


Abstract
Different research groups have recently described a proton [H+]-related mechanism underlying the initiation and progression of the neoplastic process in which all cancer cells and tissues, regardless of their origin and genetic background, have a pivotal energetic and homeostatic disturbance of their metabolism that is completely different from all normal tissues: an aberrant regulation of hydrogen ion dynamics leading to a reversal of the pH gradient in cancer cells and tissues (∆pHi to ∆pHe) as compared to normal tissue pH gradients. This basic specific abnormality of the relationship between the intracellular and the extracellular proton dynamics, a phenomenon that is increasingly considered to be one of the most differential hallmarks of cancer, has led to the formation of a unifying thermodynamic view of cancer research that embraces cancer fields from etiopathogenesis, cancer cell metabolism, multiple drug resistance (MDR), neovascularization and the metastatatic process to selective apoptosis, cancer chemotherapy and even the spontaneous regression of cancer (SRC). This reversed proton gradient is driven by a series of proton export mechanisms that underlie the initiation and progression of the neoplastic process. This means that therapeutic targeting of the transporters that are active in cancer cells could be selective for malignancy and is likely to open new pathways towards the development of more effective and less toxic therapeutic measures for all malignant diseases. Here we review the transporters involved in driving the reversed proton gradient and their specific inhibitors.
- Cancer etiopathogenesis
- pH and cancer
- multiple drug resistance (MDR)
- proton transport in cancer
- cancer angiogenesis
- cancer strategy and treatment
- review
H+-related Etiopathogenic and Therapeutic Aspects of Cancer
The induction and/or maintenance of intracellular alkalinization and its subsquent extracellular, interstitial acidosis (1-6) on intratumoral dynamics have been repeatedly implicated as playing an essential, direct and pivotal role both in cell transformation as well as in the active progression and maintenance of the neoplastic process (1, 4, 7). Indeed, this specific and pathological reversal of the pH gradient in cancer cells and tissues (∆pHi to ∆pHe) compared to the normal tissue pH gradient is considered to be one of the main characteristics defining the molecular energetics of tumors, regardless of their pathology and genetic origins (3). There is now ample data demonstrating that the aberrant regulation of hydrogen ion dynamics leading to this reversed proton gradient is driven by a series of proton export mechanisms that underlie the initiation and progression of the neoplastic process (4). These transporters and some of their most easily available inhibitors are depicted in Figure 1. While the hyperactivity of the Na+/H+ exchanger isoform 1 (NHE1) is a critical component in the up-regulation of proton extrusion and in its secondary activation of cell transformation, proliferation, motility, and invasion of cancer cells derived from a wide array of tissues (3-5), it is not the only plasma membrane-bound membrane transporter/enzyme responsible for cytosolic alkalinization of the tumor cell and acidification of the extracellular space. Additionally, the vacuolar H+-ATPases (8, 9), the H+/Cl- symporter (10), the monocarboxylate transporter (MCT, mainly MCT1) (also known as the lactate-proton symporter) (11-13), the Na+-dependent Cl-/HCO3- exchanger (2, 7), ATP synthase (14-16) and, perhaps the Na+/K+-ATPase (17, 18) can also play an important role in proton extrusion, pHi abnormalities and tumor interstitial acidification in different human malignancies (4) (Figure 1). Finally, studies on tumor microenvironment pH have shown clear evidence that some carbonic anhydrase (CAs) isozymes, mainly CAII, CAIX and CAXII, are overexpressed in various types of human tumors, an up-regulation that is inversely related to prognosis; and they also make a significant contribution to the extracellular acidity, which is one of the main functional hallmarks of invasive cancer, therefore representing promising targets for novel anticancer therapies (2, 7, 10, 19-21).
