


Abstract
Background: Plasmacytoid dendritic cells (PDCs) infiltrating solid tumor tissues and draining lymph nodes of head and neck squamous cell carcinoma (HNSCC) show an impaired immune response. Immunosuppressive cytokines secreted by HNSCC have been recognized to account for reduced interferon-α (IFN-α) production, whereas the influence on PDC migration has not yet been investigated. Materials and Methods: PDCs were isolated from human peripheral blood by magnetic bead separation. Cellular functions and characteristics were analyzed using flow cytometry, migration assays and ELISA techniques. Results: We show that HNSCC and CpG oligonucleotides controversially influence the migration and IFN-α production of PDCs. Furthermore we demonstrate that HNSCC induced migration towards stromal cell-derived factor-1 (SDF-1) and macrophage inflammatory protein 1 β (MIP-1β) is not dependent upon the surface density of chemokine (CXC motif) receptor 4 (CXCR4) and chemokine (C-C motif) receptor 5 (CCR5). Conclusion: We propose that HNSCC triggered PDC migration is most likely linked to a multilevel process strongly modulated by tumor microenvironmental influences.
Dendritic cells (DCs) are bone marrow-derived antigen-presenting cells, such as B-cells and monocytes, and appear to be indispensable for initiating appropriate immune responses. A complicated trafficking system leads them from the bone marrow through the bloodstream to distinct peripheral tissues. Different immature DC populations reveal unique kinds of chemokine responsiveness (1). Finally they migrate to lymphoid organs in order to present processed antigens to lymphocytes and to stimulate immune responses (2-6). Two main human dendritic cell subsets are distinguished: myeloid dendritic cells (MDCs) including Langerhans cells (LCs), dermal dendritic cells and interstitial DCs versus plasmacytoid dendritic cells (PDCs) (7). Referring to their plasma cell-like appearance PDCs were first identified as plasmacytoid monocytes/T-cells in T-cell areas of lymphoid organs and later also in peripheral blood and tonsils (8-12). PDCs are to a small extent capable of presenting antigens to T-cells, however, their main function is characterized by their ability to produce large amounts of interferon (IFN)-α, triggered by CpG oligonucleotides which consist of unmethylated CG dinucleotides embedded in a specific sequence context within bacterial DNA (13-17).
In contrast to MDCs, which first migrate into inflamed tissue and then approach draining lymph nodes (LNs) via afferent vessels, PDC directly enter LNs from peripheral blood via endothelial venules (18). In this context, the receptor proteins chemokine (CXC motif) receptor 3 (CXCR3), CXCR4 and chemokine (C-C motif) receptor 7 (CCR7) have been proposed to be of capital importance for transmigration, as their chemoattractants CXCL9, SDF-1 and CCL21 have been detected in human LNs (8, 19-21). CD40L-, interleukin 3 (IL-3)- and virus-induced maturation as well as long-term incubation with CpG-DNA lead to a down-regulation of CXCR3 and CXCR4 by contrast to an up-regulation of CCR7 (22-24).
It has been shown that human solid tumor tissues of head and neck squamous cell carcinoma (HNSCC) are infiltrated by PDCs, which seem to strongly compromise proper immune functions in this environment (25). Cells of head and neck cancer are known to develop molecular strategies to escape from efficient anti tumor immune responses. It is supposed that tumor production of various immunosuppressive mediators contributes to a massively deranged immune defence (26). The molecular mechanisms responsible for these immunomodulatory transformation processes and the biosynthesis of the immunosuppressive HNSCC microenvironment remain mostly unknown (27, 28). The tumor environment impairs the secretion of IFN-α in response to CpG stimulation, most likely because of lower expression levels of Toll-like receptor 9 (TLR9) (25) which recognizes CpG-oligonucleotides and thus stimulates the polyclonal activation of B lymphocytes as well as the secretion of proinflammatory cytokines by PDCs and macrophages (29-31).
