


Abstract
The side population (SP) and the CD44+/CD24-/low population have been reported in separate studies to include more tumorigenic cells than other populations, and to have the ability to form new tumors and undergo heterogeneous differentiation in breast cancer tissue. However, the relationship between these two populations has not yet been explored in breast cancer cells. Here it is shown that the SP and the CD44+/CD24-/low populations are overlapping. Both populations were resistant to paclitaxel. Components of the Hedgehog (Hh) signaling pathway were more highly expressed in these cell populations at both the mRNA and protein levels compared with other populations. Furthermore, inhibition of Hh signaling activity suppressed the proliferation of both populations. The significance of Hh signaling activity in the proliferation of both populations was confirmed by the effect of an si-RNA against Gli1, a trans-activator of the Hh signaling pathway, on the proliferation of both populations. These data suggest that the Hh signaling pathway is essential for the proliferation of the tumorigenic population of breast cancer cells, and that this pathway might represent a new candidate for breast cancer therapy targeting cancer stem cells.
It has been hypothesized that the capability of a tumor to grow and propagate is dependent on a small subset of cells within a tumor, termed cancer stem cells (CSCs). CSCs are thought to proliferate without limit and provide heterogeneous cells including new CSCs (1-3). Data have been provided to support this theory in human lymphoma, glioblastoma, and some other types of cancer (4-9). Furthermore, the supposed significance of CSCs in cancer therapy is now greater because evidence has suggested that CSCs provide resistance to chemotherapy and radiation therapy (10, 11) and that they cause the recurrence of cancer through metastasis (12, 13).
In breast cancer, CD44+CD24-/low subpopulation cancer cells have been reported to have stem/progenitor cell properties (14, 15). Al-Haji et al. reported that CD44+CD24-/low subpopulation cells in human breast cancer patients exclusively retain the ability to form new tumors in a NOD-SCID mouse xenograft model. The phenotypes of these tumors were quite similar to those of the original tumors, although other populations had no such ability (14). Sheridan et al. reported the existence of a CD44+CD24-/low population in cell lines and that the cells of this population possessed more invasive and proliferating properties than those of other populations (16).
From a clinical point of view, Abraham et al. investigated the correlation between the prevalence of CD44+CD24-/low cells in breast cancer tissues from patients and their clinical outcomes. Although there was no significant correlation between the prevalence of CD44+CD24-/low cells and tumor progression, tumors with a high percentage of CD44+CD24-/low tumor cells developed significantly more distant metastases than did other tumors (17). At the same time, Liu et al. compared the gene-expression profile of CD44+CD24-/low cancer cells with that of normal breast epithelium.
Goodell observed that when bone marrow-derived cells are incubated with Hoechst dye 33342 and analyzed by flow cytometry, a small population of cells does not accumulate an appreciable amount of dye and these were identified to be a Hoechst side population (SP) (18). Like CD44+CD24-/low population cells, the SP is also enriched with tumorigenic stem-like cancer cells expressing high levels of ABCG2, an ATP-binding cassette half-transporter associated with multidrug resistance in breast cancer cell lines (15).
The Hedgehog (Hh) signaling pathway is crucial for growth and patterning during embryonic development (19). This pathway is a highly coordinated and orchestrated network, involving inhibition of the twelve transmembrane protein Patched1 (Ptch1) by binding Hh protein, activation of the seven transmembrane protein Smoothened (Smo), release of a five-zinc finger transcription factor Gli from a large protein complex, nuclear translocation of Gli, and transcription of target genes (19-21).
The Hh pathway has been established as an important signaling system in mouse normal mammary gland development (22, 23). Previously, it was reported that the Hh signaling pathway is highly activated in human breast cancer cells and in resected breast cancer tissues compared with adjacent normal tissue. Disruption of the activity of the Hh signaling pathway suppressed the proliferation of cancer cells (24). Thus, it was hypothesized that the Hh pathway might represent a new candidate therapeutic target for the treatment of breast cancer. Furthermore, it was reported that the Hh signaling pathway is important for signal transduction downstream of the estrogen receptor (ER), one of the main regulators of normal breast stem cells (25, 26). ER stimuli activated the Hh signaling pathway through Sonic Hedgehog (Shh); conversely, inhibition of Hh activation suppressed the effect of ER stimulation on cancer cell proliferation.
Liu et al. reported that Hh genes are highly expressed in normal human mammary stem cells/progenitor cells cultured as mammospheres, and in CD44+CD24-/low cancer cells (27). However, the exact roles of the Hh signaling pathway in supposed breast CSCs, CD44+CD24-/low cancer cells and SP cells have not yet been revealed.
Based on the results of these studies, it is hypothesized that the Hh signaling pathway might maintain the breast CSC population and that this pathway could represent a treatment target for breast cancer. This paper reports that two supposedly separate breast CSC populations are overlapping, and that the Hh signaling pathway is essential for maintaining breast CSCs. In this report, it is revealed for the first time that the Hh pathway might represent a valuable target for breast cancer stem cell therapy.
Materials and Methods
Cell culture, reagents and antibodies. The human breast adenocarcinoma cell line MCF-7, the human colonic adenocarcinoma cell line DLD-1 and the human pancreatic ductal adenocarcinoma cell line AsPC-1 were maintained as monolayer cultures in complete medium composed of RPMI-1640 and 10% fetal bovine serum (FBS) (both Sigma Chemical, St. Louis, MO, USA). Paclitaxel, purchased from Sigma-Aldrich, was diluted in PBS and, cyclopamine, purchased from Toronto Research Chemicals (North York, ON, Canada), was diluted in ethanol.
Proliferation assay. Cells (4×106/well) seeded in 75 cm2 flasks in complete culture medium were incubated overnight. The medium was changed to 5% FBS-RPMI containing 0, 10, 20 nM of paclitaxel and 0, 10, 30 μM of cyclopamine. After 72 hours' incubation, cells were harvested by trypsinization and viable cells counted with a Coulter counter (Beckman Coulter, Fullerton, CA, USA).
