


Abstract
Background: Tetrandrine, a bisbenzylisoquinoline alkaloid, has cancer cell cytotoxicity. The effects of combined tetrandrine and radiation, alone or combined, on human SH-SY5Y neuroblastoma cells were examined. Materials and Methods: A combination treatment, using either concomitant irradiation at the beginning or end of the tetrandrine treatment (designated as the RT-Tet and Tet-RT protocols, respectively), was used to investigate radiosensitization by tetrandrine. The level of radiosensitization was evaluated by the dose-enhancement ratio and isobologram analysis. The cell cycle distribution of the neuroblastoma cells was examined using flow-activated cell sorter (FACS) analysis. Results: Tetrandrine had a time- and concentration-dependent cytotoxic effect (p<0.05). The dose-enhancement ratio for RT-Tet and Tet-RT was increased, and higher for RT-Tet. Isobologram analysis revealed mainly synergistic cytotoxicity for RT-Tet, but only subadditive cytotoxicity for Tet-RT. FACS analysis revealed that radiation caused accumulation in the G2/M-phase of the cell cycle, while tetrandrine caused G0/G1 accumulation. Compared to RT alone, RT-Tet increased the G0/G1 fraction and decreased the G2/M accumulation (p<0.001), whereas Tet-RT led to no reduction in radiation-induced G2/M accumulation. Conclusion: Tetrandrine radiosensitization is sequence dependent, with stronger cytotoxic effects noted when radiation is delivered at the beginning of tetrandrine treatment. The effect is, at least partly, related to the partial abrogation of radiation-induced G2/M accumulation.
Neuroblastoma is the most common extracranial solid malignancy in children (1). The prognosis of neuroblastoma is poor even using multimodality treatment (1-3). There have been numerous attempts to develop an effective treatment strategy for neuroblastomas. Such strategies have included radiotherapy and/or chemotherapy.
Tetrandrine (C38H42O6N2, MW 622.730), a bisbenzylisoquinoline alkaloid, is extracted from the root of a Chinese herb, the creeper Stephania tetrandra S. Moore) (4). Tetrandrine has anti-inflammatory, immunosuppressive, and cytoprotective effects (4-6). It suppresses T-and B-cells and inhibits the production of cytokines including histamine, prostaglandins and tumor necrosis factor-α (7, 8). Tetrandrine can suppress inflammatory reactions associated with rheumatoid arthritis, uveitis (induced by bovine serum albumin, interleukin-1 (IL-1) or endotoxin) and hepatitis in animals, and prolong survival time after allogenic cardiac transplant in mice, lipopolysaccharide-induced fulminant hepatitis or endotoxemia in mice, and the proliferation of pulmonary fibroblasts (4, 9-13). In addition, tetrandrine is also a calcium channel blocker and can reduce portal venous pressure and blood pressure (6, 8, 14). Tetrandrine has antioxidative effects and can inhibit lipid peroxidation and platelet aggregation, as well as reduce ischemia/reperfusion injury (15, 16).
In addition to these actions, the inhibition of tumor cell proliferation and induction of apoptosis in several carcinomas (including breast and lung cancer, Burkitt's lymphoma, leukemia, malignant lymphoid and myeloid cells, hepatoma/hepatoblastoma and HeLa cells) have recently been noted (5, 8, 17-20). Furthermore, an antitumor effect on allogenic ascites tumors in mice and gliomas in rats has been reported (21, 22). In designing various treatment strategies for malignant tumors, the importance of combining chemotherapy and radiotherapy is increasing (23, 24). In addition, the sequence of combination therapy using chemotherapeutic agents and radiation for the treatment of malignant tumors is closely related to their effects (24). The optimal sequence depends on the mechanisms of the chemotherapeutic agents, which are variable. The effect of combined radiation and tetrandrine on carcinomas has rarely been investigated (25, 26) and no studies have looked directly at the optimal sequence of combined radiation and tetrandrine for cancer treatment. The effects of tetrandrine on neuroblastomas are unclear, and only one report has mentioned the cytotoxic effect of tetrandrine on neuroblastoma cells (27). Furthermore, radiation therapy is a major treatment strategy for neuroblastomas and the effects of combining radiation with tetrandrine on neuroblastomas are unclear. Thus, in this study, the antitumor effects of tetrandrine on neuroblastoma cells were explored, and radiosensitization by tetrandrine in vitro was investigated using human SH-SY5Y neuroblastoma cells.
