthy control group (21, 22). In a breast cancer research program, 60% of patients expressed leptin, while none of the cases with normal breast tissue expressed leptin (16). A previous study demonstrated that when the primary cultured human breast cancer epithelial cells were pre-treated with 3 nM leptin for 24 h, the sensitivity to zeranol exposure greatly enhanced the mitogenic action of leptin.
There is also a possible interaction between leptin and estrogen systems. Studies have demonstrated that leptin induces aromatase activity in MCF-7 cells, evidencing its important role in enhancing in situ estradiol production and promoting estrogen-dependent breast cancer progression (23). Estrogen has previously been proposed as a positive regulator of leptin production (24). There have been reports that estrogen modulates ObR expression in some estrogen-responsive tissues (24, 25).
Zeranol (commercial name: Ralgro®) is a non-steroidal agent with potent estrogenic activity that is widely used as a growth promoter in the U.S.A. beef industry. There is concern regarding the potential human breast cancer health risks of zeranol residues in the edible tissues of the food-producing animals implanted with zeranol. This is largely based on its demonstrated estrogenic activity (26). A previous study showed that zeranol transforms the human normal breast epithelial cell line MCF-10A and increases primary cultured human normal and cancerous breast epithelial cell growth in a dose-dependent manner (27). Furthermore, it was found that implantation of zeranol in beef cattle greatly induces pre-adipocyte growth by up-regulating cyclin D1 and down-regulating P53 expression (28).
Gossypol is a natural polyphenolic compound extracted from cottonseed glands and used as an anticancer chemopreventive agent. It is well documented that gossypol inhibits the proliferation of many human cancer cell types such as breast (29-31), colon (32) and prostate (33-34) cancer in vitro and in vivo. Previously, research on gossypol in relation to cancer had focused mainly on the antiproliferative activity of racemic gossypol, (±)-gossypol. Furthermore, it was found that (−)-gossypol is a more potent inhibitor than (+)-gossypol and (±)-racemic gossypol (35, 36). In addition, a previous report showed that gossypol is able to inhibit aromatase activity by interfering with folliclestimulating hormone induction of aromatase in cultured porcine granulosa cells (37).
In the present study, human breast epithelial cells, pre-adipocytes and stromal cells were isolated from human normal breast and adipose tissues. The aromatase expression in these cell types was characterized to determine the specific cell for aromatase and it was determined whether the aromatase expression in such cells pretreated with leptin, as a cell model of obesity, may be enhanced by zeranol and suppressed by (−)-gossypol, as a candidate chemopreventive agent. This study may implicate a potential association of adverse health risk of breast cancer and obesity to women consuming beef products derived from zeranol-implanted beef cattle.
Materials and Methods
Reagents. Recombinant human leptin was obtained from R&D Systems (Minneapolis, MN, USA) and was prepared as stock solution of 1 mg/ml in sterile 20 mM Tris/HCl (pH 8.0) at −20°C; zeranol was obtained from Sigma Chemical Co. (St. Louis, MO, USA); (−)-gossypol was provided by USDA Southern Regional Research Center (New Orleans, LA, USA) and was prepared as 50% stock solution in dimethyl sulfoxide (DMSO).
Human breast normal and cancer tissues. Human breast normal and cancer tissues were obtained through the U.S.A. National Cancer Institute-sponsored Tissue Procurement Program of the Ohio State University Comprehensive Cancer Center Hospital (Columbus, OH, USA). At the time of procurement, the tissue samples were placed in Dulbecco's modified Eagle's medium and Ham's F12 medium (1:1) (DMEM/F12) without phenol red (Sigma Chemical Co.) and stored at 4°C before transfer to the laboratory.