These findings suggest that the targeting of proton transporters may be used to trigger selective cancer cell death through the induction of low pHi-mediated apoptosis (5, 15, 19) (Figure 2). Tumor cell proliferation is abolished through the concerted inhibition of NHE1 and Cl-/HCO3- exchangers (18). Similarly, while Cl-/HCO3- exchanger inhibition alone is insufficient to induce apoptosis in breast cancer cells, the simultaneous inhibition of the NHE1 and H+-ATPase induces apoptosis through their concurrent effects on lowering pHi (8). The failure of tumor cells to die following chemotherapeutic treatment also often appears to be highly dependent on their resistance to undergo intracellular acidification, a low intracellular pHi homeostatic situation that is apparently necessary as a prior and early condition that allows cancer cells to engage in a tumor-specific apoptotic process (2, 22-26) (Figure 2). In summary, the final aim is to target this specific aspect of cancer cell metabolism based on the H+-dependent thermodynamic advantages that malignant cells possess as compared to their normal counterparts. The exploitation of such differences in selective cancer therapeutics as chemotherapy adjuvants is a possible successful strategy that could decrease chemotherapy dosages while at the same time increasing therapeutic specificity and effectiveness regardless of tumor type and origin.
The Thermodynamic Neostrategy of Cancer Cells and Tissues: Order within Chaos
During and after neoplastic transformation, a thermodynamically advantageous reversal of the previously normal situation takes place, namely, the reversal of the transmembrane H+-gradient (alkaline inside, acidic outside), a specific feature described only in malignant disease. The main mechanism of this reversal is an intracellular alkalinization mediated by the systemic extrusion of H+ by the different proton transporters (PTs) described above, while the chloride bicarbonate exchanger brings in a bicarbonate anion exchange for a chloride anion to neutralize protons inside the cell (Figure 1). However, since not all cancer cells necessarily have the same transporters elevated at the same time, it appears that in order to maintain the abnormal cellular alkalinity, when one transporter is inhibited others can become up-regulated. The observed consequences of this initial cellular acid-base energetic change demonstrates that one of the main purposes of the biochemistry and metabolism of the concerted, dynamic, energetic defensive systems of cancer cells and tissues is to have the different transmembrane proton transport mechanisms working, at least when required by damaging microenvironmental conditions (4), to first create and then maintain a cascade of electrochemical changes and events leading to tumor development, local growth and invasion, the activation of the metastatic process and, simultaneously, resistance to treatment (3, 26-28).
These mechanisms driven by the loss of the normal acid-base homeostatic balance of the cell (initial and specific cause for cell transformation) have been summarized in Figure 1, and they are: A) Maintenance of a normal to elevated intracellular pH even under the circumstances of the metabolic microenvironment of cancer cells (interstitial acidosis, lack of blood supply, low O2 conditions) in order to protect themselves from low pHi-mediated apoptosis, at the same time that they initiate an unregulated and thermodynamically beneficial proliferative and invasive state; B) The establishment of a self-defensive, anti-apoptotic strategy mediated through different anti-acidifying mechanisms such as hyperactivity of the different membrane-bound proton extrusion transporters, inactivation of Bcl-2, Bcl-xl, and destabilization of p53 (28-30); C) These concerted dynamic changes are based upon the advantageous utilization of a H+-gradient reversal function as an anti-chemotherapeutic shield involved in multiple drug resistance (MDR) and in the development of newly resistant subpopulations of tumor cells (3); D) The above mechanisms lead to secondary acidification of the interstitial component of tumors, in either low or normal O2 conditions, which is key to the onset of local invasion and to the activation and maintenance of the metastatic process by increasing the expression of a wide array of positive angiogenic factors (e.g. HIF-1, VEGF) (6, 7), while the extracellular acidification of tumors creates even further resistance to chemotherapy, radiation-induced apoptosis and hyperthermia (28-31) (Figure 1).