Materials and Methods
Tissue culture and cell line. The human SH-SY5Y neuroblastoma cell line was purchased from the American Type Culture Collection (Rockville, MD, USA). The cells were maintained in minimal essential medium (MEM) and F12 with 10% fetal calf serum (FCS) at 37°C in a 5% CO2 incubator.
Preparation of tetrandrine. Tetrandrine was purchased from Sigma-Aldrich Chemical Co. (St. Louis, MO, USA) and dispersed into a fine suspension in 0.1 N HCl at a concentration of 25 mg/mL. It was then passed through a 0.22 μm filter for bacteriological sterilization. The preparation was sonicated and adjusted to pH 6.8 immediately before use.
Assay of cytotoxicity. Sensitivity of the SH-SY5Y cells to tetrandrine was determined in vitro by clonogenic assay. Briefly, 12-well trays were inoculated with 2 mL of medium containing 600 cells and incubated overnight. The cultures were then incubated with tetrandrine (0, 1, 5, 10, 25 or 50 μM) for 24 or 48 hours, washed free of drug with phosphate-buffered saline (PBS) and incubated in fresh, drug-free medium for 14 days. The colonies were fixed and stained with methanol containing 0.5% methylene blue to aid counting. Colony-forming efficiency (number of colonies formed/number of cells plated) ranged from 30 to 50%. The extent of cell proliferation and viability was then determined. The LC10, LC20, LC30, LC40 and LC50 of tetrandrine (the concentrations at which 90%, 80%, 70%, 60% and 50% of the neuroblastoma cells were killed) were determined. The LC50 for SH-SY5Y cells was compared with that reported in the literature for other cancer cells. The LC10, LC20, LC30 and LC40 for the SH-SY5Y cells under various combined-treatment conditions were compared as described below.
Radiation therapy. The effects of radiation (137Cs irradiation doses of 0, 25, 50, 100, or 200 cGy) on the SH-SY5Y cells were studied using the clonogenic assay (as described above). The extent of cell proliferation, viability and LC50 (the radiation dose at which 50% of the neuroblastoma cells were killed) were determined.
Tetrandrine-radiation protocols. To investigate the effects of tetrandrine and radiation treatment on the neuroblastoma cells, tetrandrine (0, 1, 5, 10 or 25 μM) for 24 or 48 hours and radiation (0, 25 or 50 cGy) were delivered concomitantly at the beginning of tetrandrine treatment (RT-Tet protocol), or radiation was delivered at the end of tetrandrine treatment (Tet-RT protocol) (24). The cell proliferation and viability were then determined using the clonogenic assay described above. The LC10, LC20, LC30 and LC40, as well as the dose enhancement ratio, were determined under various treatment conditions. The additivity of the combined treatment was assessed using isobologram analysis, which may be used when the pharmacological response to one or more of the therapeutic agents is non-linear (28). This method allows for evaluation of an additivity envelope, against which the combined-treatment data points are compared. Briefly, the locations of the data points with respect to the envelope indicate the type of interaction, with data points to the left of the envelope indicating positive effects (enhancement or supra-additivity), data points to the right of the envelope indicating negative effects (inhibition, antagonism, or sub-additivity), and data points within the envelope indicating independent mechanisms or a synergistic effect (28, 29).