Isolation and culture of epithelial cells, pre-adipocytes and stromal cells from human breast tissues. Human breast tissues were sterilized in 70% ethanol for 30 s and then washed three times with fresh DMEM/F12. They were minced and then digested using a digestion buffer which consisted of phenol red-free high-calcium DMEM/F12 (1.05 mM CaCl2) with 2% bovine serum albumin (Invitrogen, Carlsbad, CA, USA) containing 10 ng/ml cholera toxin (Sigma Chemical Co.) 6,300 U/ml collagenase (Invitrogen) and 100 U/ml hyalurinidase (Calbiochem, Gibbstown, NJ, USA). After the mixture was incubated in a humidified incubator (5% CO2, 95% air, 37°C) overnight, the solution was transferred to a 50 ml tube and centrifuged in 200 × g for 5 min. The upper, middle and lower layers were separated and centrifuged again. The upper and middle layers, containing pre-adipocytes and stromal cells, respectively, were transferred to another 15 ml tube separately, while the lower layer contained epithelial cells. All the pellets were washed by DMEM/F12 medium with an antimycotic antibiotic (100 U/ml penicillin G sodium, 100 g/ml streptomycin sulfate and 0.25 g/ml amphotericin B) (Gibco BRL, Bethesda, MD, USA) and centrifuged again. This wash procedure was repeated three times. The epithelial cell pellet was resuspended in low-calcium DMEM/F12 (0.04 mM CaCl2) supplemented with Chelex-100 (Bio-Rad Laboratories, Richmond, CA, USA)-treated 10% fetal bovine serum (FBS) (Gibco Cell Culture; Gibco, Grand Island, NY, USA), the pelleted pre-adipocytes were resuspended in phenol red-free high-calcium DMEM/F12 (1.05 mM CaCl2) supplemented with 10% FBS and the pelleted stromal cells were resuspended in the same high-calcium DMEM/F12 supplemented with 5% FBS and 5 μg/ml insulin. All three isolated specific cell types were plated separately in 75 cm2 culture flasks in a humidified incubator (5% CO2, 95% air, 37°C). The media of all primary cultured human breast cells and human breast cancer cells were changed every two days. When the cells grew to 85% confluence, they were washed twice with calcium- and magnesium-free phosphate-buffered saline (PBS) with pH 7.3 and then trypsinized with 0.5% trypsin-5.3 mM EDTA (Gibco BRL) in PBS for 10 min at 37°C. The trypsinization stopped by addition of culture medium with 5% FBS. After centrifugation, the dissociated cells were resuspended in the same medium and subcultured into 75 cm2 culture flasks at a ratio of one to five flasks.
Cell proliferation assay (MTT assay). A total volume of 100 μl medium containing 4,000 human normal breast pre-adipocytes (HNBPADs) was seeded in each well of a 96-well plate in DMEM/F12 medium and incubated in 37°C for 24 h. The following day, the medium was replaced by 100 μl DMEM/F12 supplemented with 0.2% BSA and incubated in 37°C for another 24 h. The HNBPADs were treated with zeranol, leptin or a combination with (−)-gossypol, while the control group was treated with 0.1% DMSO. The proliferation of HNBPADs was measured by adding 20 μl fresh mixture of 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxy-phenyl)-2-(4-sulfophenyl)-2H-tetrazolium (MTS) and phenazine methosulfate (PMS) (20:1) solution (Promega, Madison, WI, USA) to each well. After incubation at 37°C for 1 to 3 h, optical density values were measured by a kinetic microplate reader (Molecular Devices Cooperation, Menlo Park, CA, USA) at 490 nm wavelength and the cell growth was compared.
Cell treatment and total RNA isolation. Treatments and total RNA extractions were performed on cells not propagated beyond the third passage and the viabilities of each cell type were greater than 95%, as determined by the trypan blue dye exclusion method (38). A total of 105 viable cells/well pre-adipocytes or stromal cells were seeded in six-well plates in 5 ml high-calcium DMEM/F12 supplemented with 10% Chelex-100-treated FBS medium and cultured overnight. The medium was changed to phenol red-free high-calcium DMEM/F12 supplemented with dextran-coated charcoal (DCC) (Dextran T-70; Pharmacia Fine Chemicals Co., Uppsala, Sweden; activated charcoal; Sigma Chemical Co.)-stripped Chelex-100-treated FBS (5%). After 24 h, the cells were treated with zeranol, leptin or (−)-gossypol and vehicle as controls in phenol-red-free high-calcium DMEM/F12 supplemented with 5% DCC-treated FBS for 24 h. The epithelial cells underwent the same procedure, only replacing high-calcium DMEM/F12 with low-calcium DMEM/F12. Total RNA was isolated in 1 ml TRIzol Reagent (Invitrogen) according to the manufacturer's instructions.
