


Abstract
Background: Vascular endothelial growth inhibitor (VEGI) has been implicated in the regulation of tumour-related vasculature in certain solid tumours. However, its role in urothelial tumours is still unclear. In the present study, the role played by VEGI in urothelial tumours of the bladder was investigated. Materials and Methods: The expression of VEGI was examined in cancer human bladder tissues and in a serial of cell lines using immunochemical staining and RT-PCR respectively. The biological impact of modifying VEGI expression in urothelial cancer cells was evaluated using in vitro models. Results: VEGI mRNA was expressed in a wide variety of human cell lines. VEGI expression was seen in normal urothelial and stromal cells, but was found to be reduced or absent in urothelial cancer cells. The positive staining in normal tissue (6/7) was significantly higher than that of urothelial cancer tissues (2/12), p=0.006. Moreover, overexpression of VEGI reduced the motility and adhesion of urothelial cancer cells in vitro. However, the overexpression of VEGI had no bearing on the growth and invasion of urothelial cancer cells. Conclusion: VEGI has an inhibitory effect on cellular motility and adhesion in bladder cancer cells. Taken together with the expression pattern of VEGI in urothelial cancer of the bladder, it suggests that VEGI functions as a negative regulator of aggressiveness during development and progression of bladder cancer.
Bladder cancer is the fourth most common cancer in men and the second most common malignancy affecting the genitourinary system (1). About 90% of all bladder tumours are urothelial carcinomas, which are also called transitional cell carcinomas (TCCs) (2). It is a heterogeneous disease with considerable variations of its natural history. For example, the five-year survival rate is 97-98% for patients with a monofocal, well-differentiated and small papillary tumour, whereas this could be 0% if patients have invasive bladder cancer extending throughout the bladder wall and with gross nodal metastases. It is also characterized as a carcinoma with multifocality and a high recurrence rate (3). Recent studies have provided evidence that angiogenic factors, such as basic fibroblast growth factors (FGFs) and vascular endothelial growth factor (VEGF), are correlated with this aggressive aspect (4). It is likely that they are involved not only in the formation of new vessels, but also through juxtacrine/paracrine mechanisms in the induction of motility in malignant urothelial cells. For example, VEGF is up-regulated in bladder cancer compared with normal urothelial, and the increased VEGF protein levels correlate with tumour recurrence (5).
Vascular endothelial growth inhibitor (VEGI) was first reported as an angiogenesis inhibitor in 1997 (6). It is a member of the tumour necrosis factor superfamily and is also known as tumour necrosis factor superfamily member 15 (TNFSF15) and TNF ligand-related molecule 1 (TL1) (6-8). The full gene of VEGI is approximately 17 kb, which consists of four exons and three introns and mapped to human chromosome 9q32. Its amino acid sequence has 20-30% homology to other TNF family members, except TNFβ (8-10). Three isoforms of VEGI exist, these may arise due to splicing variation. All of them share a common region of 453bp that encodes a domain of 151 amino acids at the C-terminus. The biological activity of VEGI is mainly dependent on the solubilised extracellular domain of the three isoforms. It is able to induce apoptosis in endothelial cells via an autocrine pathway (8, 11, 12). Its mRNA has also been found in a variety of normal and tumour tissues and in tumour cell lines, suggesting a potential role in both physiological and pathological conditions. Overexpression of VEGI has been shown to inhibit tumour neovascularisation and progression in cellular and animal models (8, 11, 12). The protein was also found to be able to inhibit the growth of breast, colon and Lewis lung cancer in murine tumour models (7, 8, 13). Parr et al. reported that patients with breast tumours expressing reduced levels of VEGI had a higher local recurrence, shorter survival time and an overall poorer prognosis than those patients expressing high levels of VEGI (14). In additional, VEGI is also a T-cell co-stimulator and can directly stimulate dendric cell maturation (12, 15). However, it is still unclear whether activation of tumour-specific or nonspecific B or T lymphocytes may also operate in soluble VEGI-mediated tumour suppression.
Primer sequences for current study.