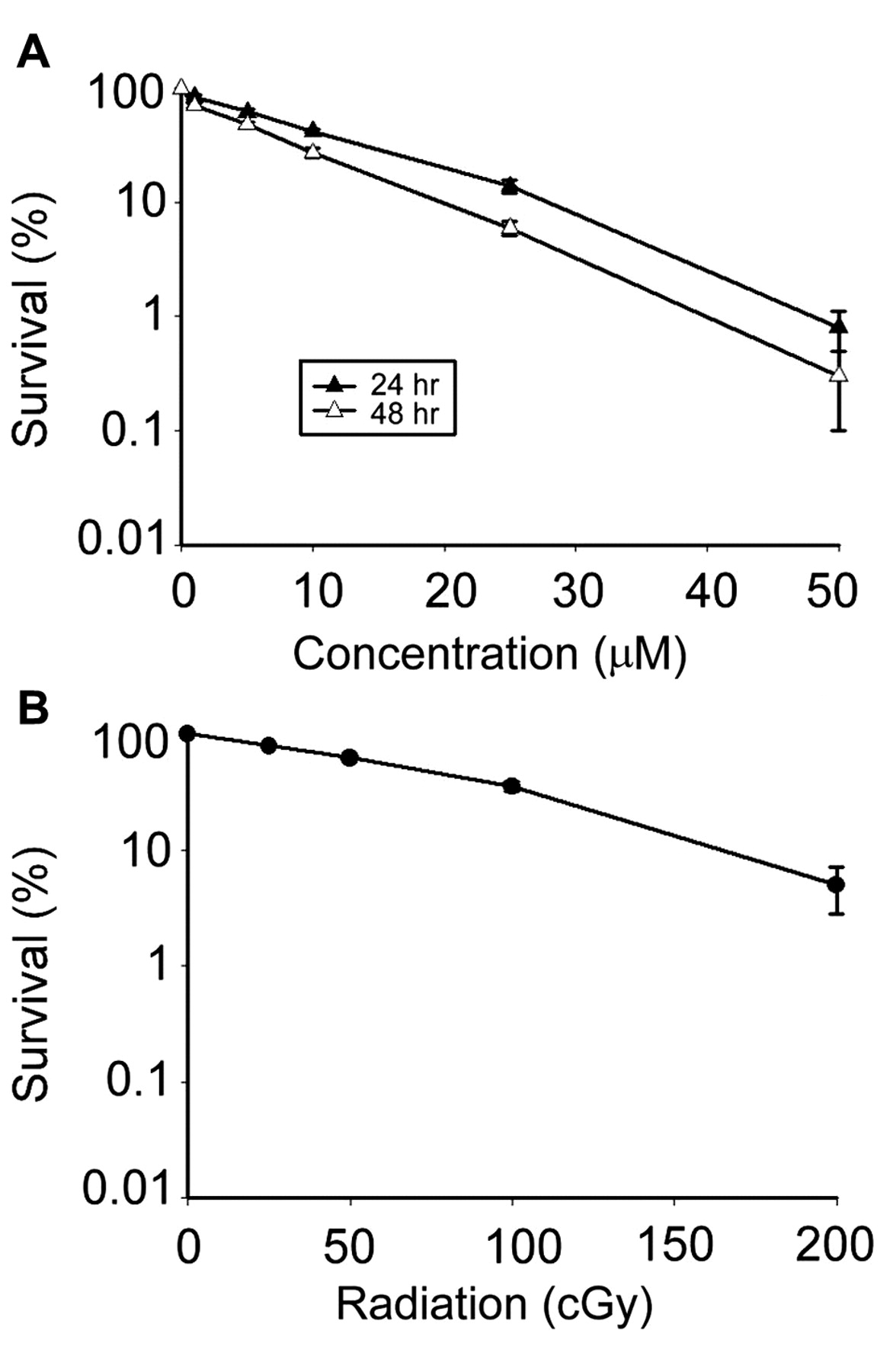
The cytotoxicity effects of tetrandrine and radiation. Survival curves for SH-SY5Y neuroblastoma cells (A) treated with various concentrations of tetrandrine, (B) treated with 137Cs irradiation. Cell viability was determined using the clonogenic assay. Each point is the average of three independent trials (nine determinations for each concentration) and presented as mean±standard deviation.