In this work we investigated the effects of HNSCC and CpG oligonucleotides on PDC migration and IFN-α production.
Materials and Methods
Isolation of PDC. PDCs were isolated from human peripheral blood (PBMC, buffy coats) provided by the blood bank of the University Hospital Lübeck, Germany. Blood donors were from 18 to 65 years old healthy donors who were tested as being negative for allergies. Additional exclusion criteria were manifest infections during the previous 4 weeks, fever and medication of any kind. PBMCs were obtained from buffy coats by Ficoll-Hypaque density gradient centrifugation as described elsewhere (32). PDCs were isolated by magnetic bead separation using magnetic labeled anti-BDCA-4 antibodies (Miltenyi Biotec, Bergisch-Gladbach, Germany).
Cell cultures. Permanent HNSCC cell lines BHY (DSMZ Germany) (33) and PCI-1 (generously given to us by Prof. T. Whiteside, Pittburgh Cancer Institute, PA, USA) were cultured in Dublecco's modified Eagle's medium (DMEM; Gibco, NY, USA) supplemented with 10% fetal calf serum (FCS), 1 mM glutamine and 0.1 mM sodium pyruvate. All compounds were purchased as endotoxin-free. By several centrifugation steps, a cell-free supernatant solution was prepared. Oligodesoxynucleotides (ODNs) were provided by Metabion GmbH (Martinsried, Germany): ODN 2216 (CpG-A): 5′-ggGGGACGATCGTCgggggG-3′ (lowercase letters: phosphorothioate linkage; capital letters: phoshodiester linkage 3′ of the base; bold: CpG-dinucleotides). ODN 2243 (5′-ggGGGAGCATGCTCgggggG-3′) was used as an internal control. On the basis of previous studies, oligonucleotides were used at a final concentration of 3 μg/ml (34).
Flow cytometry. PDCs were identified as being CD123-PE positive, HLA-DR-PerCP positive, lineage cocktail-FITC (CD3, CD14, CD16, CD19, CD20, CD56; Becton Dickinson, Heidelberg, Germany) negative and BDCA2-APC (Miltenyi Biotec) positive cells. Surface antigen staining for migration markers was performed using antibodies CXCR4-PE-CY5, CCR5-APC, CXCR3-APC and CCR7-PE-CY7 (all Becton Dickinson) whilst APC isotype control, purified CCR7 and CXCR4 were used as negative controls (all Becton Dickinson). Early apoptotic or dead cells were detected by staining the cells with Annexin-V-FITC and Propidium-Iodide-PE (Becton Dickinson). A total of 2×105 PDCs were incubated on ice in 50 μl 0.5% bovine serum albumin (BSA) in phosphate buffered saline (PBS) with 2 μl of each antibody for 25 minutes. The PDCs were washed in 500 μl 0.5% BSA and subsequently analyzed on a FACSCanto (Becton Dickinson) equipped with FACS DIVA software.
Migration analysis. PDCs (4.8×105) were incubated with HNSCC supernatants and/or CpG-ODN, respectively. Cells were subjected to migration analysis using a ChemoTx system (NeuroProbe Inc., Gaithersburg, MD, USA), which consists of a specially designed 96-well microplate and a framed filter with the filter membrane bonded to a rigid frame. The microplate provides bottom wells for chemoattractants and other reagents in cell migration assays. No top wells are required because the proprietary framed filter is coated with a hydrophobic mask that confines each cell suspension sample to its site on top of the filter. The 96 sites on the filter correspond to the 96 wells in the microplate. Cell suspension is pipetted directly onto the sites on the top side of the filter and remains in hemispherical drops during incubation. Relative numbers of migrated cells were determined using an MTT assay (In Vitro Toxicology Assay Kit, MTT-based; Sigma®-Aldrich Co., Steinheim, Germany). The MTT assay is based on the cleavage of the yellow tetrazolium salt MTT into purple formazan by metabolically active cells, which can be quantified photometrically.