Considered together, these studies imply that VEGI plays a critical role in tumour suppression. However, the role of VEGI in urothelial cancer of bladder remains unknown. In the current study, the expression of VEGI was examined in urothelial cancer of bladder specimens and bladder cancer cell lines. The biological function of this molecule was investigated in cells which were manipulated to express enhanced levels of VEGI in order to establish the functional role of VEGI in bladder cancer cells.
Materials and Methods
Materials. All cell lines used in this study were obtained from the European Collection for Animal Cell Culture (ECACC, Porton Down, Salisbury, UK). Cells were routinely cultured with Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% foetal calf serum and antibiotics. Monoclonal mouse anti-human-VEGI and anti-human-GAPDH antibodies were purchased from Santa Cruz Biotechnology, Inc. (SC-53975 and SC-47724 respectively; Santa Cruz, CA, USA).
Nineteen bladder samples were collected from patients of the Department of Urology, University Hospital of Wales, including 12 bladder tumour tissues and 9 normal background bladder tissues. These tissues were collected immediately after radical cystectomy and transurethral resection of bladder tumour. All protocols were reviewed and approved by the local Ethical Committee and all patients gave written informed consent.
Immunohistochemical staining of bladder specimens. Frozen specimens of bladder cancer (n=12) and normal bladder tissue (n=7) were cut at a thickness of 6 μm using a cryostat (Leica CM 1900; Leica Microsystems UK Ltd., Buckinghamshire, UK). The nature of the samples was independently verified by two pathologists. After fixation, the sections were blocked with horse serum and probed with or without VEGI antibody (SC-53975) for 1 h. T24 cells that had been transfected with the VEGI expression construct were used as a positive control. The secondary biotinylated antibody and the Avidin Biotin Complex were subsequently applied to detect VEGI expression in accordance with the Vectastain Universal Elite ABC kit protocol (Vector Laboratories, Peterborough, UK). After developing colour with diaminobenzidine (DAB), the sections were counterstained with Gill's Haematoxylin. Staining was independently assessed by the authors.
Total RNA preparation and reverse transcription-polymerase chain reaction (RT-PCR). Total RNA was isolated from cells using a Total RNA Isolation Reagent (Advanced Biotechnologies Ltd, Epsom, and Surrey, UK). First strand cDNA was synthesized from 0.5 μg RNA using a reverse transcription kit (Sigma, Poole, Dorset, UK). The quality of cDNA was verified through the amplification and detection of the GAPDH housekeeping gene. VEGI forward and reverse primers (Table I) were designed based on the human VEGI sequence (Genbank Accession number: BD131562). PCR was performed in a GeneAmp PCR system 2400 thermocycler (Perkin-Elmer, Norwalk CT, USA). Conditions for PCR were 40 s at 94°C, 60 s at 55°C, 60 s at 72°C (35 cycles). PCR products were separated on a 1.4% agarose gel.
Generation of the VEGI expression construct and transfection into T24 cells. Sets of primers were designed to amplify the entire coding region of human VEGI (Table I). The correct products were T-A cloned into the open reading frame of a mammalian expression vector, pEF6/V5-His vector (Invitrogen). The recombinant plasmid vectors were transformed into chemically competent TOP10 E. coli (Invitrogen, Inc., Paisley, UK). After verification, the expression constructs and empty control plasmid were amplified, purified and used to transfect T24 cells by way of electroporation (Easyjet, EquiBio Ltd, Kent, UK). Following a period of selection with blasticdin antibiotic, levels of VEGI were assessed in transfected T24 cells to confirm sufficient overexpression before being used in cell assays.
Quantitative real-time polymerase chain reaction (Q-PCR). Real-time quantitative PCR was carried out using the iCycleriQ5 system (Bio Rad, Hemel Hemstead, UK) and a recently described method (14) to determine the level of the VEGI and GAPDH transcripts in the cell lines. GAPDH was used as an internal control to allow normalization of VEGI expression in the samples. Q-PCR conditions were 95°C for 15 min, followed by 65 cycles at 95°C for 15 s, 55°C for 30 s and 72°C for 15 s.