Analysis of the cell cycle distribution following treatment. Flow-activated cell sorter (FACS) analysis was used to investigate the effects of tetrandrine (0, 5 or 10 μM) for 24 hours and radiation (0, 25, or 50 cGy) combined according to either the RT-Tet or Tet-RT protocol on the cell cycle distribution of the cultured neuroblastoma cells. After various treatments, the neuroblastoma cells were trypsinized, washed twice with PBS, suspended in 1 mL 80% ethanol/PBS and stored at -20°C for subsequent analysis. Before FACScan analysis (FACSCalibur; Becton Dickinson Immunocytometry System, San Jose, CA, USA), the cells were centrifuged (3,500×g, 5 min), washed twice with PBS, incubated with 0.5 mL of 0.5% Triton® X-100/PBS and 5 μg RNase A for 30 min and stained with 0.5 mL of 50 μg/mL propidium iodide/PBS in the dark. The cell cycle distribution was then assessed.
Enhancement ratioa of the LC10, LC20, LC30 and LC40b of the SH-SY5Y neuroblastoma cells after 24- or 48-h tetrandrine treatment and radiation.
Statistical analysis. One-way analysis of variance (ANOVA) by Scheffe's post hoc multiple comparisons test was used for the statistical analyses of the survival curves and cell cycle distribution. P<0.05 was considered statistical significance.
Results
Tetrandrine induced time- and concentration-dependent cytotoxicity. The survival curves for the cells treated with tetrandrine are presented in Figure 1A. The effect of tetrandrine on the neuroblastoma cells was both concentration- and time-dependent, with higher concentrations and longer treatments having significantly greater cytotoxic effects (p<0.05). The LC10, LC20, LC30, LC40 and LC50 were 32.4, 21.8, 16.5, 11.2 and 8.0 μM, respectively, for 24-h tetrandrine treatment and 22.1, 15.1, 9.3, 6.9 and 4.7 μM, respectively, for 48-h tetrandrine treatment.
Radiation induced cytotoxicity. Radiation dose-dependently reduced the cell survival, with higher dosages killing more neuroblastoma cells (Figure 1B; p<0.05). The LC50 was 72.6 cGy.
Cell survival with tetrandrine treatment and early- or late-phase irradiation. Survival after tetrandrine for 24 or 48 hours plus radiation (0, 25, or 50 cGy) according to the RT-Tet and Tet-RT protocols is depicted in Figure 2. Analyses of the survival curves revealed that higher radiation dosages and longer tetrandrine exposure times were significantly more effective (p<0.05). Table I shows the dose-enhancement ratios of the RT-Tet and Tet-RT protocols, using either 24- or 48-h tetrandrine treatment. Combining 24- or 48-h tetrandrine treatment with 25 cGy irradiation, the dose-enhancement ratios were similar to for the Tet-RT protocols (1.3-1.7 vs. 1.3-1.8 fold). In contrast, the dose-enhancement ratio was higher when the irradiation dosage was 50 cGy (1.8-7.5 vs. 1.3-1.8 fold for 25 cGy) and when the protocol was 50 cGy RT-Tet (2.3-7.5 vs. 1.8-3.2 fold for the 50 cGy Tet-RT protocol). However, the dose-enhancement ratios for the 24-h and 48-h tetrandrine treatment were similar in both the RT-Tet and Tet-RT protocols. These results indicated that the dose-enhancement ratio increased with increasing radiation dosage regardless of the protocol, with radiation delivered at the beginning of the combination treatment (RT-Tet protocol) causing greater cytotoxicity.

The effects of tetrandrine and early- or late-phase irradiation. Survival curves for the SH-SY5Y neuroblastoma cells treated with tetrandrine and early-phase irradiation (RT-Tet protocol; A and C), or late-phase irradiation (Tet-RT protocol; B and D). A and B, 24-h tetrandrine treatment; C and D, 48-h tetrandrine treatment. Cell viability was determined using the clonogenic assay. Each point is the average of three independent trials (nine determinations for each concentration) and presented as mean±standard deviation.