Cell staining for flow cytometry. Cells were counted and then transferred to a 10-ml tube, washed twice with phosphate-buffered saline (PBS) containing 2% heat-inactivated fetal calf serum (HICS), centrifuged for 7 min at 1,400 rpm, and resuspended in 100 μL (per 106 cells) of PBS containing 2% HICS. Antibodies (appropriate dilution per antibody) were then added (see below) and suspensions were incubated for 20 min on ice, and then washed twice with PBS/2% HICS. When needed, a secondary antibody was added by resuspending cells in 100 μl (per 106 cells) of PBS/2% HICS, and then adding 1-4 μl of secondary antibody (depending on the secondary antibody and its concentration), followed by a 20-min incubation. Cells were washed twice with PBS/2% HICS and resuspended in 0.5 ml (per 106 cells) of PBS/2% HICS containing propidium iodide (2 μg/ml final concentration).
Flow cytometry. The protocol was based on that of Al-Hajj et al. (14), with slight modifications. Briefly, cells (1×106 cells/ml) were counted, washed twice with PBS/2% HICS and resuspended in 100 μl (per 106 cells) of PBS/2% HICS. Five microliters of Sandogloblin solution (1 mg/ml) was then added and cells were incubated on ice for 10 min, after which they were washed twice with PBS/2% HICS and resuspended in 100 μl (per 106 cells) of PBS/2% HICS. Antibodies (at appropriate dilutions) were then added and cells were incubated for 20 min on ice. At the end of the incubation, cells were spun down at 4°C and resuspended in ice-cold PBS/2% HICS.
The cells were stained with anti-CD44 (FITC) and anti-CD24 (PE). Anti-mouse IgG (FITC) and anti-mouse IgG (PE) were also used as isotype controls to set up gates. Unless otherwise noted, antibodies were purchased from PharMingen (Franklin Lakes, NJ, USA). Antibodies were directly conjugated to different fluorochromes, depending on the experiment. Flow cytometry was performed on an EPICS Ultra system (Beckman Coulter). Side scatter and forward scatter profiles were used to eliminate cell doublets. Propidium iodide (PI) staining was used to eliminate dead cells. Cells were routinely sorted twice and reanalyzed for purity, which was typically >95%.
Side population analysis. The protocol was based on that of Goodell et al. (18), with slight modifications. Briefly, cells (1×106 cells/ml) were incubated in prewarmed PBS/2% HICS containing freshly added Hoechst 33342 (5 μg/ml final concentration) for 60 minutes at 37°C with intermittent mixing. In some experiments, cells were incubated with Hoechst dye in the presence of verapamil (50 μmol/l) or reserpine (100 μmol/l). At the end of the incubation, cells were spun down at 4°C and resuspended in ice-cold PBS. Propidium iodide (2 μg/mL final concentration) was added 5 minutes before fluorescence-activated cell sorting (FACS) analysis, allowing the discrimination of dead versus live cells. Samples were analyzed on an EPICS Ultra system (Beckman Coulter). The Hoechst dye was excited with a UV laser at 351 to 364 nm, and its fluorescence was measured with a 515-nm side population filter (Hoechst blue) and a 608 EFLP optical filter (Hoechst red). A 540 DSP filter was used to separate the emission wavelengths.
Real-time reverse transcription-PCR. To assess the expression levels of Shh, Gli1 and Ptch1, total RNA from sorted cells was extracted using a Qiagen (Valencia, CA, USA) RNeasy Mini Kit (catalog no. 74104) according to the manufacturer's instructions. RNA (1 μg) was quantified by spectrophotometry (Ultraspec 2100 Pro, Amersham Pharmacia Biotech, Cambridge, UK), treated with DNase and reverse transcribed to cDNA using a Quantitect Reverse Transcription kit (Qiagen) according to the manufacturer's instructions. Reactions were performed using iQ SYBR Green Supermix (Bio-Rad, Hercules, CA, USA) on a DNA Engine Opticon 2 System (MJ Research, Waltham, MA, USA). cDNAs prepared from AsPC1 and SUIT-2 cells transfected with pSHH-GFP were serially diluted in 10-fold increments and amplified in parallel with various primer pairs to generate standard curves. Each reaction was performed in triplicate. All primer sets amplified fragments <200 bp long. The sequences of the primers used were as follows: β-actin forward, 5′-TTGCCGACAGGATGCAGAAGGA-3′ and reverse, 5′-AGGTGGACAGCGAGGCCAGGAT-3′; Shh forward, 5′-GTGTACTACGAGTCCAAGGCAC-3′ and reverse, 5′-AGGAA GTCGCTGTAGAGCAGC-3′; Gli1 forward, 5′-GGTTCAAGAG CCTGGGCTGTGT-3′ and reverse, 5′-GGCAGCATTCTCAG TGATGCT-3′ and Ptch1 forward 5′-ATGCTGGCGGATCTGA GTTCGACT-3′, and reverse, 5′-GGGTGTGGGCAGGCGGTTC AAG-3′. The amount of each target gene in a given sample was normalized to the level of β-actin in that sample.
Immunostaining of cell lines. A human breast adenocarcinoma cell line (MCF-7), a human pancreatic adenocarcinoma cell line (AsPC1) and a human colonic adenocarcinoma cell line (DLD-1) were maintained as monolayer cultures in complete medium composed of RPMI 1640 and 10% FBS (Sigma Chemical). Cells (2-4×105 cells/well) were incubated on 8-well CultureSlides (BIOCOAT; Becton Dickinson) for 4 hours at 37°C. The slides were air-dried and immersed in 100% methanol for 5 minutes at -30°C. All primary antibodies were incubated with cells overnight at 4°C. The primary antibodies used were: Shh (N-19, sc-1194) and Gli1 (N-16, sc-6153) (1:250; Santa Cruz Biotechnology, Santa Cruz, CA, USA). Secondary antibodies (rabbit anti-goat inmunogloblin; Nichirei Co., Ltd., Tokyo, Japan) were applied for 1 hour at room temperature. DLD-1 and AsPC-1 cells were used as negative and positive controls for the Hh signaling pathways, respectively (24).
Small interfering RNA (siRNA) against Gli1. MCF-7 cells (2.0×106 cells) were transfected with an siRNA (100 nM) against Gli1 by nucleofection using a NucleoFector II kit (Amaxa GmbH, Koeln, Germany) according to the manufacturer's instructions, and plated in a 25-cm2 flask for 24 hours in 10% FBS-RPMI. A validated stealth RNAi against Gli1 and a stealth RNAi negative control (Invitrogen Carlsbad, CA, USA) were used.
