


Abstract
Background: To identify immunosuppressive elements present in ovarian cancer associated ascites. Patients and Methods: Ascites and plasma were obtained from ovarian, primary peritoneal or fallopian tube cancer patients. Surface markers were identified by fluorescence-activated cell sorting (FACS). Cytokine and chemokine concentrations were measured with LINCOplex microarrays. Antigen-specific T-lymphocytes from ascites and plasma were expanded with artificial antigen-presenting cells (aAPC). Cell-mediated immune response was assessed with chromium release assays. Results: Samples were collected from 37 patients with advanced ovarian cancer. FACS was performed on 27 ascites specimens. A low CD4/CD8 ratio (<1.6) was seen in 13 patient samples and associated with significantly improved overall survival (p=0.040). LINCOplex evaluation of 22 paired ascites and plasma samples demonstrated significantly elevated levels of IL-6, IL-8, IL-10, IL-15, IP-10, MCP-1, MIP-1β and VEGF and significantly reduced levels of IL-2, IL-5, IL-7, IL-17, PDGF-BB, and RANTES in ascites compared to plasma (p<0.05). Autologous ovarian cancer cell lysis with T-lymphocytes from ascites was limited. Although aAPC stimulation resulted in effective expansion of antigen specific T-cells from peripheral lymphocytes (35-fold), only limited expansion was noted from ascites-derived lymphocytes (10-fold). Conclusion: Ovarian cancer-associated ascites may provide an immunosuppressive environment. A high CD4/CD8 ratio, which may indicate the presence of regulatory T-cells, is associated with poor outcome. Reduced IL-2 and elevated IL-6 and IL-10 levels favor a Th2 inhibitory immune response. This immunosuppressive climate may explain our observation of non responsiveness in ascites derived T-cells.
Abbreviations: aAPC, Artificial antigen-presenting cells; bFGF, basic fibroblast growth factor; CMV, cytomegalovirus; CTL, cytotoxic T lymphocytes; Ep CAM, epithelial cell adhesion molecule; FACS, fluorescence-activated cell sorting; G-CSF, granulocyte colony-stimulating factor; GM-CSF, granulocyte-macrophage colony-stimulating factor; Her2/neu, human epidermal growth factor receptor 2; IL, interleukin; INF-γ, interferon γ; IP-10, induced protein 10; MART-1, melanoma antigen recognized by T-cells; MCP-1, monocyte chemotactic protein-1; MIP-1α, macrophage inflammatory protein-1α; MIP-1β, macrophage inflammatory protein-1β; MUC 1, mucin 1; PBL, peripheral blood lymphocytes; PBMC, peripheral blood mononuclear cells; PDGF-BB, platelet-derived growth factor-BB; RANTES, regulated upon activation, normal T-cell expressed, and secreted; TCGF, T-cell growth factor; TNF-α, tumor necrosis factor-α; TAL, tumor-associated lymphocytes; TIL, tumor-infiltrating lymphocytes; VEGF, vascular endothelial growth factor.
In the United States, ovarian cancer ranks fifth as a cause of cancer-related deaths among females (1). Unfortunately, the majority of cases are diagnosed at an advanced stage. Despite optimal cytoreductive surgery and effective primary chemotherapy, five-year survival rates of advance ovarian cancer remain less than 30%. Novel approaches to the treatment of ovarian cancer are necessary in order to improve survival.
Given the presence of mutations and chromosomal abnormalities, virtually all malignancies should present novel antigens capable of immune recognition. The development of a clinically apparent cancer suggests either an evasion of functional host immune responses or creation of an immunosuppressive environment. Potentially, the host immune response could be exploited to generate an anti-tumor response. Several studies suggest not only the capacity to enhance immune response against malignancy, but also the possibility of improved outcome. Rosenberg and colleagues (2) successfully expanded autologous tumor-infiltrating lymphocytes (TIL), ex vivo, and re-infused them into melanoma patients together with the cytokine, IL-2. Re-infused TIL trafficked back to tumor sites and directly induced tumor shrinkage. Continued refinements to adoptive immunotherapy have resulted in increased success rates for treatment of melanoma patients (3). Encouraging results with adoptive immunotherapy have also been reported in pancreatic and gastric cancer (4, 5).
Manipulation of the immune system represents an active area of investigation in the development of ovarian cancer therapy. Multiple attractive ovarian cancer-associated target antigens for immunotherapy have been identified; these include CA125, epithelial cell adhesion molecule (Ep CAM), mesothelin, human epidermal growth factor receptor 2 (her2/neu), and mucin 1 (MUC 1) (6-10). Additionally, tumor-specific cytotoxic T lymphocytes (CTL) have been generated in vitro and are capable of recognition and lysis of ovarian cancer cells (11, 12). Immune response appears to be clinically relevant in ovarian cancer; Zhang and colleagues (13) demonstrated that the presence of TIL in ovarian tumor specimens correlated with improved clinical outcome.