Western blot analysis of VEGI expression. Protein concentration in cell lysates was determined using the DC Protein Assay kit (BIO-RAD laboratories, CA, USA) and an ELx800 spectrophotometer (BIO-TEK, Wolf Laboratories, York, UK). Equal amounts of proteins were separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and blotted onto nitrocellulose sheets. Proteins were then probed with the VEGI antibody (sc-53975, 1:1,500) and peroxidase-conjugated secondary antibodies. GAPDH (sc-47724, 1:1,000) was used as a housekeeping control. Protein bands were visualized using the Supersignal™ West Dura system (Pierce Biotechnology, Inc., Rockford, IL, USA), and photographed using an UVITech imager (UVITech, Inc., Cambridge, UK).
Immunocytochemistry of VEGI expression. Cells were fixed with 4% formaldehyde and then permeabilised with 0.1% Triton X-100 for 5 min in Tris buffer saline (TBS). After blocking with horse serum (10%) for 60 min, the cells were probed with anti-VEGI antibody for 1 h, followed by extensive washing. Horseradish peroxidase-conjugated anti-mouse antibody was then added for 1 h and visualised using the Vectastain ABC system. Slides were mounted and the staining density of VEGI in cells was analysed using the Optimas 6.0 software package.
In vitro cell growth assay. Cell growth was assessed using a method previously reported by our laboratories (16). Briefly, the cells were plated into a 96-well plate (2,500 cells/well). Cells were fixed in 4% formaldehyde on the day of plating and days 1, 3, and 5 after plating and then stained with 0.5% (w/v) crystal violet. Following washing, stained crystal was extracted with 10% (v/v) acetic acid and absorbance determined at a wavelength of 540 nm using a spectrophotometer (BIO-TEK).
In vitro invasion assay. This was performed according to a standard procedure (17-19). Transwell inserts with 8 μm pore size were coated with 50 μg Matrigel (BD Matrigel™ Basement Membrane Matrix; BD Biosciences, Oxford, UK) and air dried. Matrigel was rehydrated before use. A total of 20,000 cells were added to each well. After 96 hours, cells that had migrated through the matrix to the other side of the insert were fixed in 4% formaldehyde, stained with 0.5% (w/v) crystal violet and counted under a microscope.
In vitro cell matrix adhesion assay. Cell matrix adhesion was assessed according to a previously described method (17-19). A total of 40,000 cells was added to each well of a 96-well plate, previously coated with Matrigel (5 μg/well). After 40 minutes of incubation, non adherent cells were washed off using balanced saline solution (BSS) buffer. The remaining adherent cells were then fixed and stained with crystal violet. The number of adherent cells in random fields were observed and counted under a microscope.
In vitro motility assay using Cytodex-2 beads. We followed the protocol previously described (17-19). A total of 1×106 cells was incubated with 100 μl of cytocarrier beads (GE Healthcare Life Sciences, Buckinghamshire, UK) in 10 ml DMEM overnight. The beads were washed twice to remove dead cells and then resuspended in 800 μl DMEM. 100 μl of beads/cells were transferred into each well of a 24-well plate. After incubation for 4 h, the medium was aspirated and cells were fixed with 4% formaldehyde for 5 minutes. They were then stained with 0.5% crystal violet for 5 minutes. The cells were washed and allowed to dry before counting a number of random fields per well.
In vitro migration assay (wounding assay). The migration of cells across a wounded surface of a near-confluent cell monolayer was examined (17-19). Cells at a density of 50,000/well were seeded into a chamber slide and allowed to reach near confluence. The monolayer of cells was then scraped with a fine gauge needle to create a wound of approximately 200 μm. The movement of cells to close the wound was recorded using a time lapse video recorder and analysed using the motion analysis feature of the Optimas 6.0 software package.
Electric cell-substrate impedance sensing (ECIS) based attachment and motility assay. The ECIS-1600R model instrument and 8W10 arrays (Applied Biophysics, Inc., NJ, USA) were used in the study. We used a method recently reported (20). Briefly, the same number of test cells (300,000 per well) was added to each well of the ECIS arrays. Impedance and resistance of the cell layer was immediately recorded for a period of up to 20 h. When confluence was reached, the monolayer in each well was electrically wounded at 6 V for 30 s to create 10 uniform wounds per well. Impedance and resistance of the wounded cells as they migrated in the wound was then recorded for a period of up to 20 h. Data was analysed using the ECIS RbA modelling software, supplied by the manufacturer.