Xenograft tumor experiments and in vivo tumorigenicity. For tumor experiments, different numbers of sorted cells (CD44+/CD24-/low cells and CD44+CD24+ cells) were injected in 40 μl of medium/Matrigel (1:1) subcutaneously, into female non-obese diabetic/severe combined immunodeficiency (NOD/SCID) mice (4-8 weeks old). All animals were obtained from The Charles River Laboratory (Wilmington, MA, USA) and maintained in standard conditions according to the institutional guidelines. Tumor development was monitored starting from the second week after injection. The primary tumor sizes were measured with a caliper on a weekly basis and approximate tumor weights were determined using the formula 0.5ab2, where b is the smaller of the two perpendicular diameters. Tumorigenicity was measured mainly by tumor incidence (that is, the number of tumors/number of injections) and latency (that is, time from injection to detection of palpable tumors). All animals were sacrifized 6 to 9 months after tumor cell injection.
Statistical analysis. Student's t-test was used for statistical analysis. Relationships between variables were assessed using the Spearman rank correlation coefficient. A p<0.05 was considered significant.
Results
CD44+/CD24-/low cells and SP cells are enriched in both populations of breast cancer cells. The CD44+/CD24-/low population and the side population have been reported to be rich among tumorigenic breast cancer cells, even in established breast cancer cell lines (16, 28). CD44+/CD24-/low cells were first isolated from MCF-7 cells by flow cytometry. Approximately 35% of the MCF-7 cells were CD44+/CD24-/low cells (Figure 1B). Sorted CD44+/CD24-/low cells were labeled with Hoechst, after which they were analyzed again by UV laser. Approximately 38.28% of the CD44+/CD24-/low cells belonged to the SP, whereas only 1.83% of the MCF-7 cells belonged to this population (Figure 1C, D). SP cells were then isolated from MCF-7 cells and the number of CD44+/CD24-/low cells was examined. Sorted SP cells were incubated with anti-CD44 (FITC) and anti-CD24 (PE) on ice and analyzed again by argon laser. The SP contained a high proportion of CD44+/CD24-/low cells, 68%, whereas only 35% of MCF-7 cells possessed these characteristics (Figure 1F, area B). These data indicated that there was some overlap between the CD44+/CD24-/low cell population and the SP.
CD44+/CD24-/low cells and SP cells maintain differentiation potential. CD44+/CD24-/low cells were purified from MCF-7 cells by flow cytometry with a sorting fidelity of 95%, and incubated for 72 hours in RPMI+10% FBS. After incubation, 35.1% of the purified CD44+/CD24-/low cells differentiated into CD44+/CD24+ cells and 50.0% differentiated into CD44+/CD24-/low cells (Figure 2A); by contrast, CD44+/CD24+ cells retained the same character, without differentiating into CD44+/CD24-/low cells (Figure 2B). The differentiation ability of SP cells was also examined. After purifying SP cells with a sorting fidelity of 93%, they were incubated for 72 hours in RPMI+10% FBS and subjected to flow cytometry. Among SP cells, 86.91% differentiated into non-SP cells and 13.09% differentiated into SP cells (Figure 2C). The sorted non-SP cells were hard to grow because of the toxicity of Hoechst 33342. These data indicated that both CD44+/CD24-/low cells and SP cells have the ability to differentiate.
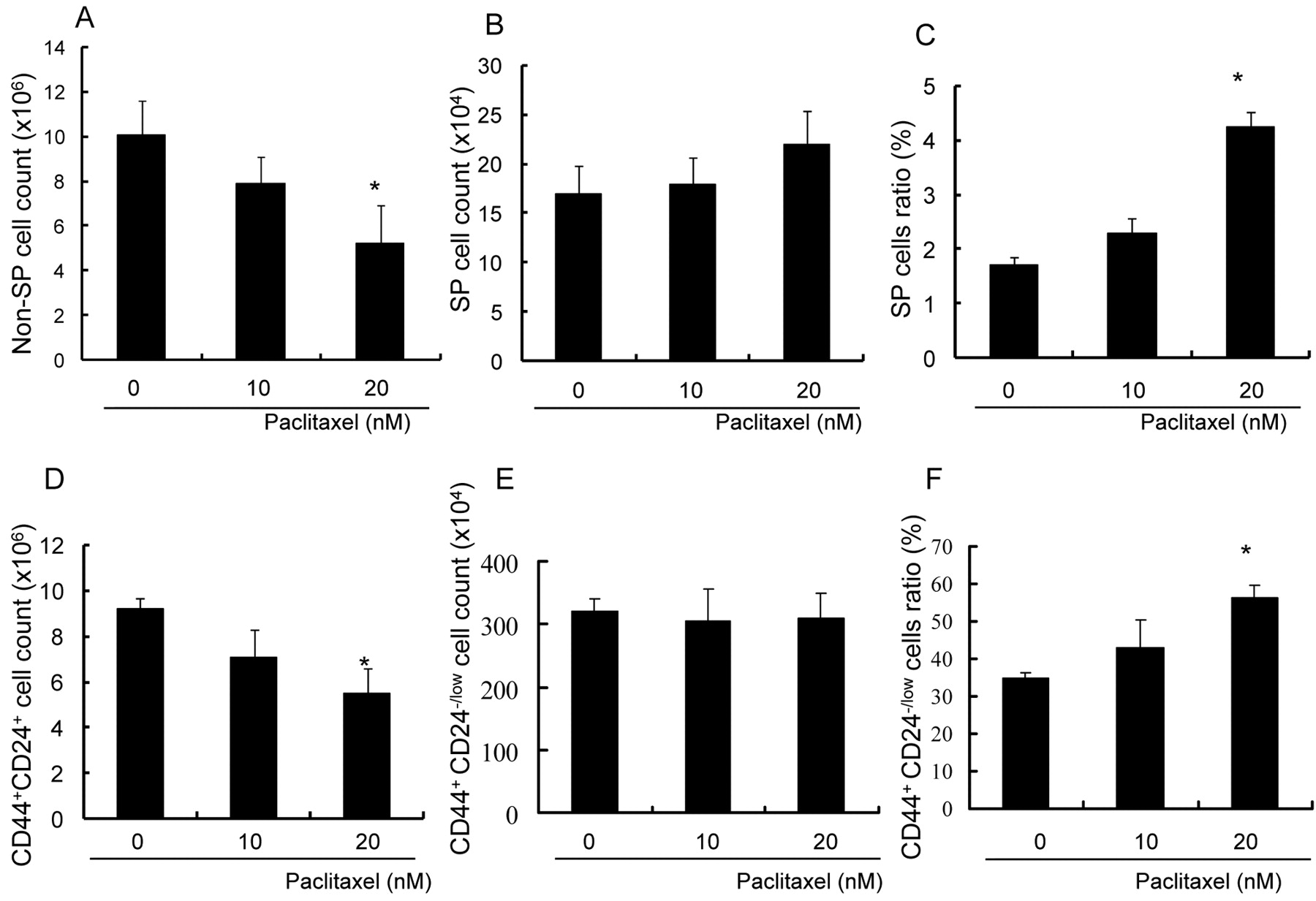
CD44+/CD24-/low cells and SP cells are resistant to paclitaxel. SP cells were originally isolated as multi-drug resistance gene-expressing cells (10). It was confirmed that the SP cells purified from MCF-7 cells were resistant to paclitaxel. MCF-7 cells were incubated with 10 and 20 nM of paclitaxel for 72 hours and counted by flow cytometry. Although the numbers of non-SP cells decreased significantly, the numbers of SP cells did not decrease, even in the presence of Paclitaxel (Figure 3A, B). Eventually, the ratio of SP cells to all cells became markedly increased (Figure 3C). CD44+/CD24-/low cells were also examined for resistance to paclitaxel. The number of cells in the CD44+/CD24-/low subpopulation did not decrease in the presence of paclitaxel, in contrast to the number of CD44+/CD24+ cells (Figure 3D, E), leading to an increase in the ratio of CD44+/CD24-/low cells to all cells (Figure 3F). These data indicated that not only SP cells, but also CD44+/CD24-/low cells showed paclitaxel resistance.