Despite evidence of viable ovarian cancer targets for immune response and an association of effector T-cells with improved survival, successful clinical trials for immunotherapy for ovarian cancer have been limited (14, 15) and have not been replicated in larger populations. Ovarian malignancies may alter the local environment resulting in a suppression of the immune system. These changes may be reflected in the large volume peritoneal fluid (ascites) associated with ovarian cancer. In addition to tumor cells, this fluid contains cellular components of the immune system such as lymphocytes and regulatory factors such as cytokines and chemokines. As such, ovarian cancer ascites provides a unique opportunity to evaluate tumor microenvironment and potential effect on anti-tumor immune response. The goal of this investigation is to identify immunosuppressive factors and cells present in ovarian cancer-associated ascites. Specifically, we analyzed ovarian cancer-associated ascites and matched plasma samples for the presence and concentration of immune stimulatory and regulatory factors. We identified alterations in interleukin expression in ovarian cancer associated ascites consistent with a Th2 (inhibitory) effect. We evaluated the composition of the T lymphocyte population in ovarian cancer-associated ascites. We found an elevated CD4/CD8 ratio to be associated with poor outcome. We further compared the immune response of T-cells isolated from either the peripheral blood or the ovarian cancer ascites against non-ovarian cancer antigens such as melanoma antigen recognized by T-cells (MART-1) and cytomegalovirus (CMV). Importantly we found that in several patients the T-cell response against these non-ovarian antigens was impaired in T-cells isolated from the ascites fluid.
Patients and Methods
Patients. The Institutional Review Board of Johns Hopkins Medicine approved this investigation. Patients undergoing surgery for suspected ovarian cancer were offered enrollment in the investigation. All donors gave written informed consent before entry into the study. Plasma and ascites was collected from patients undergoing cytoreductive surgery for ovarian cancer. In addition, plasma was obtained from normal subjects. Medical records were retrospectively reviewed and relevant clinical and pathology data extracted. The time of diagnosis was considered to be the date of the primary surgical procedure. Time from diagnosis to death or last follow up was calculated. Median follow-up for survivors was calculated for subjects not censored for death.
Peptides. Peptides used in this study were prepared by the Johns Hopkins University core facility. HLA-A2 restricted peptides from MART-1 and CMV were generated (16). HLA-A2 restricted peptides from ovarian cancer-associated target antigen mesothelin, Ep CAM, her2/neu, and MUC 1 have been described and were prepared (17-20). In addition, three HLA-A2 restricted peptides from ovarian cancer cell lines have been demonstrated to generate an antigen-restricted CTL response (11). The purity (>98%) of each peptide was confirmed by mass-spectral analysis and high pressure liquid chromatography.
Cell cultures from ovarian cancer-associated ascites. Ovarian cancer cells were isolated from ovarian cancer-associated ascites. Ascitic fluid was centrifuged at 800 × g for 15 min to pellet the cellular component. The pellet was suspended in M' medium supplemented with 5% human AB-serum (21). Adherent cells were harvested and evaluated for the presence of ovarian cancer cells by staining for appropriate markers, vimentin, CA 125, Ep-CAM. Alternatively, the cell pellet was resuspended and lymphocytes were isolated by Ficoll-Hypaque density gradient centrifugation.
Plasma and peripheral blood lymphocytes (PBL). Peripheral blood samples were obtained in heparinized tubes from healthy individuals and from ovarian cancer patients prior to primary cytoreductive surgery. The specimens were centrifuged. The plasma was collected and stored. The cell pellet was resuspended and PBL were isolated by Ficoll-Hypaque density gradient centrifugation.
Analysis of lymphocyte surface markers by flow cytometry. The surface marker analysis of lymphocytes was performed using fluorescence-activated cell sorter (FACS) (Becton Dickinson) and the FCS-Express software (DeNovo). We used the following mAbs conjugated to fluorescein isothiocyanate (FITC) or phycoerythrin (PE) for direct fluorescence: anti-CD3, anti-CD4, anti-CD8, anti-CD107α, anti-CD45RO (all from Becton Dickinson). Isotype-matched FITC- or PE-conjugated monoclonal antibodies were used as appropriate controls.
Multiplex cytokine microarrays. Cytokine and chemokine concentrations in matched plasma and ascitic fluid samples were determined using the LINCOplex Assay. This multiplex assay is capable of simultaneously quantifying multiple cytokines and chemokines. The Assay evaluated for the presence of IL-1β, IL-1ra, IL-2, IL-4, IL-5, IL-6, IL-7, IL-8, IL-9, IL-10, IL-12, IL-13, IL-15, IL-17, basic fibroblast growth factor (bFGF), Eotaxin, granulocyte colony-stimulating factor (G-CSF), granulocyte-macrophage colony-stimulating factor (GM-CSF), interferon-γ (INF-γ), induced protein 10 (IP-10), monocyte chemotactic protein-1 (MCP-1), macrophage inflammatory protein-1α (MIP-1α), macrophage inflammatory protein-1β (MIP-1β), platelet-derived growth factor-BB (PDGF-BB), regulated upon activation, normal T-cell expressed, and secreted (RANTES), tumor necrosis factor-α (TNF-α), and vascular endothelial growth factor (VEGF). Briefly, antibody specific to each cytokine is covalently coupled to Luminex microspheres. Each specific antibody is linked to a different microsphere labeled with a unique flouorescent dye. Samples, standards, and controls (25 μl) are incubated with the antibody coupled microspheres in 96-well microtiter filter plates for 1 hour at room temperature. After the plate is washed, detection antibody, a mixture of antibodies against the individual cytokines, is added. The plate is then incubated for 30 min. Streptavidinphycoerythrin is added and the plate is incubated for an additional 30 min. The plate is again washed and the beads are resuspended and read on the Luminex instrument. All specimens were run in duplicate and results are reported as means (22).