cDNA synthesis. RNA concentration was measured by a DU-70 spectrophotometer (Beckman Instruments Inc., Fullerton, CA, USA). RNA (1 μg) from cultured cells were reverse transcribed with 200 U M-MLV reverse transcriptase (Invitrogen) at 37°C for 50 min then at 70°C for 15 min in the presence of 1 μl 10 mM dNTP (10 mM each dATP, dGTP, dCTP, and dTTP at neutral PH) (Invitrogen), 1 μl 50 μM random hexamer (Amersham, Piscataway, New Jersey, USA), RNase Inhibitor (Invitrogen), 10 μl 5× first strand buffer, 5 μl 0.1 M DTT and 1 μl RNase inhibitor (Invitrogen) in a total volume of 50 μl in a gradient mastercycle (Eppendorf, Westbury, NY, USA).
Reverse transcription-polymerase chain reaction (RT-PCR). RT-PCR conditions were optimized for every primer pair and performed with a thermocycler Gene Amp PCR (Eppendorf). The newly synthesized cDNA (2 μl) was used as a template for RT-PCR and 1 U platinum Taq DNA polymerase (Gibco BRL) was added in a total volume of 25 μl. MgCl2 concentration, annealing temperature and cycle number were optimized for the amplification of the PCR product, CYP19A1. The primer sequences were: 5′-CCT GGC TAC TGC ATG GGA AT-3′ (sense) and 3′-GCC TTT CTC ATG CAT ACC GA -5′ (antisense, product size 246 bp). The PCR protocol was: denaturation at 95°C for 5 min, 36 cycles of 94°C for 45 s, 60°C for 45 s, 72°C for 60 s, then extension at 72°C for 10 min. The primer sequences for 36B4, an internal control, were: 5′-AGC TGA TCA AGA CTG GAG ACA AA-3′ (sense) and 3′-GGG TAG CCA ATC TGC AGA CA-5′ (antisense, product size 220 bp). The PCR protocol was: denaturation at 95°C for 5 min, 29 cycles of 95°C for 60 s, 60°C for 60 s, 72°C for 60 s, then extension at 72°C for 10 min. 36B4 RNA is unmodified by treatment.
The final RT-PCR products (10 μl) mixed with 1 μl 10× loading buffer were separated on 1.5% agarose gel and visualized by staining with ethidium bromide. Electronic images were taken under a Fujifilm LAS-3000 image system (Fujifilm Medical Systems USA, Inc., Stamford, CT, USA). The densities of specific bands were quantified by ImageQuant software (Molecular Dynamics, Sunnyvale, CA, USA). The results are presented as the ratio of CYP19A1 to 36B4.
Statistical analysis. The results for the cell proliferation assay are presented as mean±standard deviation for four replicate culture wells. Analysis was performed using Minitab 15 (Minitab Inc., State College, PA, USA). Statistical difference was determined by using two-sample t-test analyses for independent samples. P-values < 0.05 were considered statistically significant.
Results
Comparison of aromatase mRNA expression levels in normal and cancerous human breast tissues. To compare the mRNA expression of aromatase in matched healthy and cancerous human breast tissues, CYP19A1 mRNA levels were determined by RT-PCR in fresh tissue obtained from reduction mammoplasty of three breast cancer patients. Figure 1 shows that breast cancer tissue had 2.8-3.5-fold higher levels of CYP19A1 mRNA than the normal breast tissue.

Comparison of aromatase mRNA expression levels in normal and cancerous human breast tissues. A: Representative image of gel electrophoresis of RT-PCR. B: Each bar represents the mean±SD of 3 experiments quantified by MultiGauge software.
Comparison of aromatase mRNA expression levels among epithelial, stromal cells and pre-adipocytes. In order to determine the cell-specific characteristics of aromatase expression, three cell types, which included epithelial cells, stromal cells and pre-adipocytes, were isolated from cancer tissue from the same patient and their CYP19A1 mRNA expression levels (Figure 2) showed that pre-adipocytes have 44% and 147% higher aromatase mRNA expression than stromal cells and epithelial cells respectively, suggesting a cell-specific expression pattern of the aromatase gene.