Identification of a CD44+/CD24-/low subpopulation and the side population (SP) in the breast cancer cell line MCF-7 by flow cytometry. A, Flow cytometric density plot demonstrating isotype control. B, Flow cytometric density plot demonstrating determination of the CD44+/CD24-/low subpopulation (Gate A 35%) and the CD44+/CD24+ subpopulation (Gate B 53%) by double staining. C, Isolated CD44+/CD24-/low cells were immediately dyed with Hoechst33342 and re-analyzed by flow cytometry after the first sort. They comprised 38% SP cells (Gate SP), whereas only 1.83% of MCF-7 cells were SP cells. D, MCF-7 cells were dyed with Hoechst 33342 and analyzed by UV laser flow cytometry to determine the SP (Gate SP 1.8%) and main population (Gate MP 55%). E, SP was completely inhibited by verapamil. F, Isolated SP cells were stained with anti-CD24 and anti-CD44 antibodies immediately and re-analyzed after the first sort. They contained 46% SP cells (Gate A), whereas only 16% of MCF-7 cells were SP cells.
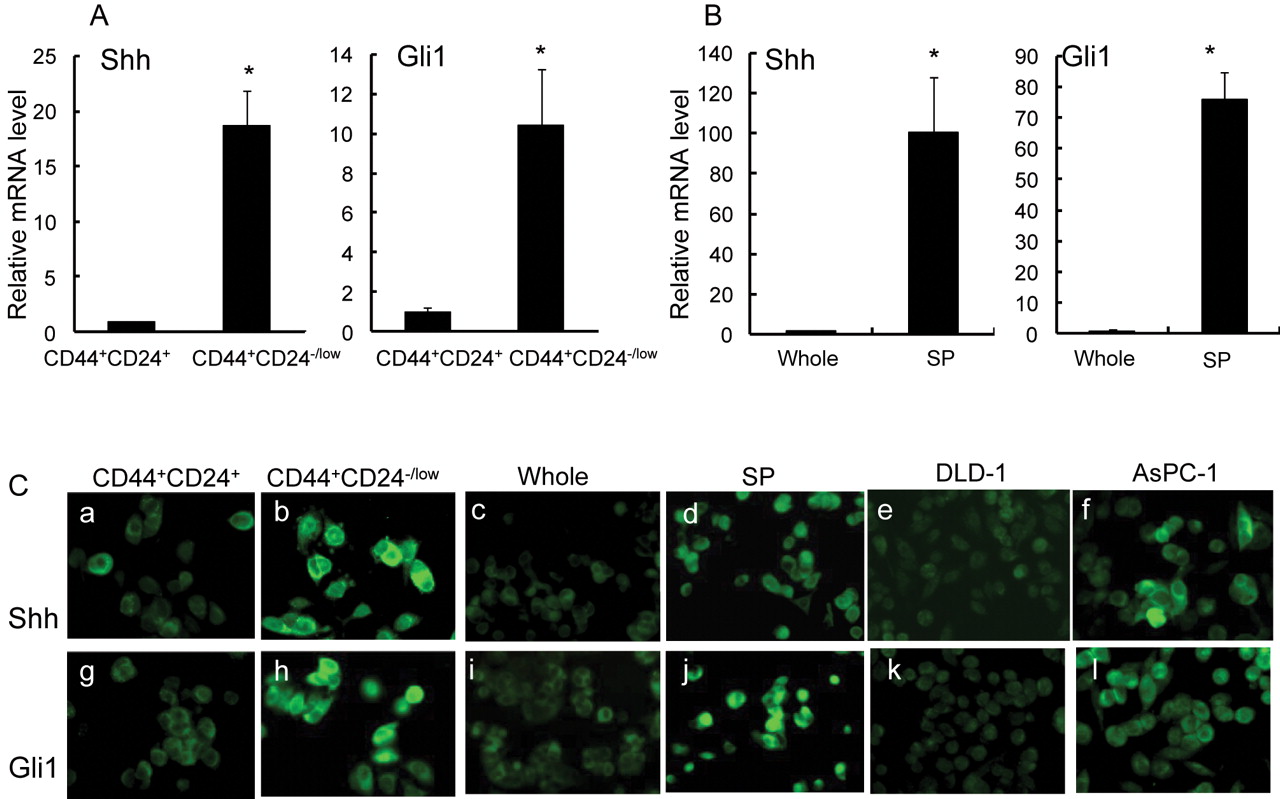
Components of the Hedgehog signaling pathway are highly expressed in CD44+/CD24-/low cells and SP cells. The expression levels of components of the Hh signaling pathway in CD44+/CD24-/low cells and SP cells were examined. CD44+/CD24-/low cells and SP cells were purified from MCF-7 cells and subjected to real-time RT-PCR. Shh, one of three Hh ligands, and Gli1, a trans-activator of Hh, were highly expressed in CD44+/CD24-/low cells, in contrast to CD44+CD24+ cells (Figure 4A). SP cells also showed high-level expression of Shh and Gli1, in sharp contrast to unsorted cells (Figure 4B). Fluorescence immunostaining revealed high signal intensities for both Shh and Gli1 in sorted CD44+/CD24-/low cells, but low levels in CD44+CD24+ cells. These two factors were also highly expressed in sorted SP cells compared with unsorted cells (Figure 4C), consistent with the results of real-time PCR.