Isobologram analysis (Figures 3 and 4) demonstrated a mainly synergistic response for the RT-Tet protocol at the 10, 20, 30 and 40% neuroblastoma cell survival levels, in contrast, to a mainly subadditive response for the Tet-RT protocol. These results also indicated greater cytotoxicity for the RT-Tet than for the Tet-RT protocol and therefore the sequence dependence of this increase in efficacy.
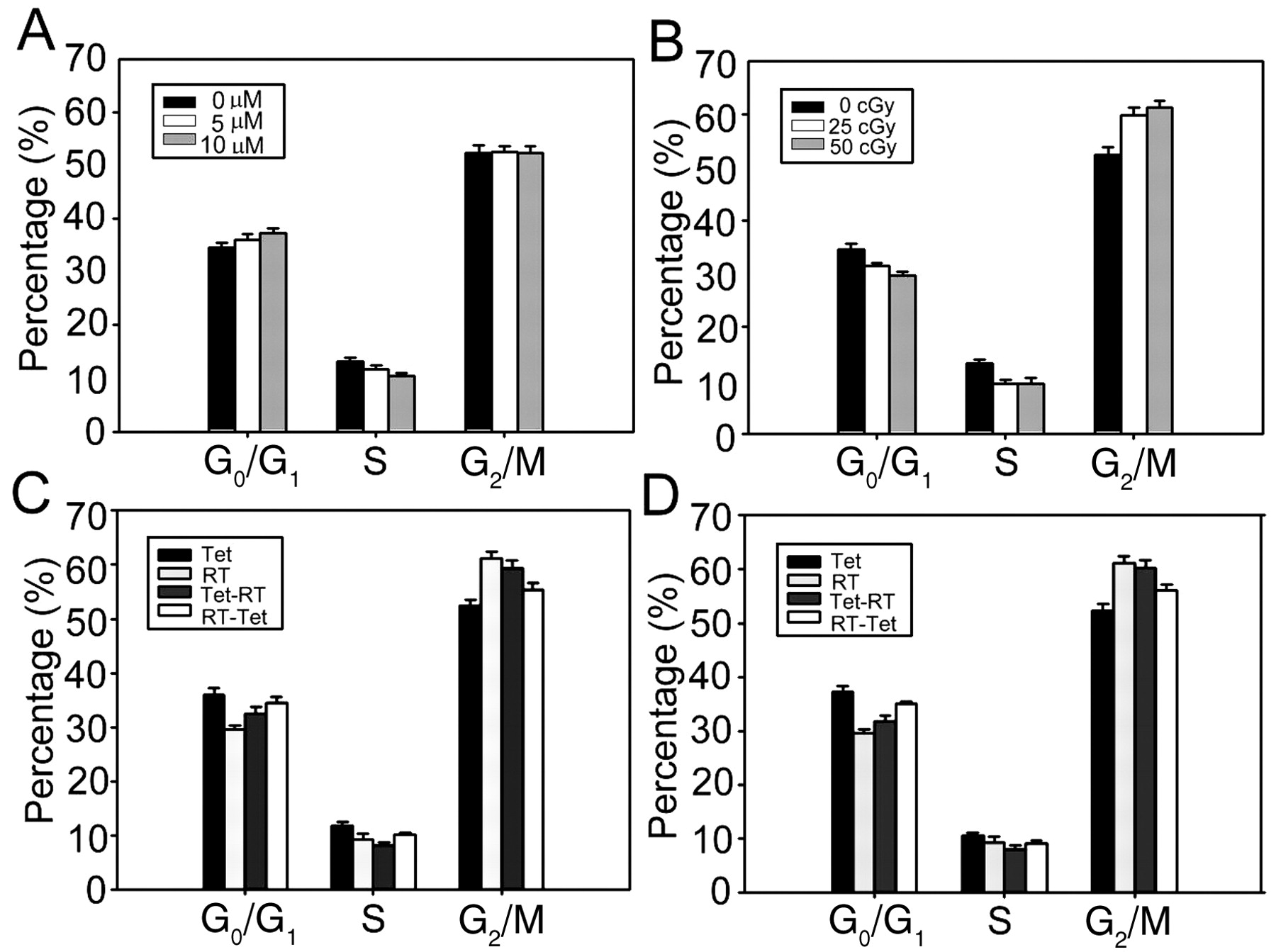
Cell cycle analysis. Tetrandrine treatment for 24 hours increased the fraction of cells in the G0/G1-phase (0 μM vs. 10 μM, p=0.02), reduced the fraction in the S-phase (0 μM vs. 10 μM, p=0.001) and had no effect on the fraction in the G2/M-phase (0 μM vs. 10 μM, p=0.48), as compared with the control (Figure 5A). Radiation reduced the fraction of cells in G0/G1 and S and increased the fraction in G2/M, compared with the control (0 cGy vs. 50 cGy, p<0.001) (Figure 5B).