Preparation of artificial antigen presenting-cells (aAPC). aAPC were generated by coupling HLA-Ig and anti-CD28 onto microbeads, (Dynal, Lake Success, NY, USA). Briefly, beads were washed twice in sterile 0.1 M borate buffer. The beads were incubated with a 1 to 1 mixture of the HLA-A2-Ig and the anti-CD28 specific monoclonal antibody 9.3 in borate buffer for 24 h at 4°C on a rotator, washed twice with bead wash buffer, and after another 24 h incubation at 4°C in bead wash buffer, the buffer was replaced. Resulting aAPC were found to have 0.9×105 molecules of A2-Ig/bead and 1.9×105 anti-CD28 molecules/bead. For peptide loading, HLA-Ig-coated aAPC were washed twice with PBS and adjusted to 107 beads/ml in 30 μg/ml of peptide, final concentration. aAPC beads were stored in the peptide solution at 4°C (23).
Induction and expansion of antigen-specific T-cells. CD8+ T-cells, enriched by negative selection (removal of CD8- T-cells such as T helper lymphocytes, B-cells, monocytes) with magnetic sorting columns follow by positive selection with a magnetic cell sorting columns (Miltenyi Biotec), were co-cultured with peptide-pulsed aAPC or autologous cultured ovarian cancer cells, in a 96-well-round-bottom plate in 200 μl M'/well supplemented with 5% plasma and 1.7% T-cell growth factor (TCGF). On day 7, T-cells were harvested, counted, and with fresh stimulus replated. Restimulations were performed once a week. After 3-4 weeks of culture, we characterized the CD8+ T-cells for their phenotype by staining with appropriate tetrameric peptide-HLA-A*0201 complexes (24) and then analyzing with FACS. In addition, we performed intracellular staining (ICS) and CD107α analysis against peptide-loaded target cells as well as target cells expressing endogenous antigens, to verify their specificity and functionality (25).
Chromium release assay. The cytotoxic activity of T-cell lines was measured by a conventional 4-h 51Cr release assay using triplicate cultures in V-bottomed plates. (26) The target cells analyzed included HLA-A2+ and HLA-A2- cultured ovarian cancer cells, and peptide pulsed and unpulsed T2 cells. E:T ratios were 25:1, 5:1, and 1:1 on 2,000 target cells/well. Triplicate wells were averaged and percentage specific cytotoxicity was calculated as [(sample ± spontaneous release)/(maximum release ± spontaneous release)]. For spontaneous release, targets were plated without T-cells in M0 medium plus 5% autologous plasma. For maximum release, targets were plated with 5% Nonidet P-40 (IGEPAL; Sigma) detergent.
Intracellular detection of IL-4 and IFN-γ. For intracellular cytokine detection 106 effector cells were stimulated for 5 h at 37°C with 105 T2 cells pulsed with the appropriate peptide (30 μg/ml), supplemented by 0.5 ng/ml phorbol 12-myristate 13-acetate (PMA) and 4 ng/ml ionomycin. (27) Control experiments revealed that low doses of PMA and ionomycin did not induce secretion of IFN-g or IL-4 in the effector cells. Monensin (PharMingen) was added during the last 4 h. Cells were transferred to FACS tubes, surface stained with anti-CD8-APC mAb (Becton Dickinson), fixed and permeabilized with 200 μl Cytofix/Cytoperm Solution (PharMingen) for 20 min at 4°C. Intracellular staining was performed with PE-labeled mAbs against IFN-γ (PharMingen) and IL-4 (Becton Dickinson). The samples were analyzed using FACS.
Statistical analysis. Survival was calculated from time of surgery. Patient data were otherwise censored at the time of latest contact. Survival curves were generated using the Kaplan-Meier method and compared using the log-rank test. (28) Ascites and plasma was assayed for individual cytokine and chemokine concentrations as described above. The assays were run in duplicate and results were calculated as the average of these 2 values. For each cytokine and chemokine, the mean and standard error of the mean (SEM) was calculated for the 22 ascites specimens and for the 22 plasma samples. The ascites and plasma levels for the individual cytokines and chemokines were then compared using Student's t-test. P<0.05 was considered significant.
Results
Patient specimens. Ascites and plasma was collected from 37 patients with ovarian, fallopian tube or primary peritoneal cancer. Median age at time of cytoreductive surgery was 58.5 years (range 36.6-93.0 years). The majority of patients had advanced disease including 31 patients (84%) with stage IIIc disease and 5 patients (14%) with stage IV disease. One patient (3%) was stage Ic. Specimens were collected at the time of primary surgery in 95% (35/37) of cases. In the 5% (2/37) of cases, collection of specimens occurred during surgery for recurrent disease. Both patients were >6 months from chemotherapy with no clinical evidence of immunosuppression. Pathology was consistent with ovarian, fallopian tube and primary peritoneal cancer in 84% (31/37), 8% (3/37) and 8% (3/37) of cases respectively. High-grade (poorly differentiated) malignancies were noted in 89% (33/37) of patients. Moderately differentiated and well-differentiated tumors were reported in 3% (1/37) and 8% (3/37) of cases respectively. Serous histology was reported in 78% (29/37) of tumors. The remaining specimens included 2 endometrioid carcinomas (5%), 1 malignant mixed mesodermal tumor (3%), 2 mixed serous/clear cell carcinomas (5%), 2 mixed serous/ endometrioid carcinomas (5%), and 1 adenocarcinoma not otherwise specified (3%). Specimens were available for FACS analysis to determine the composition of the lymphocyte population and for Lincoplex, multiplex cytokine and chemokine microarray, analysis. Specimens from 12 patients (32%) were available for analysis by both FACS and Lincoplex (Table I).