CD44+/CD24-/low cells and SP cells maintain their differentiation potential. Isolated CD44+/CD24-/low cells and SP cells incubated for 72 hours in RPMI+10% FBS, re-analyzed by flow cytometry. A, Sorted CD44+/CD24-/low cells differentiated into 50.44% CD44+/CD24-/low cells (Gate A) and 35.16% CD44+/CD24+ cells (Gate B) after a 72-hour incubation. B, Sorted CD44+/CD24+ cells included almost no CD44+/CD24-/low cells (Gate A 0.4%). C, Sorted SP cells differentiated into 13.09% SP cells and 86.91% non-SP cells after a 72-hour incubation.
The suppression of Hh activity in CD44+/CD24-/low cells and SP cells leads to growth suppression of cancer cells. To explore the role of activated Hh activity in the proliferation of CD44+/CD24-/low cells and SP cells, sorted cells were incubated with cyclopamine, an inhibitor of the Hh signaling pathway. Total cell number was reduced by incubating these cells with cyclopamine (Figure 5A). The number of CD44+/CD24-/low cells was reduced dramatically by cyclopamine; eventually the ratio of CD44+/CD24-/low cells to all cells also decreased (Figure 5B, C). In a similar way, the number of SP cells was reduced significantly by cyclopamine, and the ratio of SP cells to all cells was also decreased (Figure 5D, E). These data suggested that the proliferation of CD44+/CD24-/low cells and SP cells may depend more on Hh activity than the whole population of breast cancer cells.
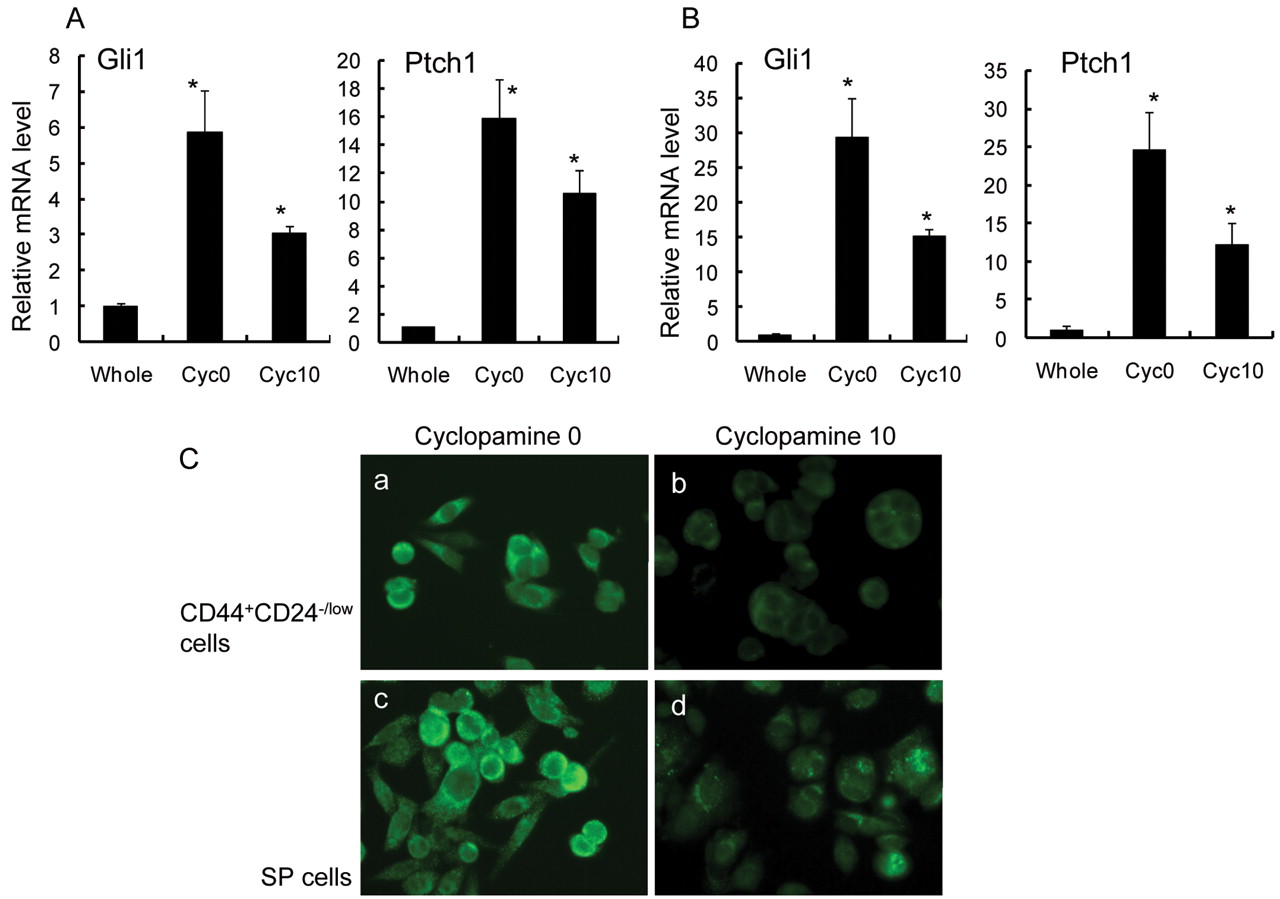
To confirm a decrease in Hh signaling activity in the presence of cyclopamine, the expression levels of Hh target genes were analyzed by real-time RT-PCR. The expression levels of Gli1 and Ptch1 were clearly suppressed after incubation of both CD44+/CD24-/low cells and SP cells with cyclopamine (Figure 6A, B). These findings were further confirmed by immunfluorescence studies. Both CD44+/CD24-/low cells and SP cells were stained for Gli1, a trans-activator and one of the targets of the Hh signaling pathway, after incubation with cyclopamine. The intensity of staining for Gli1 was significantly reduced by cyclopamine.