The effects of tetrandrine (5 or 10 μM) combined with radiation (50 cGy), according to the RT-Tet or Tet-RT protocol, are shown in Figures 5C and 5D. The RT-Tet or Tet-RT protocol reduced the fraction of cells in G0/G1 and S and increased the fraction in G2/M, compared to treatment with tetrandrine alone (p<0.05), with these changes in cell cycle distribution being greater after Tet-RT treatment than RT-Tet treatment (p<0.05). On the other hand, as compared to radiation alone, RT-Tet significantly increased the fraction in G0/G1 and reduced the fraction of cells in G2/M (p<0.001). In contrast, Tet-RT only mildly increased the fraction of cells in G0/G1 (p<0.05) and did not change the fraction in G2/M (p>0.16), as compared to the radiation alone treatment.
Discussion
In this study, significant concentration- and time-dependent tetrandrine cytotoxicity was demonstrated for human SH-SY5Y neuroblastoma cells. The tetrandrine LC50 was 8.0 or 4.7 μM after 24 or 48 h of treatment, respectively, being comparable with previously reported LC50 values for tetrandrine against breast cancer, lung cancer, neuroblastoma, Burkitt's lymphoma, hepatoma and leukemia (5, 17-20, 30). Postoperative radiotherapy has been the standard treatment strategy for neuroblastoma (31) and tetrandrine was shown to induce apoptosis in several types of cancer cells (5, 8, 17, 19, 20) and neuroblastoma cells in this study. Thus, combining radiotherapy with tetrandrine might be a potential approach for increasing neuroblastoma cell radiosensitivity. An advantage of combination therapy is that it limits the nonspecific toxicity often associated with exceedingly high doses of a single treatment regimen (25, 26). A literature review revealed only two studies of the radiosensitization effect of tetrandrine (25, 26) and the optimal schedule of combining radiotherapy with tetrandrine has yet to be determined. In this study, a mainly synergistic effect of tetrandrine plus radiation when used in the RT-Tet protocol, while only a subadditive effect when used in the Tet-RT analog was found. Notably, a higher dose-enhancement ratio was revealed for the RT-Tet protocol. Furthermore, the increase in dose-enhancement ratio was more significant with the higher radiation dose (50 cGy), but not with longer drug exposure time. These results suggested that the RT-Tet protocol was more cytotoxic for SH-SY5Y neuroblastoma cells than the Tet-RT variant. The sequence-dependence of tetrandrine-radiation exposure suggested that the cytotoxicity was maximized when the radiation was delivered concomitantly at the beginning of tetrandrine treatment, which may have future application in the clinical treatment of neuroblastomas.

Isobologram analysis of the effects of tetrandrine for 24 hours and early- or late-phase irradiation on SH-SY5Y neuroblastoma cells. Early-phase irradiation (RT-Tet protocol, A, C, E and G), late-phase irradiation (Tet-RT protocol, B, D, F and H) (tetrandrine: 0, 5 or 10 μM; irradiation: 0, 25 or 50 cGy). The isobologram analyses took 10% (A and B), 20% (C and D), 30% (E and F) and 40% (G and H) survival as the end-points. The isoelectric mode I and mode II boundaries delineating the envelope of additivity were calculated from the response to radiation or drug alone. Data points (▴) were determined from the combination treatment experiments.