Statistical analysis. All statistical analysis was performed using the SPSS 16.0 software (SPSS Inc., Chicago, IL, USA). Two-sample t-tests were used for normally distributed data. Fisher's exact test was used for analysing immunohistochemical staining in bladder tissues. Differences were considered to be statistically significant at p<0.05.
Results
The expression of VEGI in cell lines and bladder tissues.
Detection of VEGI transcript by RT-PCR. The expression of VEGI was examined in 27 human normal and cancer cell lines using RT-PCR (listed in Table II). VEGI transcript was detectable in most cell lines (21/27) except BT549, EJ138, HRT18, MRC5, A549 and Corl47 (Figure 1 A), as reported in previous studies. Interestingly, in bladder cancer cell lines, the T24 cell line expressed low levels of VEGI, and VEGI expression was almost absent for EJ138 cells.
Human cancer and normal cell lines used in the current study.
Detection of VEGI protein by immunohistochemistry. In immunohistochemical staining, VEGI was seen in normal bladder urothelial and stromal cells, but the staining was decreased in or absent from malignant urothelial cells (Figure 1 B). The positive staining of normal tissue (6/7) was significantly higher than that of bladder cancer tissues (2/12), p=0.006.
Transfection of VEGI in the T24 cell line.
Expression of VEGI transcript as shown by RT-PCR. Two human VEGI expression plasmids were constructed, coded as VEGIexp4 and VEGIexp7. T24 cells, which expressed low level of VEGI transcript, were transfected with VEGI expression constructs. As shown by RT-PCR analysis. VEGI mRNA expression was increased in T24VEGIexp4 and T24VEGIexp7 cells compared with wild-type (T24wt) cells and empty plasmid control (T24pEF/His) cells (Figure 2 A). Q-PCR showed that VEGI mRNA expression was dramatically increased in T24VEGIexp4 and T24VEGIexp7 cells, compared to corresponding controls (Figure 2 B).
VEGI overexpression as shown by immunocytochemistry. Overexpression of VEGI was also reflected at the protein level using immunocytochemistal staining and Western blot analysis. Increased protein levels of VEGI were seen in cells transfected with the VEGI expression construct (T24VEGIexp4) compared to controls (Figure 2 C, D and E). T24VEGIexp4 had higher efficiency than T24VEGIexp7 and was used in the subsequent experiments.
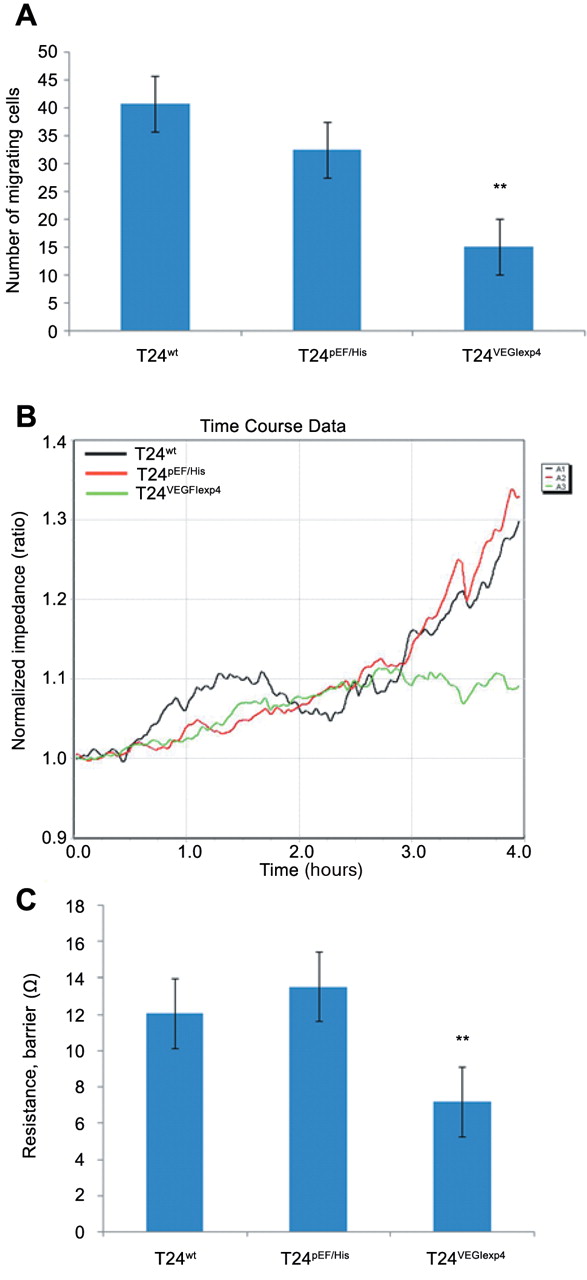
The influence of VEGI overexpression on cell matrix adhesion in urothelial cancer cells. We first examined the effect of VEGI on the cell matrix adhesion of bladder cancer cell lines. Overexpression of VEGI exhibited a significant inhibitory effect on cell matrix adhesion of the cells. Compared with T24wt (36.5±6.68) and T24pEF/His (32.6±4.25), the number of adherent cells for T24VEGIexp4 (15.7±4.75) was significantly reduced (p<0.001 vs. both controls) (Figure 3 A).