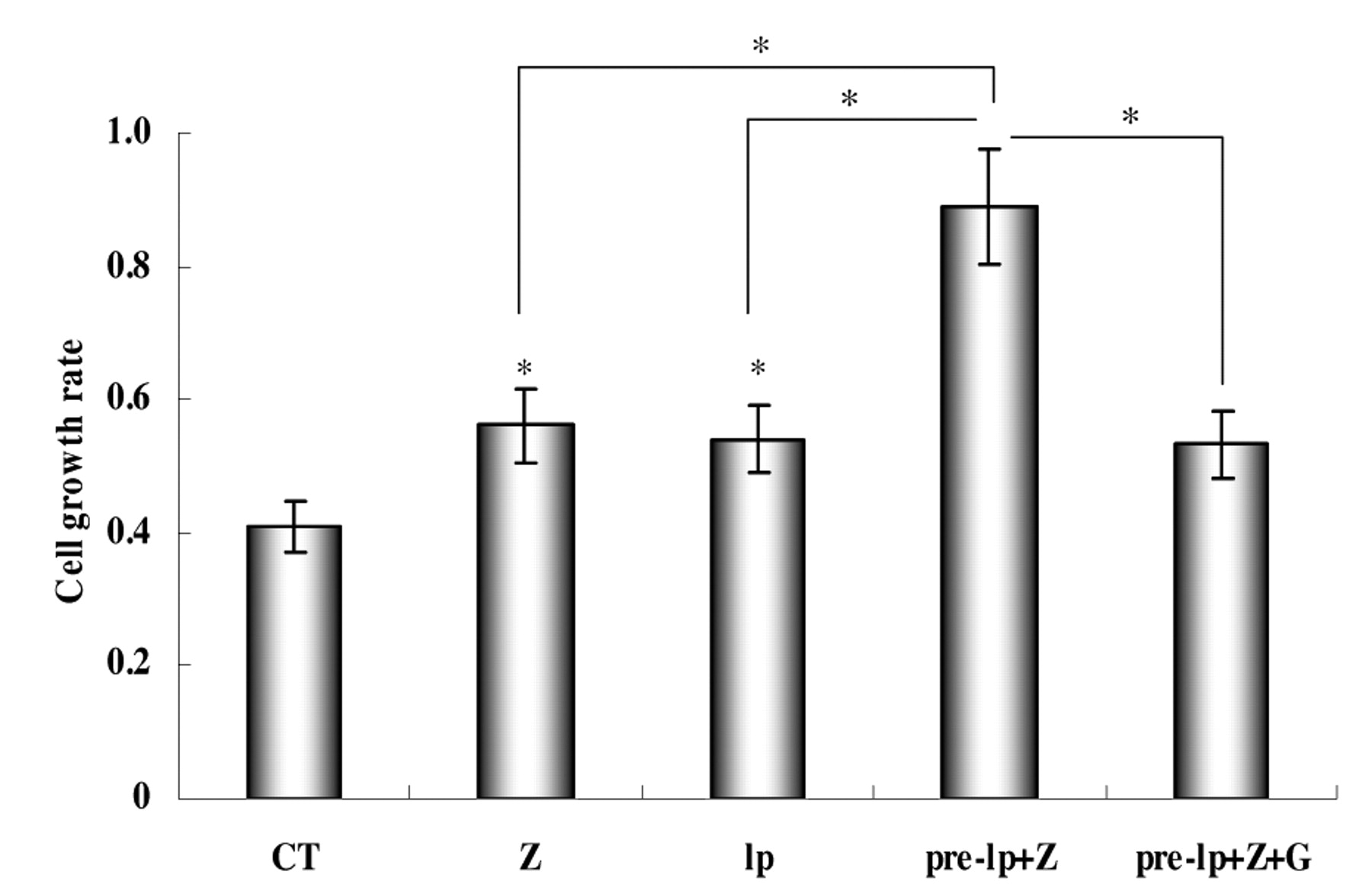
Effect of zeranol and (−)-gossypol on proliferation of leptin-pretreated pre-adipocytes. The cell proliferative activity of zeranol and leptin on the MCF-7 cell line has been demonstrated in a previous study (39). The present study focused on pre-adipocytes. As shown in Figure 3, 20 nM zeranol and 1.5 nM leptin significantly increased the growth of pre-adipocytes by 37% and 32%, respectively, after 48 h treatment. A functional cross-talk between zeranol and leptin in MCF-7 was shown in a previous report (39). The present study examined their interaction on cell proliferation. Pre-adipocytes were treated with vehicle, zeranol, leptin and their combination and cell growth was assayed using MTS method. As Figure 3 shows, 20 nM zeranol exposure significantly enhanced the proliferation of leptin (1.5 nM)-pretreated pre-adipocytes by 65%. The results showed that the potency of zeranol to increase pre-adipocyte growth was enhanced by pretreatment with leptin. (−)-Gossypol has been demonstrated as an anticancer compound in many previous studies. Figure 3 also shows that stimulation of growth with zeranol in leptin-pretreated pre-adipocytes may be significantly suppressed by (−)-gossypol at a dose of 5 μM.