Isobologram analysis of the effects of tetrandrine for 48 hours and early- or late-phase irradiation on SH-SY5Y neuroblastoma cells. Early-phase irradiation (RT-Tet protocol, A, C, E and G), late-phase irradiation (Tet-RT protocol, B, D, F and H) (tetrandrine: 0, 5 or 10 μM; irradiation: 0, 25 or 50 cGy). The isobologram analyses took 10% (A and B), 20% (C and D), 30% (E and F) and 40% (G and H) survival as the end-points. The isoelectric mode I and mode II boundaries delineating the envelope of additivity were calculated from the response to radiation or drug alone. Data points (▴) were determined from the combination treatment experiments.

Cell cycle distribution of the SH-SY5Y neuroblastoma cells treated for 24 hours with tetrandrine and/or irradiation. (A) Tetrandrine alone (B) Irradiation alone (C) RT-Tet or Tet-RT treatment with 5 μM of tetrandrine (Tet) and 50 cGy irradiation (RT). (D) RT-Tet or Tet-RT treatment with 10 μM of tetrandrine and 50 cGy irradiation. Data from three independent experiments are shown as mean±standard deviation, representing the proportion of cells in the G0/G1-, S-, or G2/M-phases. Bars, standard deviation.
Although the precise mechanisms underlying the sequence dependence of tetrandrine radiosensitization remain elusive, the interaction between the drug's effect on the cell cycle and the cell's capacity to repair DNA damage after irradiation are probably involved in this mechanism (32). In the present study, tetrandrine caused accumulation of neuroblastoma cells in the G0/G1 phase, which was consistent with findings in most cell types so far studied (20, 33, 34). In contrast, irradiation arrested the neuroblastoma cells in the G2/M-phase of the cell cycle, which is a common finding in many cell types (31). Arrest of the cell cycle in the G2/M-phase promotes cell viability by allowing time for DNA repair prior to the entry into mitosis (35, 36) and causes delay in the induction of apoptosis because cells arrested in G2 have higher levels of DNA repair (37). A G2 delay may thus allow a proportion of DNA-damaged cells to survive and contribute to radioresistance (38-40). Therefore, disruption of this delay might reasonably be a mechanism for the induction of radiosensitization. In contrast to the Tet-RT protocol, RT-Tet significantly reduced the accumulation of cells in the G2/M-phase caused by irradiation and increased the fraction in G0/G1. Thus, the RT-Tet protocol (but not Tet-RT) could partially abrogate the radiation-induced G2/M arrest, which has previously been found to be related to tetrandrine-induced radiosensitization in nasopharyngeal carcinoma cells (26).
Since radiosensitization requires that cytotoxic quantities of a drug reach the target cells shortly before irradiation (29) and persist post-irradiation (to effectively impair DNA repair) (41), combining drug therapy with radiation at the beginning of drug treatment is a reasonable treatment schedule. By abolishing the G2 delay after irradiation, the subsequent tetrandrine treatment in the RT-Tet protocol could prevent the normally rapid repair of radiation-induced DNA damage and radiation resistance, resulting in a synergistic and stronger cytotoxic effect. In contrast, late-phase irradiation (Tet-RT protocol) failed to abolish the G2 delay because it was induced after tetrandrine was removed from the culture medium. Certainly, other mechanisms, such as the influence of cell-cycle regulators, apoptosis induction and doubling-time of tumor cells, might also contribute to radiosensitization by tetrandrine (39). Further studies are necessary to elucidate the underlying pathophysiology.
Acknowledgements
This study was supported by research grant FEMH-95-D-004 from Far Eastern Memorial Hospital, Taiwan, R.O.C., awarded to Dr. Yun Chen.
- Received March 6, 2009.
- Revision received June 2, 2009.
- Accepted June 17, 2009.
- Copyright© 2009 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.