Ovarian Cancer Specimen Analysis
CD-3+ T lymphocytes. FACS analysis was performed on unprocessed ascites from 27 patients. All specimens demonstrated a population of CD-3+ lymphocytes. In 78% (21/27) of ascites specimens, CD4+ T-cells comprised >50% of CD3+ lymphocyte population. Only 22% (6/28) of specimens demonstrated an excess of CD8+ lymphocytes. At Johns Hopkins Medical Institutions, plasma CD4/CD8 ratios between 1.6 and 2.2 are considered normal. Only 11% (3/27) of ascites specimens had a CD4/CD8 ratio within this range. The CD4/CD8 ratio was below 1.6 in 48% (13/27) of specimens and above 2.2 in 44% (12/27) of specimens.
Univariate analysis. Anti-tumor cell mediated immune responses are influenced by both CD8+ cells, which directly lyse cells, and CD4+ cells, which can function as helpers cells, augmenting response or function as regulatory cells, suppressing response. A Kaplan-Meier curve was generated to determine the influence of CD4/CD8 ratio on overall survival. All 27 patients had stage IIIc or IV disease. The majority had primary disease, with only 7% (2/27) of patients having recurrent disease. As mentioned, both of these patients were over 6 months from chemotherapy and had no evidence of immunosuppression. CD4/CD8 ratio below 1.6 was associated with significantly improved overall survival (p=0.040) (Figure 1).
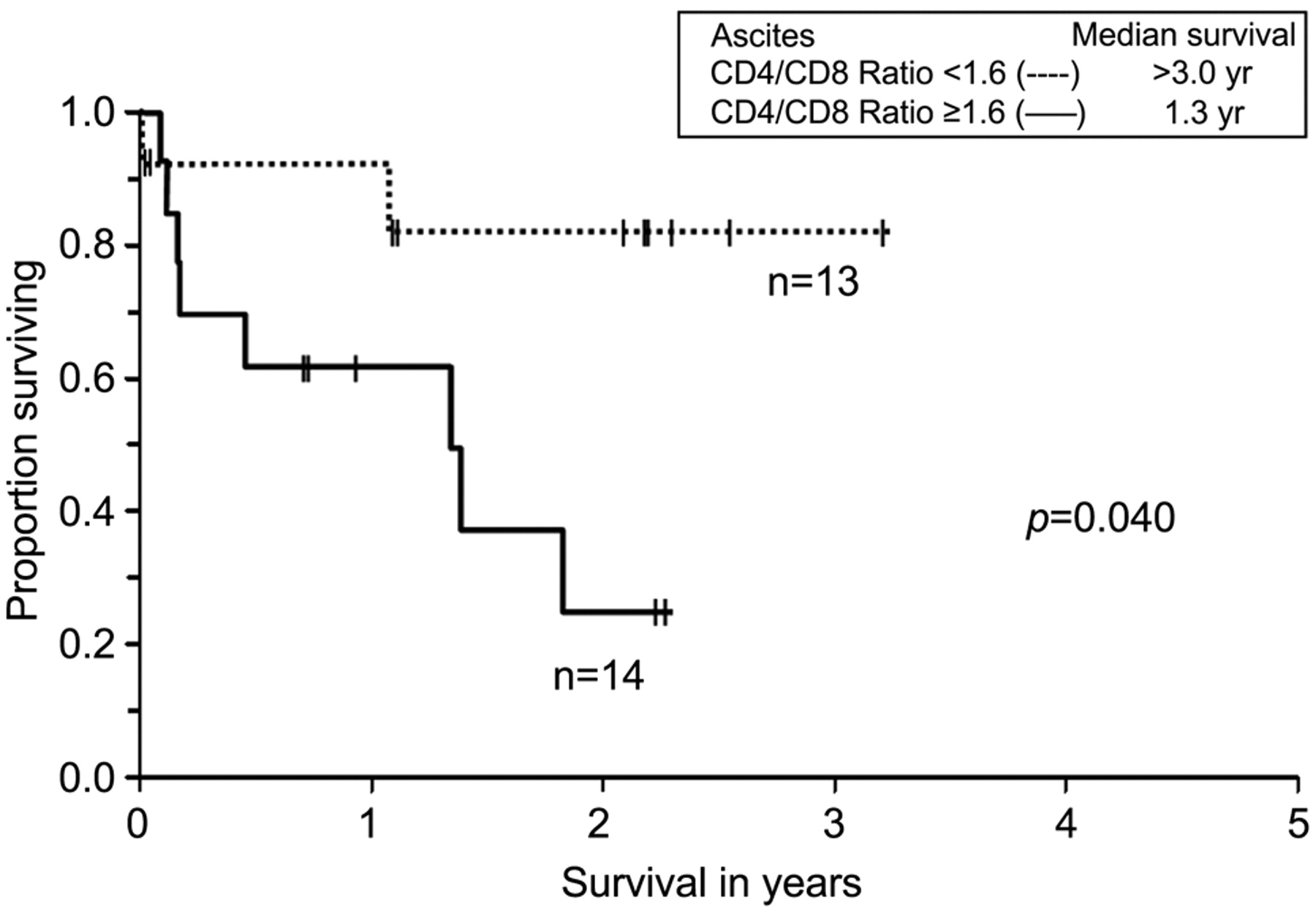
Kaplan-Meier analysis of overall survival for patients with ovarian cancer, fallopian tube or primary peritoneal cancer. The x-axis indicates overall survival in years after cytoreductive surgery. The y-axis indicates the proportion of patients surviving with uncensored data. Overall survival was stratified by CD4:CD8 ratio.