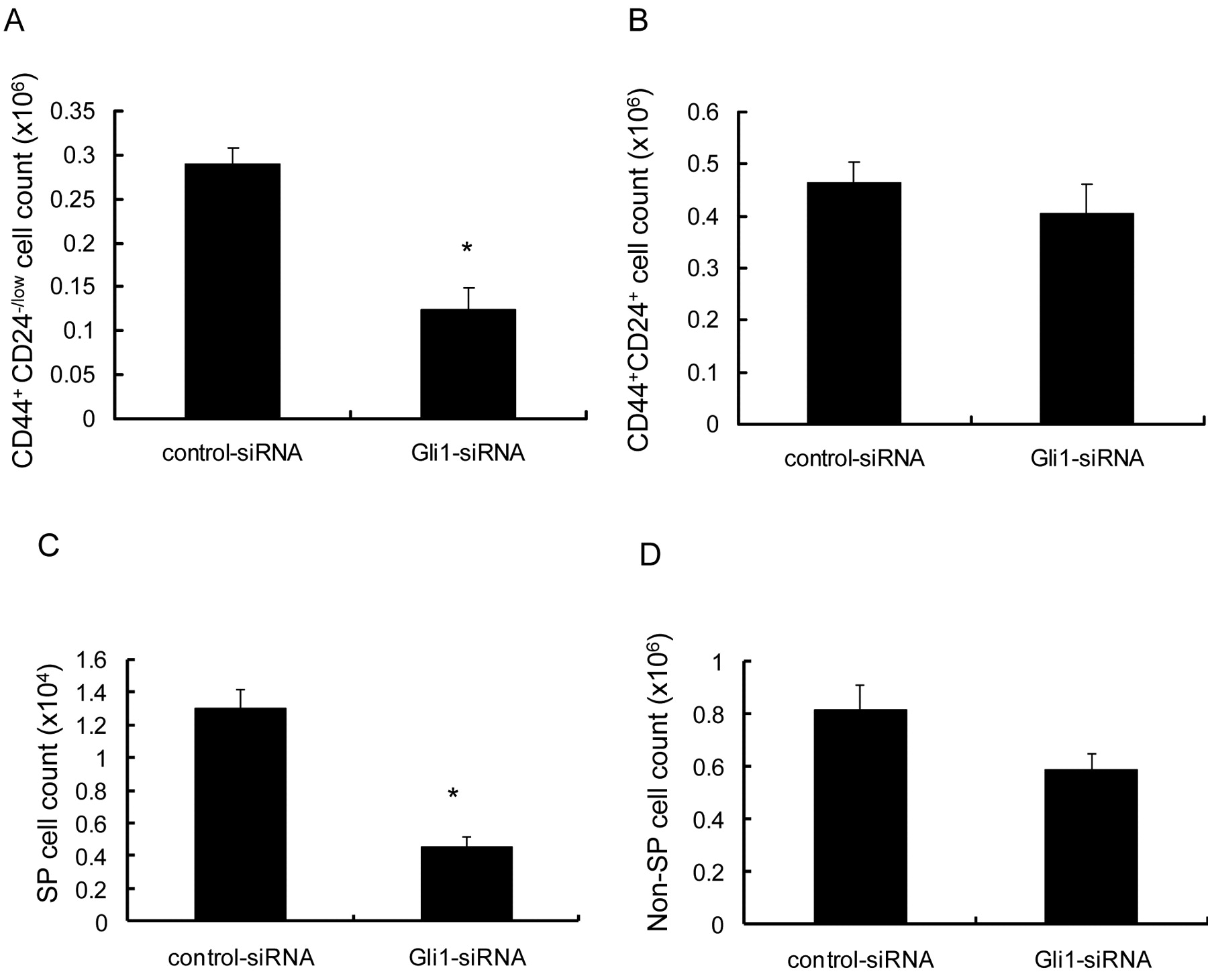
Gli1 plays significant roles in maintaining the CD44+/CD24-/low population and the SP. The significant role of Hh activity in the maintenance of CD44+/CD24-/low cells and SP cells was examined using an siRNA deduced from the Gli1 mRNA sequence. MCF-7 cells were incubated with the Gli1 siRNA and the cell numbers of CD44+/CD24-/low populations, CD44+/CD24+ populations, SP and non-SP were examined by flow cytometry. The number of CD44+/CD24-/low cells decreased significantly (p=0.011) following incubation with the Gli1 siRNA, whereas the number of CD44+/CD24+ cells did not (Figure 7A, B). The Gli1 siRNA also inhibited the proliferation of SP cells, but not non-SP cells. These data indicated that the Hh signaling pathway and especially Gli1 are essential for the proliferation of tumorigenic breast cancer cells.

CD44+/CD24-/low cells and SP cells are resistant to paclitaxel. MCF-7 cells were incubated with 0, 10 and 20 nM paclitaxel for 72 hours, counted and analyzed by flow cytometry. Columns, mean; bars, SD; *p<0.05, statistically significant difference. A, The number of non-SP cells was reduced by paclitaxel in a concentration-dependent manner. B, The number of SP cells did not change significantly in the presence of paclitaxel. C, The ratio of SP cells to all cells increased. D, The number of CD44+/CD24+ cells was decreased by Paclitaxel in a concentration-dependent manner. E, The number of CD44+/CD24-/low cells did not change significantly in the presence of paclitaxel. F, The ratio of CD44+/CD24-/low cells to all cells increased.
CD44+/CD24-/low population cells are more tumorigenic than CD44+/CD24+ population cells. To confirm the higher tumorigenicity of the CD44+/CD24-/low cells, purified CD44+/CD24-/low and CD44+/CD24high population MCF-7 cells were injected into the NOD/SCID mouse xenograft model. When injected into NOD/SCID mice, 2×103 CD44+/CD24-/low cells could not generate tumors, but 2×104 CD44+/CD24-/low MCF-7 cells were able generate tumors with 100% (8 of 8) efficiency (Table I). By contrast, no tumors were observed when 2×104 CD44+/CD24high cells were injected (0 of 8) (Table I). This result suggested that MCF-7 CD44+/CD24-/low cells are probably more tumorigenic than CD44+/CD24high cells in vivo.