(A) Self-protection against the caustic extracellular tumor microenvironment. While non-transformed cells and tissues die under conditions of extracellular acidosis, the multiple-transporter strategy allows malignant cells of diverse origins to defend themselves from any acidic and/or therapeutic and/or apoptotic attack by taking advantage of a concerted system of membrane-bound ionic transporters whose main role is to extrude hydrogen ions from the cell. This allows transformed cells of all genetic origins to first survive and then multiply under these extremely difficult environmental circumstances. This tumor-specific metabolic condition suggests a possible therapeutic solution: it is this same highly pathological and specific pH gradient reversal of all cancer cells and tumors which becomes the key factor that offers the opportunity to target it as one of the few, if not the only truly differential characteristic that separates all malignant tissues from all normal ones. This would be to attempt to selectively induce a cancer cell self-poisoning through diverse low pHi-related therapeutic measures (Figure 1) (32-33). Since no cancer cell can survive for long with pHi conditions below a certain acid threshold (22), successful therapeutic interventions targeting this H+-mediated gradient reversal through the concerted utilization of proton transport inhibitors (PTIs) of the different families can become a key therapeutic strategy to selectively trigger the apoptotic process in malignant cells and tissues (2, 7, 22) (Figure 2). Unfortunately, with very few exceptions, this approach is still to be tested in clinical oncology (34) (Figure 1).

Homeostatic and dysregulated pH-control systems in cancer cells. Specific targets for the different families of proton transport inhibitors as pHi-lowering anticancer agents (Nos. 1, 2 3, 4, 5 and 6). Mechanisms that induce intracellular alkalinisation as the key factor in cell transformation and progression with its secondary abnormalities. Secondary pHi-dependent extracellular acidification, pH-gradient reversal and hypoxia as triggers for the metastatic process. Six targets for inhibition of proton extrusion of cancer cells as targets for metabolically-directed anticancer treatment and examples of drugs of the different proton transport inhibitors at the sites of their activity. Abbreviations: NHE1: Na+/H+ exchanger: HMA: 5-(N,N-hexamethylene)-amiloride, DMA: 5-(N,N-dimethyl)amiloride; HIF-1: hypoxia-inducible factor; MCT1: monocarboxylate transporter or H+-lactate co-transporter; CAIX: carbonic anhydrase IX; V-H+-ATPase: vacuolar H+-ATPase; VEGF: vasoendothelial growth factor; UKt-PA: urokinase-type plasminogen activator; P-gp: P-glycoprotein; MDR: multiple drug resistance; pHi: intracellular pH; pHe: extracellular/interstitial tumoral pH (For further details, see text).