Comparison of aromatase mRNA expression levels among primary cultured human breast epithelial cells, pre-adipocytes and stromal cells isolated from the same patient. A: Representative image of gel electrophoresis of RT-PCR. B: Each bar represents the Mean±SD of 3 experiments quantified by MultiGauge software.
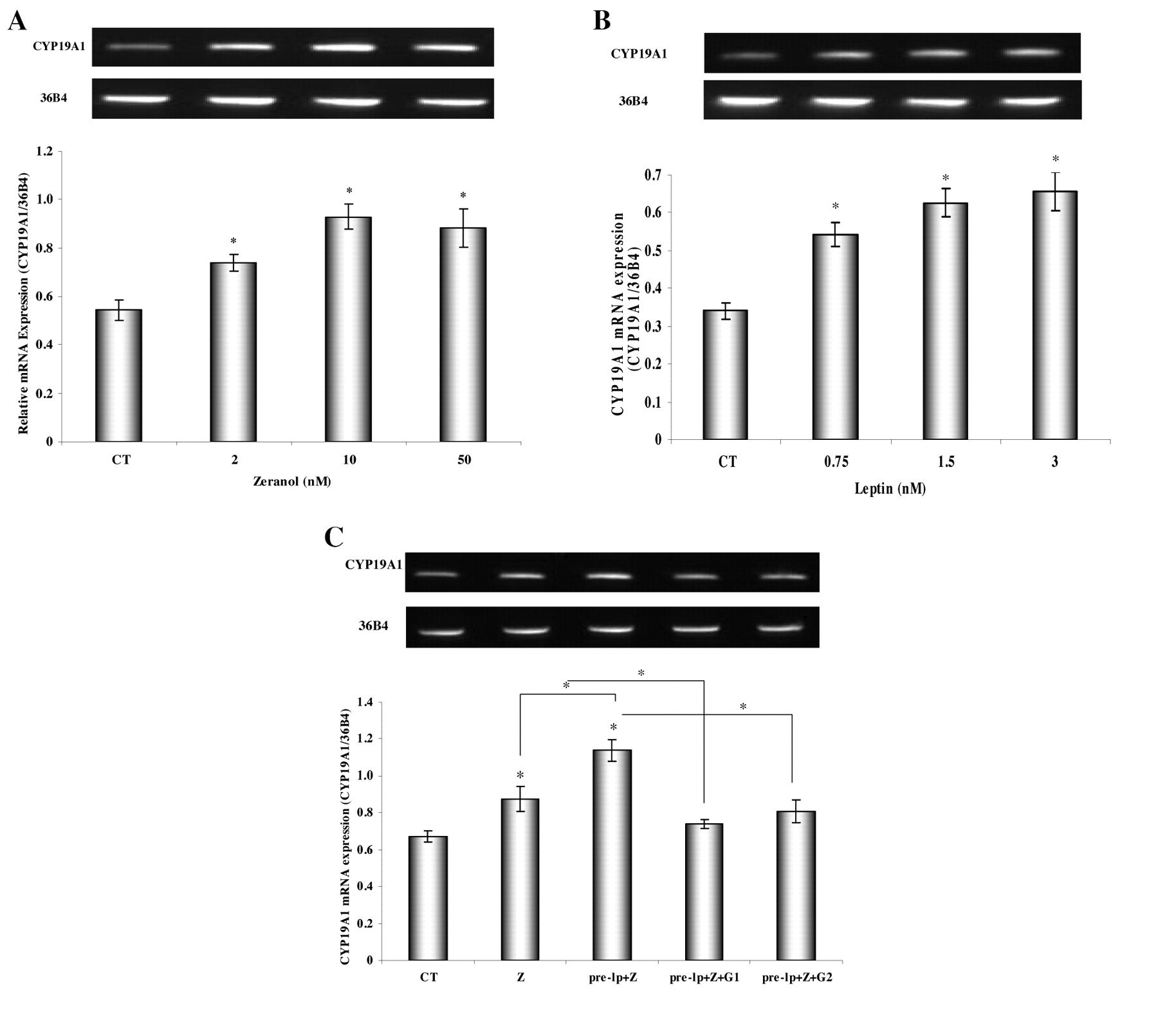
Effect of zeranol and (−)-gossypol on aromatase mRNA expression on leptin-pretreated HNBPADs. Figure 4A shows that aromatase mRNA expression was significantly increased by the chosen doses of zeranol in HNBPADs. Figure 4B shows that pretreated pre-adipocytes with leptin for 24 h enhanced the aromatase mRNA expression in a dose-dependent manner between 0.75 nM and 3 nM. Figure 4C shows that 20 nM zeranol significantly enhanced the aromatase mRNA expression of leptin (1.5 nM)-pretreated pre-adipocytes by 30%, while (−)-gossypol at the dose of 5 μM and 2.5 μM significantly suppressed the leptin- and zeranol-induced aromatase expression up-regulation by 35% and 29%, respectively.