The ECIS system was used to further investigate the effect of enhanced expression of VEGI on T24 cell adhesion. The attachment capacity was remarkably reduced in T24VEGIexp4 cells compared with T24wt and T24pEF/His cells (Figure 3 B and C).
Effect of VEGI on motility of urothelial cancer cells. In the cytocarrier based cell motility assay, we found that cellular motility was significantly reduced in T24VEGIexp4 cells. The number of migrating T24VEGIexp4 cells was 15.2±4.45 compared with 40.8±23.29 for T24wt cells and 32.5±6.76 for T24pEF/His cells (p<0.001 in both comparisons) (Figure 4A).
The effect of VEGI on T24 cellular motility was also assessed using the ECIS system. The migrating capacity was remarkably reduced in T24VEGIexp4 cells compared with T24wt and T24pEF/His cells (Figure 4B and C).
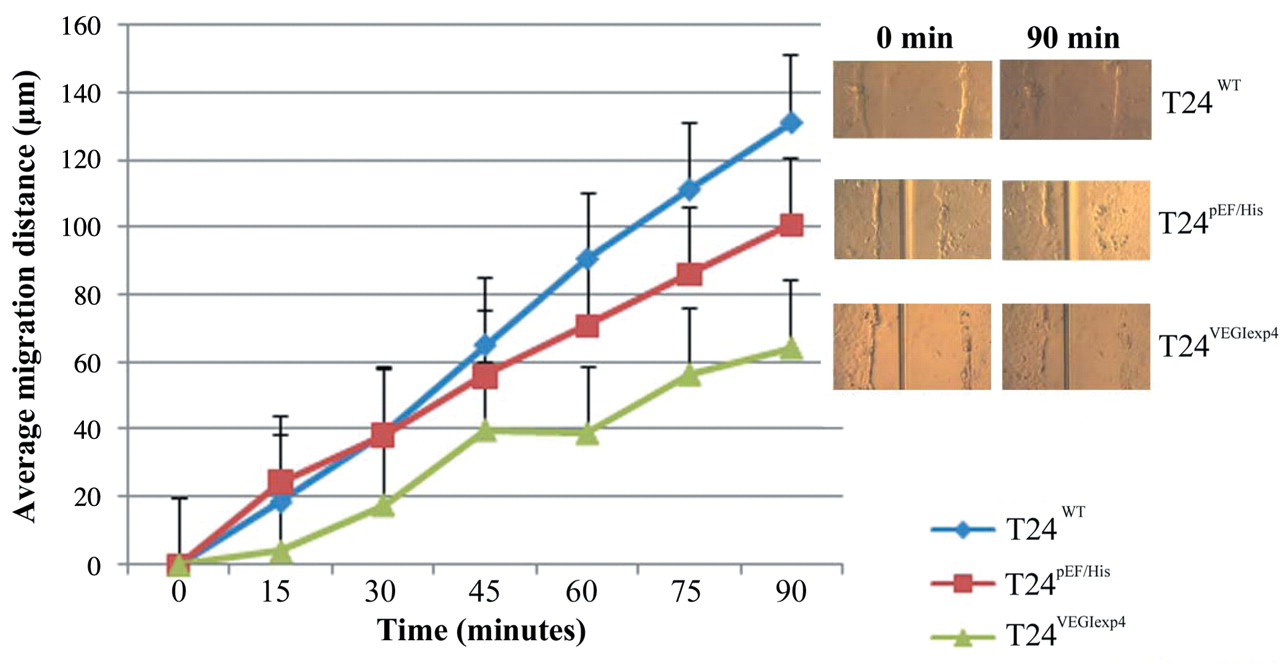
Similar trends were also seen using the scratch wounding assay to examine cellular migration where once again the motility of T24VEGIexp4 cells was reduced significantly compared to the controls. The average migrating distance of T24VEGIexp4 was 78.5±8.52 μm, p<0.001 compared with both T24wt (164.4±34.61 μm) and T24pEF/His (112.8±14.93 μm) cells (Figure 5).