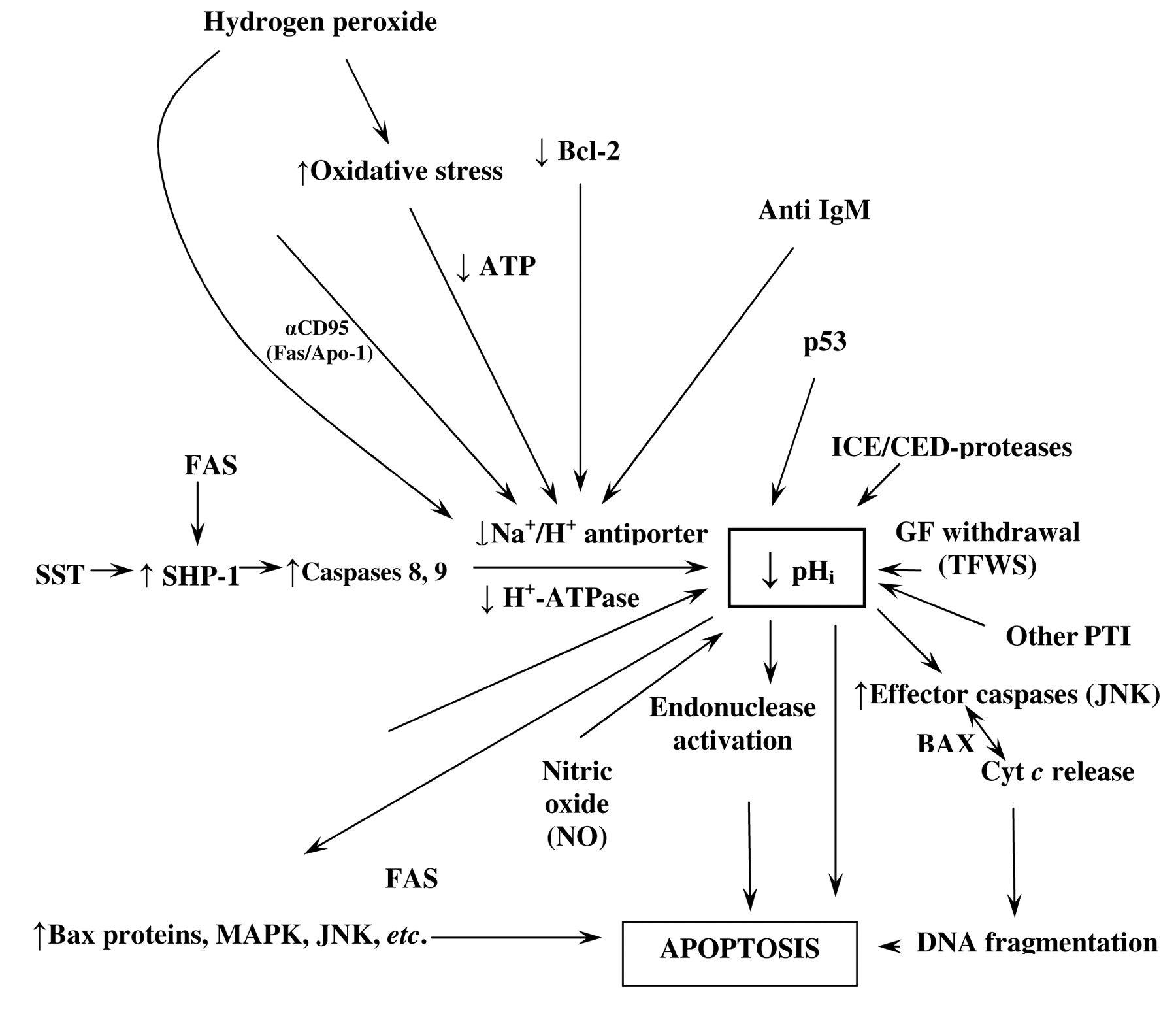
(B) pHi and selective apoptosis in cancer. Figure 2 depicts a map of the intracellular dynamics of the hydrogen ion and the main factors leading to the stimulation of low pHi-induced apoptosis. This figure represents an attempt to integrate the different approaches to trigger pro-apoptotic mechanisms, new therapeutic targets and more selective anticancer drugs. A series of studies using different chemotherapeutic substances in a variety of tumor cells have reported that cytosolic acidification is a very early event in the onset of malignant cell apoptosis (35-37). The induction of an intracellular acid environment has been reported to trigger the onset of apoptosis of leukemic cells by up-regulating the expression of Bax protein expression, which is pro-apoptotic. This seems to be mediated by the activation of interleukin-1β-converting enzyme (ICE/caspase-1) or the apoptosis-effector protease CPP32 (Caspase-3), irreversibly leading to acid stress-induced apoptosis, and thus to the control of cell proliferation and arrest of tumor growth (Figure 2). Inhibition of the NHE1 plays a fundamental role in paclitaxel-induced apoptosis of breast cancer cells and this is synergistically potentiated by inhibition of the NHE1 with the amiloride analog, 5-(N,N-dimethyl)amiloride (DMA), while the recently developed, potent inhibitor of the NHE1, cariporide (HOE-642) seems to induce a similar effect (36). Since NHE1 inhibition reduces transformed cell pHi well below parental cell values, and this intracellular pH decrease does not show a significant effect on normal cells, this indicates a certain degree of therapeutic selectivity and specificity for at least certain NHE1 inhibitors in malignancy (22), even more so since proton transporters are differently expressed in normal and tumor tissues (2). Finally, the outstanding results of Rich et al. (22) in different kinds of leukemic cells with the potent amiloride derivative inhibitor of NHE1, 5-(N,N-hexamethylene)-amiloride (HMA), which reduces the intracellular pH well below the acid-base survival threshold, leads to consider that inducing a low pHi-mediated apoptosis might become a selective therapeutic modality for many different cancer cells and tissues if these results could be translated to clinical therapeutics.