Discussion
Estrogen is a well-known carcinogenic factor in the development and mitogenic stimulation of breast cancer. Aromatase, a cytochrome P-450 enzyme, is generally considered the most important enzyme that catalyzes the conversion of androgens to estrogens, which is the major mechanism of estrogen biosynthesis in postmenopausal women and effective aromatase inhibitors are explored for use in the treatment of human breast carcinoma as an estrogen deprivation therapy.
Previous work has indicated an intriguing link between aromatase and cancer. Utsumi et al. reported that the aromatase mRNA levels in breast cancer tissues are significantly higher than in nonmalignant breast tissues (40). There have also been some reports showing that aromatase activity in tumors or surrounding tissues may play a significant role in promoting tumor growth due to this local estrogen production (41, 42). The present study confirmed that the expression levels of aromatase mRNA are significantly higher in breast cancer tissues than in non-neoplastic breast tissues from the same patient (Figure 1).
Previous studies have demonstrated that aromatase is present in both epithelial and stromal cells. A question remains with regard to which cell type is the most important for aromatase expression. Stromal pre-adipocytes are immature adipocytes that express the S-100 protein, a marker that is not found in fibroblasts or endothelial cells. The role of pre-adipocytes in tumor development is still controversial. Some studies suggest that pre-adipocytes stimulate the growth of breast tumor cells (43), while other studies show that pre-adipocytes are either inhibitory or have no effect on breast cancer cell proliferation (44, 45).
The data of Figure 2 demonstrates that the ranking of cell types for aromatase according to their relative importance would be: pre-adipocytes > stromal cells > epithelial cells. This suggests that pre-adipocytes may contribute to the most important mechanism of estrogen biosynthesis in human breast adipose tissue and may be associated with stimulation of breast epithelial cancer cell growth. The primary cultured human normal pre-adipoctyes may serve as an excellent model to study the possible effects of zeranol, leptin and gossypol on the local biosynthesis of estrogens. Moreover, previous work from this research group has proven that the purity of the epithelial cell and stromal cell populations, which were isolated following the procedures described in the present study, is more than 95%, based on cytokeratin and vimentin by immunohistochemistry (46) and the purity of the pre-adipocytes was also verified by red oil O staining (data not shown).
Previous studies have shown that both zeranol and leptin or their combinations are able to induce cell proliferation in the MCF-7 cell line and primary cultured human breast cancer epithelial cells. Furthermore, it was found that pretreatment with leptin increases the sensitivity of MCF-7 Adr cells and primary cultured human breast cancer epithelial cells to zeranol, thus, enhancing their proliferation (39, 47). The present data (Figure 3) indicate that both treatments with 20 nM zeranol and 1.5 nM leptin for 48 h are able to induce significant pre-adipocyte proliferation and the pretreatment with leptin for 24 h is able to increase the sensitivity of pre-adipocytes to zeranol, thus stimulating cell growth. In contrast, this stimulated cell proliferation may be suppressed by 5 μM (−)-gossypol. These findings further implicate the possible relationship between obesity and breast cancer and point to the potential risk for breast cancer in obese people, especially in those consuming meat from animals treated with zeranol, since leptin is able to increase the cell sensitivity to zeranol.