Cytokine analysis. An IFN-α module set from Bender MedSystems (Vienna, Austria) was used to detect IFN-α in cell culture supernatants according to the instructions provided by the manufacturer. The Bio-Plex Cytokine Assay (Bio-Rad, Munich, Germany) was used for the determinations of various cytokines such as IL-2, IL-4, IL-5, IL-8, IL-10, IL-12, IL-13, granulocyte macrophage colony-stimulating factor (GM-CSF), IFN-y, vascular endothelial growth factor (VEGF), prostaglandin E2 (PGE2) or tumor-necrosis factor-α (TNF-α). These cytokine assays allow the multiplexed quantitative measurement of multiple cytokines in a single small volume of cell culture supernatant. The protein array was analyzed by a dedicated microplate reader system (Bio-Plex Array Reader; Bio-Rad) and the data were calculated by the Bio-Plex Manager software.
Statistical analysis. Statistical significance was determined by paired two-tailed Students t-test. Significant data were defined as resulting in a p<0.05. At least ten experiments were performed per stimulus.
Results
Cytokine analysis of HNSCC cell supernatant. Cytokine analysis of the used HNSCC supernatants revealed the presence of various cytokines such as IL-6, IL-8, IL-10, GM-CSF, VEGF, and PGE2, which are believed to play a critical role in the tumorigenesis and maintenance of head and neck cancer (26, 35-39). Table I gives an overview of the detected cytokines and their proposed functions. Incubation of isolated PDCs with PGE2, which is known to interfere with monocyte functions such as migration and adherence to endothelial cells (36, 40), did not significantly affect PDC migration (Figure 1).
PDC migration and IFN-α production in response to HNSCC and CpG ODN. It has been shown that PDCs infiltrate tumor tissue and draining LNs of patients with head and neck squamous cell carcinoma. The ability of these PDCs to produce IFN-α in response to CpG motif containing oligonucleotides was significantly impaired (25). Hence, we investigated the influence of HNSCC and CpG oligonucleotides on PDC migration against SDF-1 which is present in human HEV (19) and has been implicated in metastasis of cancer (41). In addition, we analyzed PDC migration against MIP-1β which is expressed primarily by T-cells, B-cells, and monocytes after antigen or mitogen stimulation.
Cytokines present in the supernatant of permanent HNSCC cell lines and their proposed sites of action.
PDCs were isolated and incubated with fresh supernatants of different permanent HNSCC cell lines, CpG2216, CpG2243 and standard culture medium without any additional effectors (control) for 12 hours. In order to estimate early and late effects of CpG2216 on HNSCC-stimulated cells. PDCs were also incubated with HNSCC supernatants for 12 hours adding CpG2216 after 10 hours (late effect) and similarly by coincubation for 2 hours (early effect). Our investigations revealed a significantly increased PDC migration towards both SDF-1 and MIP-1β in response to 12 hours of incubation with HNSCC supernatants, with no differences between the effects of both stimuli (Figure 1).
Our data indicate that the stimulation of PDCs with CpG2216 led to strongly reduced migration levels compared to the medium control (Figure 1). These contrasting effects of HNSCC and CpG2216 on PDC migration were clearly corroborated by incubating PDCs simultaneously with both effectors, with HNSCC supernatant mitigating the reduction in migration caused by CpG2246.
Our data reveal a contrasting influence of HNSCC and CpG oligonucleotides on PDC migration and IFN-α production. HNSCC supernatants strongly increased migration towards SDF-1 whilst cells incubated with CpG2216 and CpG2243 showed impaired migration behaviour. Only CpG2216 stimulation caused potent IFN-α production. Double stimulation revealed both a decreased migration and IFN-α production when compared to the influence of single stimuli. Applying CpG2216 after 2 hours caused a greater impairment of migration than of IFN-α production; applying CpG2216 after 10 hours led to the converse (Figure 1).