Cytokine microarrays. We performed multiplex cytokine and chemokine microarrays using the LINCOplex assay on 22 matched preoperative plasma and ascites samples from 22 patients. Specimens were simultaneously evaluated for the presence of IL-1β, IL-1ra, IL-2, IL-4, IL-5, IL-6, IL-7, IL-8, IL-9, IL-10, IL-12, IL-13, IL-15, IL-17, bFGF, Eotaxin, G-CSF, GM-CSF, INF-γ, IP-10, MCP-1, MIP-1α, MIP-1β, PDGF-BB, RANTES, TNF-α, and VEGF. All assays were run in duplicate. Results in pg/ml for each cytokine or chemokine for each specimen were calculated as an average of the duplicate assays. The mean concentration and the standard error of the mean (SEM) was then calculated for each cytokine and chemokine. The mean concentration in ascites was compared to plasma using Student's t-test for each cytokine and chemokine. Ascites specimens demonstrated a significant elevated level of IL-6, IL-8, IL-10, IL-15, IP-10, MCP-1, MIP-1β and VEGF. Ascites specimens demonstrated a significantly reduced level of IL-2, IL-5, IL-7, IL-17, PDGF-BB, and RANTES. (p<0.05) (Table II and Figure 2). Only 12 specimens were evaluated by both FACS and Lincoplex. Not surprisingly given the small numbers, no correlation was noted between CD4/CD8 ratio and chemokine or cytokine levels (data not shown).
Lincoplex data.
T-cells from ovarian cancer associated ascites demonstrate limited anti-tumor response. Ovarian cancer cells derived from ascites were maintained in culture. In addition, lymphocytes were isolated from ovarian cancer ascites by Ficoll-Hypaque density gradient centrifugation. Lymphocytes from a single ovarian cancer patient (OC-01) were maintained in culture and stimulated with terminally radiated autologous tumor cells (OC-01). These stimulated lymphocytes served as effector cells. A chromium release assay was then performed. Tumor cells served as target cells. Autologous ovarian cancer cells (OC-01) and tumor cells from a different ovarian cancer patient (OC-02) were labeled with 51chromium. Stimulated lymphocytes (OC-01) were incubated at different ratios with labeled ovarian cancer target cells from either OC-01 or OC-02. Cytolytic activity was then measured based on 51chromium release from lysed target cells minus background. Stimulated lymphocytes from OC-01 demonstrated cytolytic activity against tumor cells from OC-01 but not against tumor cells from OC-02. It should be noted however that these T-cells could not be expanded and the observed anti-tumor response was limited even at high effector to target ratios (Figure 3).
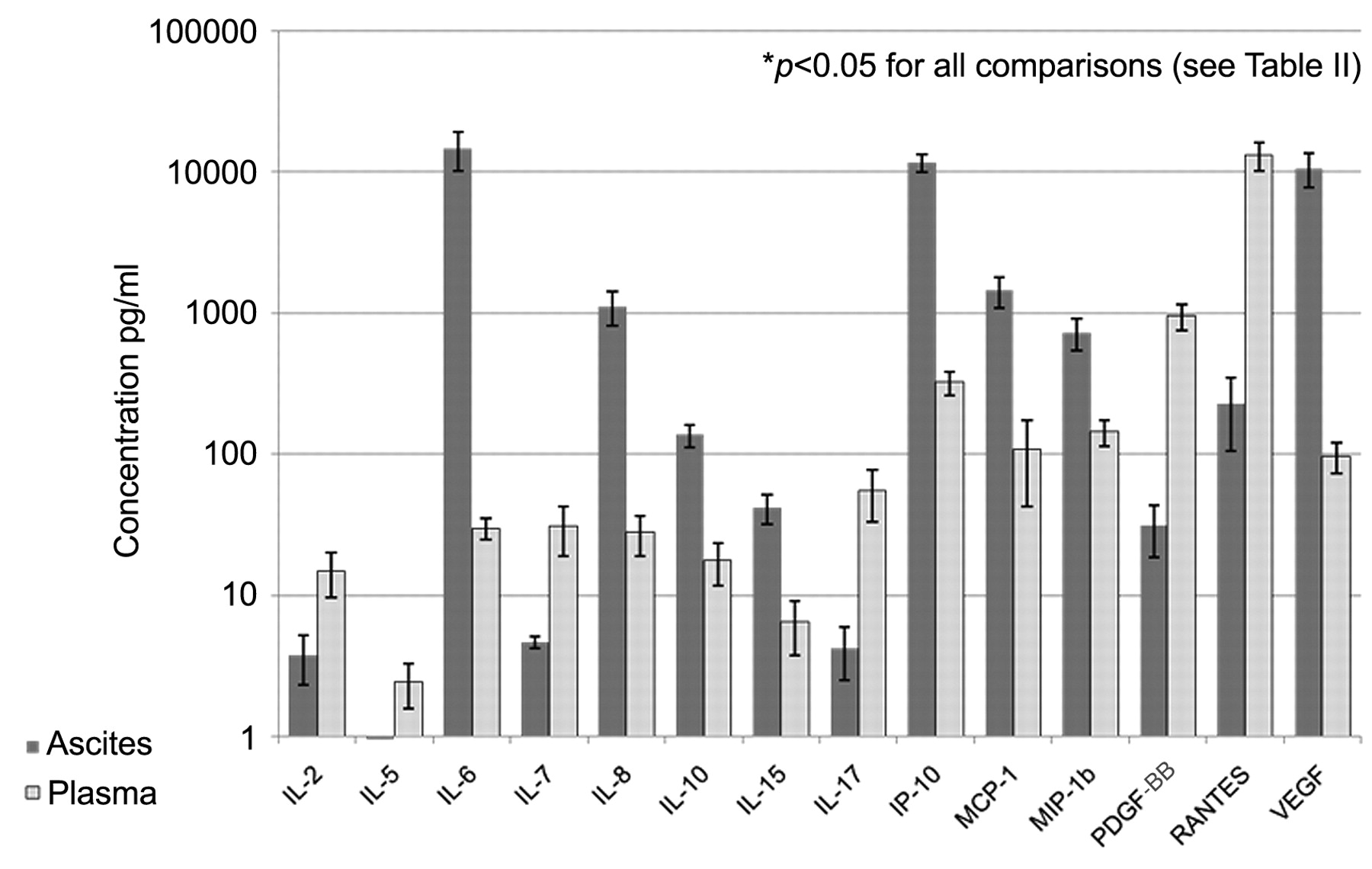
Multiplex cytokine microarrays using the Lincoplex assay on matched ovarian cancer patient ascites and plasma samples. Results are shown for 14 cytokines or chemokines with significant differences between ovarian cancer ascites levels and plasma levels. The x axis shows the 14 cytokines and chemokines (IL-6, IL-8, IL-10, IL-15, IP-10, MCP-1, MIP-1b, VEGF, IL-2, IL-5, IL-7, IL-17, PDGF-BB and RANTES); the y axis shows pg/ml. A log scale was used to allow for the data to be presented on one figure.