Discussion
In the present study, it was discovered that the SP and CD44+/CD24-/low populations partially overlap. SP cells and CD44+/CD24-/low cells were resistant to paclitaxel. On the other hand, components of the Hh signaling pathway, namely Shh and Gli1, were more highly expressed in CD44+/CD24-/low cells and SP cells, at both mRNA and protein levels, than in other populations. Inhibition of Hh signaling activity suppressed the proliferation of both populations. The significance of the Hh signaling pathway in the proliferation of these cells was demonstrated more precisely by the use of a Gli1 siRNA.

Components of the Hh signaling pathway were highly expressed in CD44+/CD24-/low cells and SP cells. Columns, mean; bars, SD; *p<0.05, statistically significant difference from the control group. A, Relative mRNA levels of Shh and Gli1 in CD44+/CD24+ cells and CD44+/CD24-/low cells. B, Relative mRNA levels of Shh and Gli1 in unsorted cells and SP cells. C, Shh (a-f) and Gli1 (g-l) were also highly expressed in CD44+/CD24-/low cells and SP cells, as detected by fluorescence immunostaining. Sorted CD44+/CD24+ cells (a, g), CD44+/CD24-/low cells (b, h) and also unsorted cells (c, i) and SP (d, j) cells were stained for Shh (a-f) and Gli1 (g-l) using a monoclonal antibody. The human colonic adenocarcinoma cell line DLD-1 cells (e-k) was used as negative control for Hh signaling activities, and the human pancreatic ductal adenocarcinoma cell line AsPC-1 cells (f-l) was used as positive control for Hh signaling activities. Original magnifications ×100.
The SP and the CD44+CD24-/low subpopulation are two major candidate sources of CSCs in breast cancer tissues; these two subpopulations have been reported separately to be more enriched in tumorigenic cells than other populations and to have heterogeneous differentiation abilities (14, 15). Furthermore, using the breast cancer cell line MCF-7, it was possible to confirm that the CD44+/CD24-/low population is also more tumorigenic than the CD44+/CD24high population in the NOD/SCID mouse xenograft model. Further, Patrawala et al. showed that the tumorigenicity of the SP of MCF-7 cells is higher than that of non-SP cells in vivo (15). A relationship was found between these two major CSC candidates as the CD44+/CD24-/low population and the SP overlapped: the CD44+/CD24-/low population comprised 38% SP cells, and the SP comprised 65% CD44+/CD24-/low cells. By contrast, 35% of MCF-7 cells were CD44+/CD24-/low cells and 1.8% of MCF-7 cells were SP cells. As described above, it was concluded that the CD44+/CD24-/low population and the SP overlap considerably. The differences between the overlapping population of CD44+/CD24-/low and SP cells and other populations are currently being investigated.
MCF-7 CD44+/CD24-/low cells are enriched with tumorigenic cells.

Suppression of Hh activity in CD44+/CD24-/low cells and SP cells leads to growth suppression of cancer cells. Columns, mean; bars, SD; *p<0.05, statistically significant difference from the control group. Sorted CD44+/CD24-/low cells and SP cells were incubated with 10 and 30 μM cyclopamine for 72 hours and analyzed by flow cytometry. A, The total number of MCF-7 cells was reduced by incubating them with cyclopamine. B, C, The ratio of CD44+/CD24-/low cells to all cells further decreased significantly, and the number of CD44+/CD24-/low cells decreased dramatically. D, E, The ratio of SP cells to all cells and the number of SP cells were also dramatically reduced by cyclopamine.
Stem cells were first isolated based on their high Hoechst 33342 efflux ability, which results from their high expression of multi-drug resistant genes (18). SP cells also express high levels of ABCG2 (15). However, the drug-resistance ability of the CD44+/CD24-/low population has not been explored to date. It is hereby reported that the number of cells in the CD44+/CD24-/low population did not decrease in the presence of paclitaxel and that the ratio of CD44+/CD24-/low cells to all cells increased. These data are consistent with and further confirm the fact that the CD44+/CD24-/low population overlaps with the SP, and show that this overlapping cell population overexpresses multi-drug resistance genes.
However, one contradiction remains. When CD44+/CD24-/low cells are sorted first and SP cells second, the proportion of MCF-7 cells representing the overlapping population is 13%; however, this result appears to contradict the finding that only 1.8% of MCF-7 cells were SP cells. One possible explanation for this discrepancy is that the stimulation of CD44 by the antibodies used in flow cytometry experiments transformed non-SP cancer cells into SP cells. This is the next significant issue to be investigated in the search for the roles of CD44 signaling in CSCs.
Sims-Mourtada et al. reported that activation of the Hh signaling pathway promotes multiple drug resistance by regulating drug transport (29). Consistent with the above study, it is hereby reported that both SP cells and CD44+/CD24-/low cells express high levels of components of the Hh signaling pathway. Furthermore, we determined an additional significance of the Hh signaling pathway in these cells, in that their proliferation was suppressed by an inhibitor of the Hh signaling pathway. Finally, an siRNA against Gli1, a trans-activator of the Hh signaling pathway, suppressed the proliferation of CD44+/CD24-/low and SP cells. These data indicate that the Hh signaling pathway represents a candidate molecular target for CSC-controlling therapies.