Analyzes of PDC chemokine receptors. In a further step, we investigated the SDF-1 receptor CXCR4 and the MIP-1β receptor CCR5 by flow cytometric analysis under identical conditions as above. Surprisingly, the median fluorescence (MF) of CXCR4 was not significantly (p>0.05) affected after incubation with HNSCC supernatants but was highly up-regulated (p<0.05) after early CpG2216 influence. The effect of CpG2216, CpG2243 and late stimulation with CPG2216 resulted in reduced (p<0.05) CXCR4 expression as expected (Figure 2A).
The MF of CCR5 revealed impaired (p<0.005) signalling after incubation with HNSCC and no significant influences after early CpG2216 stimulation. Similary to the effect on CXCR4, CpG2216, Gpc2243 and late CpG2216 stimulation caused a down-regulation of CCR5 (Figure 2B).
It has been demonstrated that some migration receptors have the ability to modulate the sensitivity of others against their attractant (8, 23). Thus we also measured the MF of CXCR3 and CCR7 under the same conditions. Just as shown for CCR5, the MF of CXCR3 was reduced (p<0.05) after incubation with HNSCC supernatants, CpG2216 and CpG2216 after 10 hours. The other stimuli did not evoke a significant change (Figure 2C).
Astonishingly, only the MF of CCR7 was up-regulated (p<0.05) after incubation with HNSCC supernatants. Early and late CpG2216 stimulation increased CCR7 expression whilst CpG2243 was down-regulated CCR7. CpG2216 did not show a significant effect (Figure 2D).
Discussion
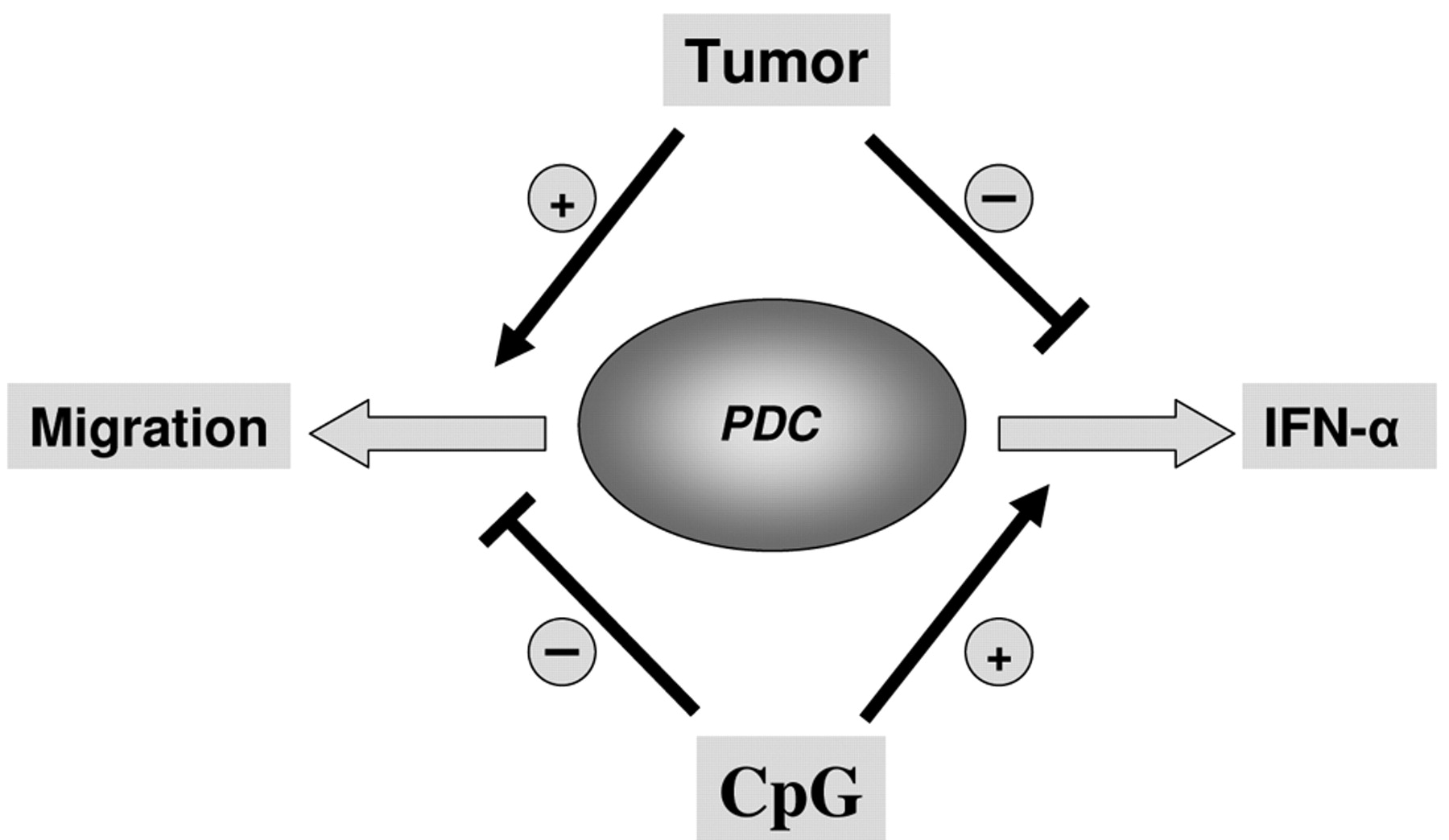
Numerous immunomodulating cytokines within the HNSCC microenvironment are responsible for strongly impaired immune functions, which often allow human tumors to escape from anti tumor immune responses (27, 35). Solid HNSCC have been found to be infiltrated by various species of immune cells, which are believed to participate in the formation of the HNSCC microenvironment because of functional alterations (25, 26, 42, 43). In this work we demonstrate that human PDCs are functionally affected by the microenvironment of HNSCC resulting in an increased migration activity and an impaired production of IFN-α in response to CpG-ODN (see model in Figure 3).