Overexpression of VEGI had no impact on the growth or invasiveness of T24 cells. The growth and invasive capacity of T24VEGIexp4 cells was not found to be significantly different from that of wild-type and empty plasmid control cells.
Discussion
VEGI, first identified from human umbilical vein endothelial cells is known to exist abundantly in arterial endothelial cells (6-8). According to the literature, it is able to induce apoptosis in endothelial cells via an autocrine pathway (8, 11, 12). Overexpression of VEGI has been shown to inhibit tumour neovascularisation and progression in cellular and animal models (7, 12). In studies, using recombinant VEGI protein, Zhai et al. found that the protein could markedly inhibit the growth of breast and colon xenograft tumours and suggested that the effect may be through an indirect inhibition on capillary-like structures and cell growth (7, 8). Chew et al. also reported that overexpression of VEGI induced apoptosis in endothelial cells and inhibited the growth of xenograft tumours, together with a reduction in the microvessel density (11). Further investigation showed that the density of the endothelial cells exhibited an 88% decrease within 1 week of treatment with recombinant human VEGI and a further decrease within 3 weeks in Lewis lung cancer in murine tumour models. As much as 50% inhibition of the tumour growth rate was achieved with the treatment (13). VEGI prevents G0/G1 phase endothelial cells from re-entering the cell cycle in response to growth stimuli, and induces caspase-3-mediated apoptosis in actively proliferating cells (11, 21). These reports suggest that the antitumour effect of VEGI was likely due to the ability of VEGI to suppress neovascularisation, because recombinant VEGI had no inhibitory activity on the growth of the cancer cells in vitro except for some specific cells (such as human histiocytic lymphoma (U-937), human breast carcinoma (MCF-7), human epithelial carcinoma (HeLa) and human myeloid lymphoma (ML-1a)) (22).

VEGI Expression in bladder cancer tissues and cell lines. A, Detection of the VEGI transcript using RT-PCR in a panel of human cell lines. B, Immunohistochemical staining of human bladder specimens. B1 and B2 were normal bladder tissue. The VEGI protein was found to be well stained in the cytoplasmic area of normal bladder urothelial cells (indicated by black arrows). B3 and B4 were bladder cancer tissue. VEGI was only weakly stained in malignant urothelial tumours cells (black arrows).

Effects of overexpression on VEGI transcript in T24 cells. As show in RT-PCR (A), mRNA of VEGI was overexpressed in T24VEGIexp4 and T24VEGIexp7 cells, compared with the wild-type and empty plasmid control cells. The result of Q-PCR showed similar trends to RT-PCR. T24VEGIexp4 had a higher number of transcripts than T24VEGIexp7 cells. **Indicates p<0.01 versus T24wt and T24pEF/His cells. Q-PCR was performed in triplicates for each cell line and repeated independently three times. C and D: Forced expression of VEGI at the protein level using Western Blot analysis and immunocytochemistry for T24 cells. VEGI protein was found to be stained more strongly in the cytoplasmic region of T24VEGIexp4 cells than that of wild-type and empty plasmid control cells. The results of Western blotting were the same as those of ICC. E: The staining intensity of VEGI in T24 cells as determined by image analysis. **Indicates p<0.01 versus wild-type and empty plasmid control cells.