Artificial antigen presenting cell expansion of T-lymphocytes. Having found that tumor specific T-cells are present in the ascites, we tried to expand them in vitro using aAPC. We have previously reported that aAPC provide a reproducible method of stimulating and expanding antigen-specific CTL (23, 29). We chose 7 ovarian cancer-related peptides which have been previously described to be expressed in ovarian cancer cell lines (6-10, 17-20). Individual peptides were loaded onto aAPC and in separate assays, CD8+ T-lymphocytes isolated from ovarian cancer-associated ascites specimens were stimulated with the peptide-loaded aAPC. However in more than 25 experiments, we were not able to generate or expand a significant number of ovarian cancer-specific T-lymphocytes (data not shown).
Our findings could be secondary to the presence of immunosuppressive elements or to a low precursor frequency of ovarian cancer antigen-specific CTL. Therefore, we decided to attempt to expand CMV- and MART-1-specific CTL, which demonstrate a considerable precursor frequency in healthy controls. As with the above experiments, we enriched CD8+ T lymphocytes from plasma and ascites obtained from ovarian cancer patients.
For expanding CMV-specific CTL the precursor frequencies were 9% (0.9×105) in the peripheral blood mononuclear cell (PBMC) derived CD8 T-cells and 3% (0.1×105) in the ascites derived CD8+ T-cells as determined by CMV tetramer staining. After a single aAPC stimulation, the PBMC samples were 91% CMV specific, which represents a 35-fold expansion, while the ascites sample reached only a frequency of 11% CMV-specific cells that represents a only a 10-fold expansion of the CMV specific T-cells (Figure 4). In addition, even after 3 stimulations, the ascites sample only reached 60% specificity for CMV and never expanded to the level obtained by the PBMC sample after one stimulation (data not shown). Although CTL from ovarian cancer-associated ascites showed only limited expansion, these T lymphocytes appear functional as demonstrated by CD107α expression after antigen stimulation (data not shown).
Discussion
A growing body of evidence demonstrates the capacity of the immune system to recognize tumor-associated antigens and to develop antigen-specific humoral and cell mediated responses to these targets. Adoptive immunotherapy has resulted in tumor regression in melanoma patients (3). In ovarian cancer patients, a correlation between the presence of TIL and improved clinical has been reported (13) and suggests that efforts towards active and /or adoptive immunotherapy in this patient population may ultimately be successful. However despite the identification of many ovarian cancer-specific antigens, successful and reproducible immunotherapy for ovarian cancer has been elusive. The presence of a tumor-induced immunosuppressive environment, preventing individual elements of the immune system from providing a coordinated, effective antitumor response, could explain the disappointing results.

51Chromium release assay. Lymphocytes from an ovarian cancer patient demonstrated cytolytic activity against autologous tumor cells from OC-01 (dashed line) but not from OC-02 (solid line).
Ovarian cancer-specific CD8+ T lymphocytes can be induced and expanded in vitro and are capable of autologous tumor cell lysis. (30) In general, CD4+ T lymphocytes can either augment these cell-mediated immune responses as T helper cells, or inhibit these responses as regulatory T-cells. Santin and colleagues (31) report an excess of CD4+ compared to CD8+ T lymphocytes in multiple ovarian cancer-associated ascites specimens but not in TIL. Ioannides et al. (30) report similar findings with a higher percentage of CD4+ T-cells compared to CD8+ T-cells in non-stimulated TAL samples. Our findings suggest more diversity of ovarian cancer-associated ascites lymphocyte populations, with CD4+ T-cells predominating in some specimens and CD8+ T-cells predominating in others.
Recently, Sato and colleagues (32) evaluated 117 ovarian cancer specimens for the presence of T lymphocytes. Higher frequencies of intraepithelial CD8+ TIL were associated with improved outcome. This finding was not observed for stromal CD8+ TIL. Intraepithelial CD8/CD4 ratios ranged from 0-7.3. High intraepithelial CD8/CD4 ratios were associated with improved survival. Our data on ovarian cancer-associated ascites concurs with these data from ovarian cancer tumor specimens in that the ratio between cancer-associated CD4+ and CD8+ T lymphocytes appears to be predictive of ovarian cancer patient survival. A low CD4/CD8 (or high CD8/CD4) ratio were associated with significantly improved survival. Our data was limited by lower patient numbers (27 patients) and potentially by the inclusion of 2 patients with recurrent disease. However, the findings do confirm earlier data which suggest that the lymphocyte population found in ovarian cancer-associated ascites is representative of that found in the intraepithelial component of ovarian cancer tumor specimens.