It has been demonstrated that freshly isolated circulating PDCs do not migrate towards inflammatory chemokines although they display low levels of CXC and CC receptors (24, 44). Contrarily to our expectations we found that PDCs strongly migrated towards SDF-1 and MIP-1β subsequent to incubation with HNSCC supernatants for 12 hours, even though the MF levels of CCR5 (p<0.05) and CXCR4 (p>0.05) were down-regulated. We propose that HNSCC-induced migration is most likely linked to a multi level process modulated by tumor microenvironmental influences. We found that CXCR4 was highly up-regulated after double-incubation with HNSCC supernatants and CpG2216 for 2 hours whilst migration activity hardly differed from that of the medium control. On the one hand, we assume a time-dependent latency between temporary phenotypical activation after 2 hours and persisting functional activation of CXCR4/CCR5 after 12 hours. On the other hand, we consider a CXCR4/CCR5-independent impairment of migration induced by CpG2216, since migration also decreased compared to single supernatant stimuli, when adding CpG2216 after 10 hours. We suggest that CCR5 is up-regulated in an earlier or later time-frame, as the MF was not significantly elevated after 2 hours. Possibly CpG2216 interferes with receptor-sensitizing mechanisms such as Ly49D-L in rodent natural killer (NK) cells or soluble CXCR3 ligands which have been shown to influence the threshold of sensitivity to SDF-1 and may have similar effects on CCR5/MIP1β in HNSCC (8, 23, 45). However CXCR3 was also significantly down-regulated after incubation with tumor supernatants. Furthermore prostaglangin (PG)E2 which is essentially implicated in monocyte and blood DC migration (46-48) has been demonstrated to be secreted by HNSCC cells (49) and could have an analogous influence on tumor infiltrating PDCs. Tryptophan