The effect of VEGI on cell matrix adhesion in vitro. All experiments were repeated 5 times. A: Effect of VEGI expression on cell adhesion to matrix-coated surface. Overexpression of VEGI reduced the number of adherent cells in T24VEGIexp4 cells. **indicates p<0.01 versus T42wt and T24pEF/His cells. Cell matrix adhesion assay carried out using 5 wells per cell line per experiment. Error bars represent the SD. B, Effect of VEGI expression on cell attachment as analysed by ECIS. The same number of cells (300,000 per well) were added to each well of the ECIS arrays. The impedance change is shown. The T24VEGIexp4 cells which overexpressed VEGI showed a markedly reduced attachment. C: ECIS RbA modelling of cell attachment indicated a significant reduction of attachment in VEGI transfected cells. **indicates p<0.01 versus wild-type and empty plasmid control cells.

Effect of overexpression of VEGI on the migration of T24 cells. All experiments were repeated 4 times. A: Migration of cells using the cytocarrier bead assay. Motility of T24VEGIexp4 cells was reduced remarkably, compared with wild-type and empty plasmid control cells. **Indicates p<0.01 versus T24wt and T24pEF/His cells. B: Effect of VEGI expression on cell migration as analysed by ECIS (wounding assays). Cell monolayers were first wounded at 6 V for 30 s. The impedance changes during the migration process are shown. The T24VEGIexp4 cells which overexpressed VEGI showed a markedly reduced migration. C: ECIS RbA modelling of cell attachment indicated a significant reduction of attachment in VEGI transfected cells. **Indicates p <0.01 versus wild-type and empty plasmid control cells.
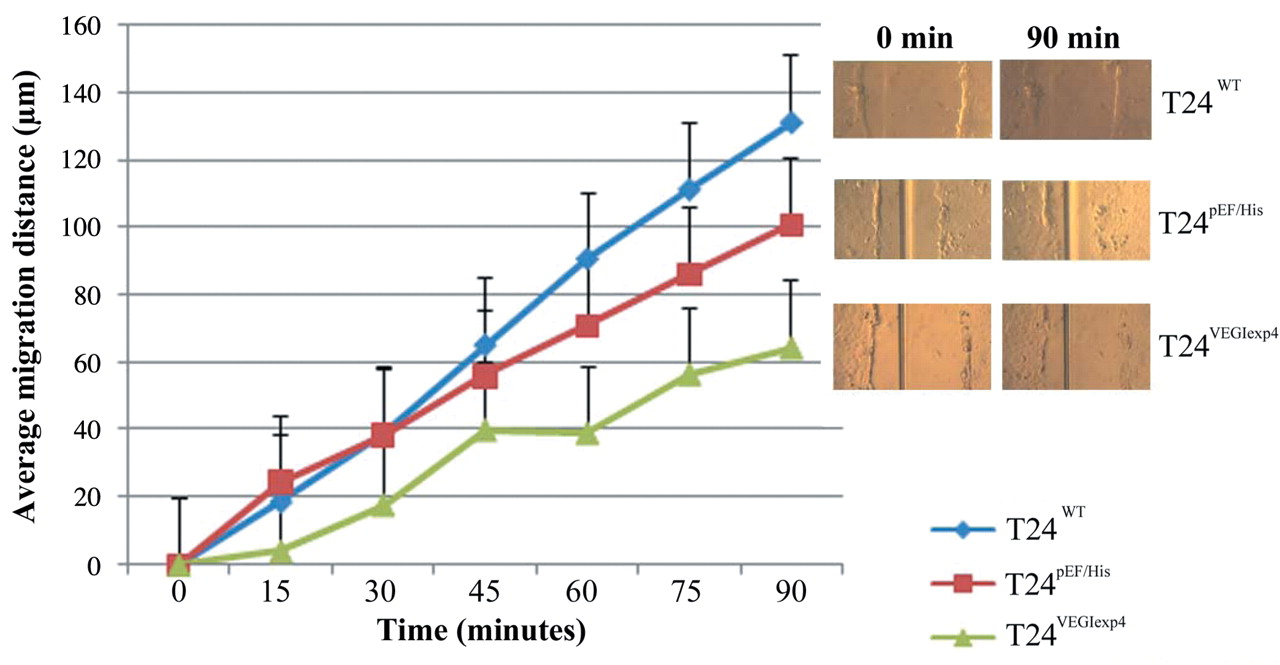
Effect of transfection of VEGI on migration using the scratch wounding assay. The experiment was repeated 3 times. The movement was reduced significantly in T24VEGIexp4 cells compared with wild-type and empty plasmid control cells (p<0.01).