Stimulation of ovarian cancer-associated ascites and plasma with CMV peptide-loaded aAPC. Cell numbers are given in 1×105. Values are given for total CTL number and CMV-specific CTL at day 0 and day 8 for both ovarian cancer-associated ascites and plasma. Plasma sample demonstrated 35-fold expansion (from 0.9×105 to 31.9×105) with high specify (91%). In contrast ovarian cancer-associated ascites demonstrated only 10-fold expansion (from 0.1×105 to 1.1×105) with low specificity (11%).
Curiel and associates (33) reported that CD4+ CD25+ FOXP3+ regulatory T-cells accumulate in tumors and ascites and that the presence of these cells was associated with reduced survival. The presence of CD4+ regulatory T-cells in ovarian cancer ascites could explain our findings of an association between a high CD4/CD8 ratio and poor outcome. The wide range of CD4/CD8 ratios suggests that regulatory T-cells represent one mechanism by which some but not all ovarian malignancies may elude immune surveillance.
Secreted immunosuppressive factors could both inhibit innate host anti-tumor immune responses and prevent successful induction of such a response by immunotherapy. Cell-mediated immunity is required for effective anti-tumor responses and is regulated by Th1 (enhancing) and Th2 (inhibitory) cytokines. Th1 cytokines include IL-2, IL-3, INF-γ, and TNF-α. Th-2 cytokines include IL-4, IL-5, IL-6, IL-10, and IL-13. In addition to functioning as a Th-2 cytokine, IL-6 has been associated with ascites formation in ovarian cancer patients by up-regulating VEGF expression which leads to increased vascular permeability (34). Several investigators have evaluated cytokine concentrations in ovarian cancer associated ascites and found increased levels of inhibitor cytokines such as IL6 and Il-10 but in some cases also elevated levels of stimulatory cytokines such as TNF-α and IL-1b compared to normal controls (35, 36). In addition, Xie and associates (37) noted decreased levels of IL-7, which appears to have similar enhancing effects as IL-2, in ovarian cancer-associated ascites compared to serum.
We compared levels of 27 cytokines and chemokines in ovarian cancer-associated ascites and serum. This is the most comprehensive comparison of plasma and ovarian cancer-associated ascites to date. We found significant differences in 14 of the 27 tested cytokines and chemokines. These alterations included significantly elevated levels of IL-6, IL-8, IL-10, IL-15, IP-10, MCP-1, MIP-1β and VEGF and significantly reduced levels of IL-2, IL-5, IL-7, IL-17, PDGF-BB and RANTES in ovarian cancer-associated ascites compared to serum. These differences could influence immune-mediated anti-tumor responses.
As mentioned, several interleukins are clearly associated with a Th1 or Th2 response. A significantly lower level of IL-2 in ascites is consistent with a reduced Th1 response. Although lower levels of IL-2 mRNA in ovarian cancer ascites has been reported (38), this is the first investigation to note a significant reduction in the actual cytokine. IL-7, which is important for lymphocyte survival, was also significantly reduced in ovarian cancer-associated ascites. The Th2 cytokines IL-6 and IL-10 were significantly elevated in ovarian cancer ascites compared to serum. The Th2 cytokine IL-5 was reduced in ascites (0.5 pg/ml), but also very low in plasma (2.4 pg/ml), which was the lowest level for any serum cytokine or chemokine. These findings together indicate a reduction in Th1 (enhancing) response and an increase in Th2 (inhibitory) response both of which could lead to an inhibition of cell mediated anti-tumor response.
We identified alterations in additional interleukins that could potentially enhance tumor progression. Overexpression of the proangiogenic chemokine IL-8 has been associated with poor outcome in ovarian cancer (39). IL-17 is a proinflammatory cytokine which appears to induce anti-tumor T-cell response (40). In ovarian cancer-associated ascites, elevated IL-8 levels may lead to neovascularization of tumor implants and decreased IL-17 levels may reduce anti-tumor immune response.
Several chemokines could affect tumor progression. Our data confirms earlier findings of elevated levels of the chemokine, VEGF, in malignant ovarian cancer ascites compared to serum. (41) Overexpression of VEGF appears to play a role in ovarian cancer progression and blocking this pathway appears to improve survival (42, 43). Neutralization of the chemokine MCP-1 with antibody results in reduced tumor burden in a mouse prostate cancer model (44). RANTES is a chemokine, which appears to induce the activation and of natural-killer cells and enhances anti-tumor immunity in a mouse model (45). The over-expression of VEGF and MCP-1 and the reduction of RANTES in ovarian cancer-associated ascites may provide pathways for tumor growth.
Other changes found in cytokine and chemokine levels in ascites may limit tumor growth. IL-15 binds to the IL-2 receptor and induces both antigen-independent expansion and long-term survival of anti-tumor CD8+ CTL (46). In addition, it has been reported that IL-15 induces and enhances anti-tumor CTL responses against an ovarian cancer cell line (47). The chemokines IP-10 and MIP-1β are both elevated in ovarian cancer-associated ascites. Expression of IP-10 enhances anti-tumor responses in a mouse central nervous system model (48). Intratumoral expression of MIP-1β induced inhibition of colorectal tumor growth and associated tumor-specific CTL response (49). Therefore in contrast to our other findings, the overexpression of IL-15, IP-10 and MIP-1β in ovarian cancer-associated ascites could be associated with inhibition of tumor growth.