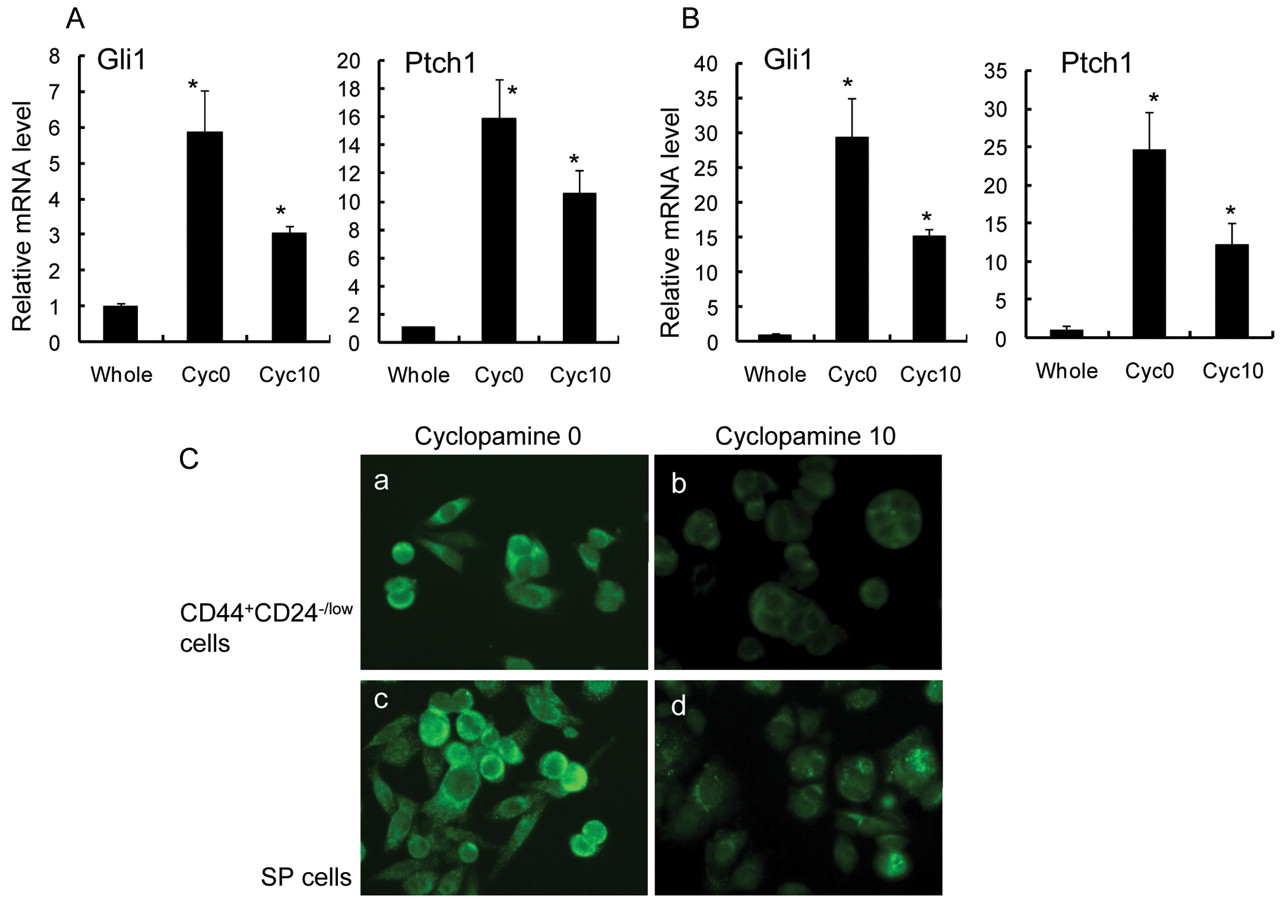
Gli1 and Ptch1 mRNA expressions in CD44+/CD24-/low cells and SP cells were clearly suppressed after incubation with cyclopamine. Columns, mean; bars, SD; *p<0.05, statistically significant difference from the control group (unsorted, whole MCF-7 cells). A, Sorted CD44+/CD24-/low cells were incubated with 0 and 10 μM cyclopamine for 72 hours. The expression levels of Gli1 and Ptch1 mRNAs in CD44+/CD24-/low cells were examined by real-time RT-PCR. ‘Total cells’ indicates all MCF-7 cells. B, Sorted SP cells were incubated with 0 and 10 μM cyclopamine for 72 hours. The expression levels of Gli1 and Ptch1 in SP cells were examined by real-time RT-PCR. ‘Total cells’ indicates all MCF-7 cells. C, Expression of Gli1 was also suppressed by cyclopamine, as detected by fluorescence immunostaining. Sorted CD44+/CD24-/low cells and SP cells incubated with 0 μM (a, c) and 10 μM (b, d) cyclopamine for 72 hours were stained for Gli1 using a monoclonal antibody. Original magnifications ×100.
Thus, it is concluded that the Hh signaling pathway, especially Gli1, is essential for the proliferation of the tumorigenic population of breast cancer cells, and that this pathway represents a new candidate target of cancer stem cell therapies for breast cancer.

Gli1 plays significant roles in maintaining the CD44+/CD24-/low population and the SP. Columns, mean; bars, SD; *p<0.05, statistically significant difference from the control group. CD44+/CD24-/low cells (A, B) and SP cells (C, D) were incubated with Gli1 siRNA and the cell number was examined by flow cytometry. The numbers of CD44+/CD24-/low cells and SP cells decreased significantly following incubation with a Gli1 siRNA, while the numbers of CD44+/CD24+ cells and non-SP cells did not.
Acknowledgements
General Scientific Research Grants 18390350, 18591439, and 19390337, the Ministry of Education, Culture, Sports, and Technology of Japan.
- Received January 5, 2009.
- Revision received February 26, 2009.
- Accepted April 6, 2009.
- Copyright© 2009 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Activation of hedgehog signaling associates with early disease progression in chronic lymphocytic leukemia
- miR-200c suppresses stemness and increases cellular sensitivity to trastuzumab in HER2+ breast cancer
- CD24 Modulates Chemosensitivity of MCF-7 Breast Cancer Cells
- Targeting Cellular Signaling Pathways in Breast Cancer Stem Cells and its Implication for Cancer Treatment
- Excessive Hyaluronan Production Promotes Acquisition of Cancer Stem Cell Signatures through the Coordinated Regulation of Twist and the Transforming Growth Factor {beta} (TGF-{beta})-Snail Signaling Axis
- Hedgehog Signaling: Networking to Nurture a Promalignant Tumor Microenvironment
- Global Profiling of Signaling Networks: Study of Breast Cancer Stem Cells and Potential Regulation
- PTEN Loss Accelerates KrasG12D-Induced Pancreatic Cancer Development
- Frequent Deregulations in the Hedgehog Signaling Network and Cross-Talks with the Epidermal Growth Factor Receptor Pathway Involved in Cancer Progression and Targeted Therapies
- Inhibition of Wnt Signaling Pathway Decreases Chemotherapy-resistant Side-population Colon Cancer Cells