Furthermore, previous studies (39, 47) have shown that leptin enhances the sensitivity of MCF-7 Adr cells to zeranol and increases cell growth via increasing cyclin D1 mRNA expression. Exposure to zeranol may lead to the transformation of normal breast cells to breast preneoplastic cells by up-regulating cyclin D1 and down-regulating P53. However, a possible effect of zeranol, leptin and their combination on the local biosynthesis of estrogens had not been examined yet. In order to study whether zeranol may enhance the aromatase expression in leptin-pretreated pre-adipocytes, thus increasing the local estrogen biosynthesis in human breast adipose tissue, the aromatase mRNA expression was studied. The results, determined using RT-PCR assay (Figure 4), indicated that aromatase expression in pre-adipocytes was significantly increased by treatment with 2 nM to 50 nM zeranol and increased with dose-dependent manner with leptin at the doses ranging from 0.75 nM to 3 nM. Pretreatment with 1.5 nM leptin for 24 h significantly enhanced the zeranol effect of increasing the aromatase expression in pre-adipocytes. Such characteristics of leptin may increase aromatase expression at the postmenopausal state where leptin may be higher in a non-obese person. Moreover, the increasing level of aromatase expression in leptin-pretreated pre-adipocytes induced by zeranol may be suppressed by 2.5 μM and 5 μM (−)-gossypol. These findings may lead to the consideration of zeranol as a stimulator for estrogen production in obese patients by increasing aromatase expression in adipose tissue. In contrast, (−)-gossypol may be an alternative aromatase inhibitor for breast cancer therapy.

Effects of zeranol and (−)-gossypol on proliferation of HNBPADs. Data are presented as mean±standard deviation for four replicate experiments. CT: 0.1% DMSO as control; Z: zeranol (48 h); lp: 1.5 nM leptin (48 h); pre-lp+Z: 1.5 nM leptin pretreated for 24 h and 20 nM added Z for another 24h; pre-lp+Z+G: 1.5 nM leptin pretreated for 24 h and 20 nM Z+5 μM (−)-gossypol added for another 24 h; *statistically significant difference (p<0.05).
In summary, the findings reported here extend previous work to explain the implication that obese individuals may be at greater risk of developing zeranol-induced breast cancer. Estrogen release from pre-adipocytes may act as a functional signal linking adipose tissue to epithelial tissue and zeranol in vivo may increase estrogen biosynthesis, thus, promoting further mammary cell proliferation. The use of the co-culture system to clarify whether the production of estrogen from the pre-adipocyte response to aromatase enhancement is able to promote epithelial cell growth will be investigated in a future study.

A: Effect of zeranol on aromatase mRNA expression in HNBPADs. HNBPADs were treated with 2 nM, 10 nM or 50 nM zeranol. B: Effects of leptin on aromatase mRNA expression in HNBPADs. HNBPADs were treated with 0.75 nM, 1.5 nM or 3 nM Leptin. C: Effects of zeranol and (−)-gossypol on aromatase mRNA expression on leptin-pretreated HNBPADs. Data are presented as the mean±standard deviation for three replicate experiments. CT: 0.1% DMSO as control; Z: zeranol (48 h); pre-lp+Z: 1.5 nM leptin pretreated for 24 h and 20 nM Z added for another 24 h; pre-lp+Z+G1: 1.5 nM leptin pretreated for 24 h and added 20 nM Z+5 μM (−)-gossypol for another 24h; pre-lp+Z+G2: 1.5 nM leptin pretreated for 24 h and 20 nM Z+2.5 μM (−)-gossypol added for another 24h; *statistically significant difference (p<0.05). For each figure, the top panel is a representative image of gel electrophoresis of RT-PCR. In the bottom panel, each bar represents the mean±SD of 3 experiments quantified by MultiGauge software.
Acknowledgements
This research was supported by NIH R01 Grant ES 015212. Saiyi Zhong was supported by the China Scholarship Council (Beijing, P.R. China) (CSC No. 20093012).
- Received October 18, 2010.
- Revision received November 13, 2010.
- Accepted November 16, 2010.
- Copyright© 2010 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved





{kind=link}
{kind=link}
{kind=link}
{kind=link}