metabolism regulated by the enzyme indoleamine 2, 3-dioxygenase (IDO; IDO is strongly expressed by HNSCC, our unpublished data) and which has been associated with modulation of CCR5 and CXCR4, as well as DC responsiveness to chemokines (50), could comprise a further target for CpG2216. Moreover, autocrine mechanisms such as the nuclear protein high mobility group box 1 (HMGB1) which regulates the expression of CXC receptors in MDCs (51) have yet to be investigated in HNSCC-induced migration. Down-regulation of CXCR3, CXCR4, CCR5 and impaired migration towards SDF-1 after incubation with CpG2216 reflects PDC behaviour after viral stimulation as reported before (23, 52). We show for the first time that CpG2243 also significantly decreases the fluorescence levels of CXCR4 and CCR5 implicating a decreas in migration activity. It seems to be conflicting that the receptors for inflammatory chemokines (CXCR3, CXCR4, CCR5) are down-regulated whilst CCR7 is up-regulated after stimulation with HNSCC supernatants, similary to the reaction triggered in response to pathogenic stimuli and CpG2243 (19, 23, 24, 53). However it has recently been shown that the CCR7 pathway is not only used for the induction of protective immunity but also for the establishment of immunological tolerance (54). Furthermore the expression of CCR7 promoted lymph node metastasis of a human non-small cell lung cancer line Lu-99 (55).
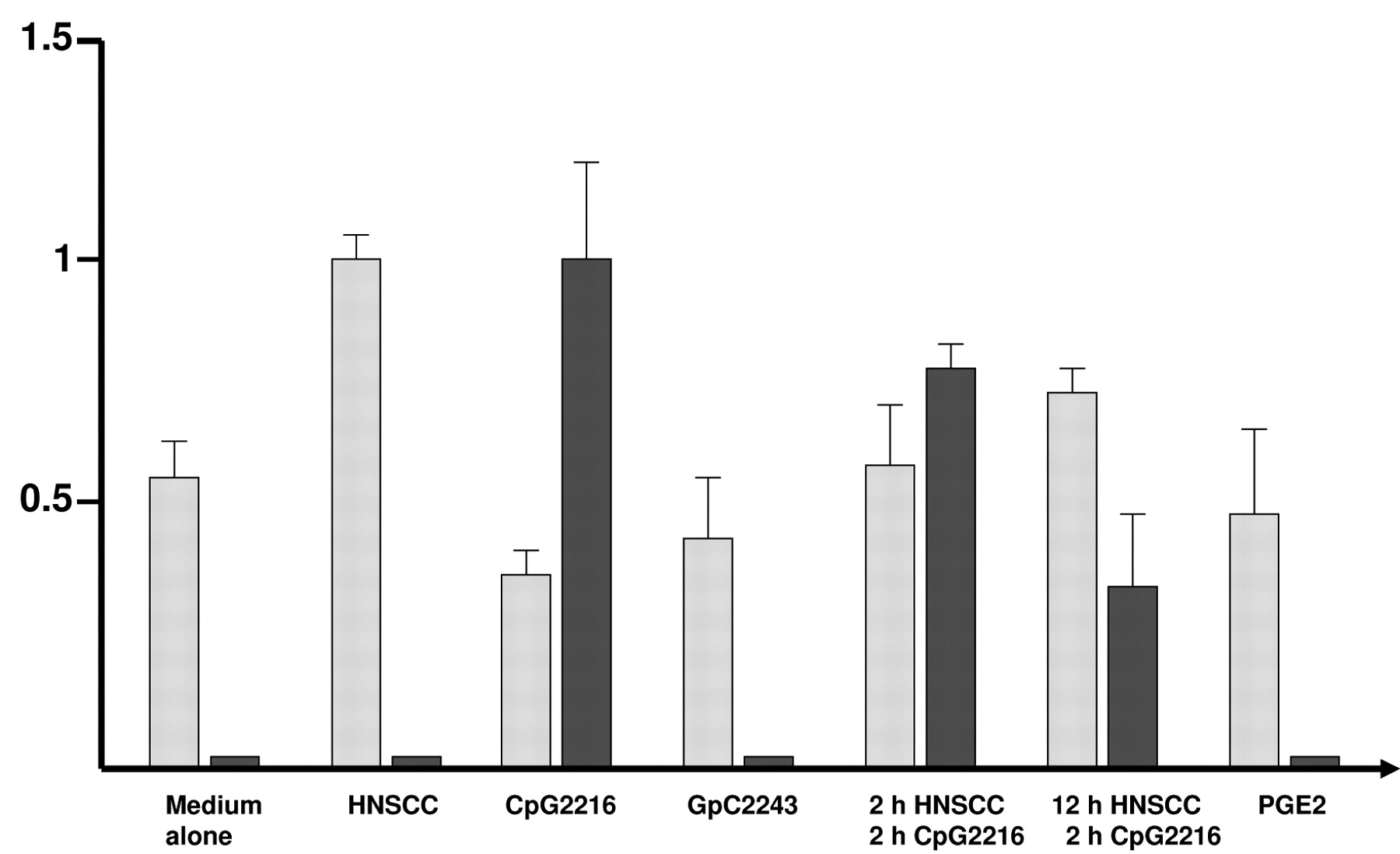
Migration and IFN-α production of human PDC. Increased migration of PDCs was observed in response to HNSCC supernatants. CpG stimulation of PDCs resulted in a reduced cellular migration, whereas IFN-α production was strongly activated. Inhibitory GpC2243 was used as an internal negative control. Simultaneous treatment with CpG2216 and HNSCC supernatants demonstrated the contradictory effects of both stimuli. Levels of cellular migration activity and IFN-α secretion are illustrated with dark and light bars, respectively. Experiments were performed at least three times for each data point.