VEGF as a key pro-angiogenic factor has been implicated in a variety of human tumours. VEGF is up-regulated in malignant urothelial compared with normal urothelial, and this increased expression associates with disease progression. However, the role played by VEGI in bladder cancer is largely unknown. In the present study, we first noted that VEGI protein levels were lower in urothelial cancer of bladder tissues compared with that of normal bladder tissues. Intensive staining was seen within the normal urothelial and stomal cells and was much weaker in or absent from malignant urothelial cells. Unfortunately, due to a limited number of samples, no correlations of VEGI expression and clinical aspects of the urothelial cancer could be made in this study. However, the decrease or absence of VEGI expression in urothelial bladder cancer at least suggests that there might be a shift in the balance between pro- and anti-angiogenic stimuli during the disease progression. This may result in a microenvironment conducive to tumour growth and survival (12, 14, 23).
Our result shows that the overexpression of VEGI can directly affect the motility and adhesion of urothelial tumour cells. Cell matrix adhesion in T24 cells transfected with the VEGI expression plasmid was found to be greatly reduced, displaying approximately 43% of the level seen in wild-type cells. The average distance of VEGI overexpressing cells also dramatically decreased compared with controls. It is suggested that VEGI can directly reduce the aggressiveness of urothelial cancer cells; this is in line with the observation that the expression of VEGI is decreased in urothelial bladder cancer specimens. Together with the observation that VEGI has little bearing on apoptosis and growth of urothelial tumour cells, these data indicate VEGI may be a profound cell adhesion and migration regulator. In addition, it is well known that multifocality and polychronotropism are two main characteristics of urothelial bladder cancer. According to the evidence that multifocal tumours occurring in the same individuals demonstrated clonality (24), reimplantation of tumour cells is one of the most commonly accepted theory to explain these characteristics (25, 26). The lower cell adhesion and motility, but lack of impact on invasion of urothelial tumour cells suggests that VEGI may be a key regulator of reimplantation of urothelial bladder cancer. This highlights an exciting direction to pursue in future scientific research. Together with reports that overexpression of VEGI inhibits tumour neovascularisation and directly stimulates dendritic cell maturation (27-30), an essential component of host immunity against cancer development, our study has provided further evidence to indicate a therapeutic implication of VEGI in bladder cancer.
In summary, we first noted that VEGI, an endogenous angiogenesis inhibitors, was decreased in urothelial bladder cancer. It can suppress the aggressiveness of bladder cancer cells through inhibiting cell migration and adhesion. It suggests that VEGI may be a putative tumour suppressive factor and a potential therapeutic target to prevent development and reimplantation in bladder cancer.
Acknowledgements
Dr Zhang is a recipient of Cardiff University China Medical Scholarship. The Authors wish to thank the Albert Hung Foundation and Cancer Research Wales for supporting their work and Dr David Griffiths and Dr M. Varna for their expert help in histological evaluation.
Footnotes
-
Conflict of Interest
None declared.
- Received June 24, 2009.
- Revision received December 1, 2009.
- Accepted December 1, 2009.
- Copyright© 2010 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- TNFSF15 facilitates the differentiation of CD11b+ myeloid cells into vascular pericytes in tumors
- Identification of Novel Proteins Interacting with Vascular Endothelial Growth Inhibitor 174 in Renal Cell Carcinoma
- Protein of Vascular Endothelial Growth Inhibitor 174 Inhibits Epithelial-Mesenchymal Transition in Renal Cell Carcinoma In Vivo
- Vascular Endothelial Growth Inhibitor 174 Is a Negative Regulator of Aggressiveness and Microvascular Density in Human Clear Cell Renal Cell Carcinoma
- The Differential Expression of Vascular Endothelial Growth Inhibitor Isoforms, VEGI251, VEGI174 and VEGI192 in Human Clear-cell Renal Cell Carcinoma
- Expression of Bone Morphogenetic Protein 7 in Lung Cancer and its Biological Impact on Lung Cancer Cells