PDGF regulates cell growth and division with a significant role in angiogenesis. PDGF induces VEGF production in ovarian cancer cells and expression of PDGF-BB has been correlated with VEGF expression in ovarian cancer ascites. (41) Our data is the first to demonstrate a significantly decreased expression of PDGF-BB in ovarian cancer associated ascites compared to plasma. A reduction in PDGF-BB expression should be associated with a reduction in VEGF. However, we observed elevated VEGF levels compared to plasma. This suggests the effects of reduced PDGF-BB expression are outweighed by stimulatory effects of increased IL-6 levels on VEGF expression.
While our observations indicate a mixed picture, the overall comparison of cytokine and chemokine levels in ovarian cancer associated ascites and plasma suggests an interleukin pattern favoring a Th2 (inhibitory) pattern in ascites. Nevertheless, reduced to a single patient, these findings suggest a patient specific battle between the immune system and the ovarian malignancy with actions by the immune system to limit tumor growth and responses by the ovarian cancer to circumvent these measures.
We utilized chromium release assays and aAPC to determine the potential for antigen-specific CD8+ T-cell response in PBMC from ovarian cancer patients and in ovarian cancer-associated ascites. CD8+ T lymphocytes, isolated from ovarian cancer-associated ascites demonstrate only limited anti-tumor activity in vitro, even at high effector to target ratios. Attempts to expand ovarian cancer antigen-specific T lymphocytes from ovarian cancer using aAPC have been unsuccessful. Given our previous success with expansion of anti-CMV T lymphocytes isolated from the plasma of healthy donors (23) we attempted to expand CMV-specific T-cells from ovarian cancer patients. Using aAPC, we successfully expanded anti-CMV Tlymphocytes from ovarian cancer patients, but detected only limited expansion from CTL isolated from ovarian cancer-associated ascites. Despite the modest expansion, these T-cells appear functional with appropriate target lysis.
Interestingly, while these findings could potentially explain the limited cytotoxic activity and expansion of the ascites derived CTL described in Figure 3, they should not impact on the aAPC stimulation experiment. In this experiment, we have not only removed the T-cells from their inhibitor cytokine milieu, but also depleted potentially regulatory CD4+ T-cells. The fact that the CD8+ T-cells still did not respond as expected suggests that the T-cells were either previously altered by their environment or it could reflect the presence of a subpopulation of regulatory CD8+ T lymphocytes that have been previously described by Zou et al. (50) These regulatory CD8+ T-cells lack cytotoxic effector functions and secrete inhibitory cytokines upon stimulation.
The immune system appears capable of effective anti-tumor responses against melanoma and several other cancers. However, such responses have not been associated with ovarian malignancies. Our findings demonstrate two potential mechanisms of ovarian cancer-mediated suppression of effective anti-tumor response including the presence of regulatory T-cells and secreted inhibitory factors. The association of elevated CD4/CD8 ratio with poor outcome may reflect suppression of immune function by regulatory T-cells, which could explain increased levels of inhibitory cytokines and chemokines in the ascites fluid and the non-responsiveness of ascites derived T-cells against autologous tumor cells as well as against antigen-specific aAPC stimulation. In contrast, aAPC stimulation of PBMC-derived T-cells in matched samples resulted in induction and expansion of antigen-specific T-cells as expected. Future studies to dissect detailed mechanisms of the observed immunosupression are ongoing. The identification and neutralization of these inhibitory cells and factors will improve the likelihood of developing successful ovarian cancer-specific immunotherapy.
Acknowledgements
This study was supported by the Gynecologic Cancer Foundation/Ann Schreiber Ovarian Cancer Research Grant.
Footnotes
-
Presented (Posters) at the 39th and 40th Annual meeting of the Society of Gynecologic Oncologists (2008, 2009).
- Received April 21, 2009.
- Accepted June 11, 2009.
- Copyright© 2009 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Society for Immunotherapy of Cancer (SITC) clinical practice guideline on immunotherapy for the treatment of gynecologic cancer
- Humanized NBSGW PDX Models of Disseminated Ovarian Cancer Recapitulate Key Aspects of the Tumor Immune Environment within the Peritoneal Cavity
- Genetically Defined Syngeneic Mouse Models of Ovarian Cancer as Tools for the Discovery of Combination Immunotherapy
- Rapid tumor vaccine using Toll-like receptor-activated ovarian cancer ascites monocytes
- Circulating CD14high CD16low intermediate blood monocytes as a biomarker of ascites immune status and ovarian cancer progression
- A mosaic renal myeloid subtype with T-cell inhibitory and protumoral features is linked to immune escape and survival in clear cell renal cell cancer
- Increased Immunosuppression Is Related to Increased Amounts of Ascites and Inferior Prognosis in Ovarian Cancer
- Overexpression of thymic stromal lymphopoietin is correlated with poor prognosis in epithelial ovarian carcinoma
- Alternatively-Activated Macrophages Upregulate Mesothelial Expression of P-Selectin to Enhance Adhesion of Ovarian Cancer Cells
- VEGF Potentiates GD3-Mediated Immunosuppression by Human Ovarian Cancer Cells
- A Low-density Antigen Array for Detection of Disease-associated Autoantibodies in Human Plasma
- CD47 Enhances In Vivo Functionality of Artificial Antigen-Presenting Cells
- Revisiting the Complexity of the Ovarian Cancer Microenvironment--Clinical Implications for Treatment Strategies
- Haptoglobin Phenotype and Epithelial Ovarian Cancer
- Role of Soluble {beta}-(1-3),(1-6)-D-Glucan from Saccharomyces cerevisiae in the Murine P388 Ascites Tumor Model