Analysis of PDC chemokine receptors. Chemokine receptors CXCR4 (A), CCR5 (B), CXCR3 (C), and CCR7 (D) were analyzed by flow cytomety under conditions as indicated. The results of median fluorescence (MF) were calculated from at least ten independent experiments.

Migration and IFN-α secretion of human PDCs are contradictory affected by HNSCC supernatants and CpG2216.
We propose that HNSCC-induced PDC migration is a complex process which is presumably strongly influenced by the tumor microenvironment and is not dependent upon migration receptor density. After tumor exposure, PDCs assumingly migrate into draining LNs in a SDF-1, MIP-1β and particularly CCL19/21 mediated manner, facilitating metastasis by inducing tolerance. This is underlined by the fact that the presence of CCL19 in breast cancer has been correlated with prolonged overall survival (56). Local tumor chemokines may well prevent the facilitation of metastasis by inhibiting migration into draining LNs.
The precise cellular functions of the different HNSCC cytokines within the process of immunosuppression and immunomodulation are yet mostly unclear and have to be further elucidated. The understanding of these regulatory routes is an essential step for the development of novel immunotherapeutic strategies against HNSCC.
Acknowledgements
We thank Nicole Bohnert for skilful support in some parts of this work. We are grateful to all members of the Department of Otorhinolaryngology for helpful discussions and a comfortable atmosphere. This work was supported by grants of the Mildred Scheel Stiftung (Deutsche Krebshilfe), the Possehl-Stiftung, the Werner and Klara Kreitz Stiftung, the Monika Kutzner Stiftung and the Rudolf Bartling Stiftung.
Footnotes
-
↵* Both authors contributed equally to this work.
- Received April 10, 2009.
- Revision received June 3, 2009.
- Accepted June 16, 2009.
- Copyright© 2009 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved





{kind=link}
{kind=link}
{kind=link}