Similarly, to inhibit the MCT, lonidamine and cinnamate have been used (12). Recently, AstraZeneca developed even more exciting agents, in that they can inhibit MCT in the nanomolar range and are more specific (38; Wahl unpublished results). MCT levels have been found to be high in neuroblastoma cells (12) and in melanoma cells exposed to a low pHe (11). In neuroblastoma cells, the gene for the MCT (SLC16A1) is amplified and not only did cell death occur as a function of pHe in vitro, but the correlation between high levels of MCT and poor prognosis was also found in biopsy specimens from children with neuroblastoma, a pediatric malignancy with a very high mortality rate (12). Parallel results are obtained when other proton transporters, such as CAIX, are considered (20, 21).
(C) pHi, pHe and MDR. A direct cause effect relationship among the degree of MDR and the elevation of tumor pHi has been recognized by different groups studying the dynamic interrelationships between cell pHi and MDR (2, 4, 9, 34). High pHi, mediated either by overexpression/activity of the NHE1 and/or other proton-extruding mechanisms such as V-ATPases, MCTs and carbonic anhydrases (CAs) have been found to be responsible for cisplatin resistance (14, 35) and, similarly, to contribute to the onset and/or maintenance of MDR, so protecting against tumor cell death from anticancer drugs (9, 20, 21, 35). Furthermore, drugs such as adriamycin, cisplatinum, paclitaxel and camptothecin have been shown to be unable to induce apoptosis under non-acidified cellular conditions (24, 25, 36) and, indeed, resistance to several anticancer drugs such as camptothecin, vinblastine, adriamycin and etoposide has been shown to be dependent on overexpression of different proton transporters and/or intracellular alkalinization (4, 5, 24, 25, 34). Recently, third-generation camptothecin analoges have been developed that are more active at low pH (39). This design should lead to more selectivity and less toxicity of this chemotherapeutic agent. In this context, specific H+-ATPase inhibitors, such as bafilomycin A1, salcylihalamide, lobatamides and oximidines have been also considered as potential anticancer agents and MDR-reversal agents (9), in a similar way to CAIX inhibitors such as acetazolamide (Figure 1) (2, 21).
The fact that cells with an active MDR transporter show cytoplasmatic alkalinization has led some authors to conclude that P-glycoprotein can be mainly considered as a proton extrusion pump (26, 27, 40). However, P-glycoprotein (P-gp) activity is stimulated by interstitial acidification secondary to the abnormal H+-dynamics of cancer tissues (28) (Figure 1) and, indeed, the therapeutic failure to induce cytoplasmic acidification has been proposed as the main underlying factor for MDR because of resistance to the induction of therapeutic apoptosis in both normal or slightly alkaline and highly alkaline cancer cells (4, 29, 33, 41) (Figure 2). Thus, in many instances it seems that MDR can be attributed to the failure to induce intracellular acidification by compounds such as chloroquine, imidazol, glutathione, apart from overexpression/activity of proton transporters (2, 9, 17). Finally, the MDR-promoting effects of the Bcl-2 family of proteins, as well as a dysfunctional p53, which also contribute to pro-carcinogenic and antiapoptotic effects, have also been shown to be dependent on their ability to maintain a sufficiently elevated intracellular cell pH in order to avoid therapeutic apoptosis (29, 30, 35).
All these findings are further corroborated by the fact that a large variety of MDR modifiers known to be able to revert resistance to chemotherapeutic drugs (e.g. verapamil, amiodarone, bafilomyicin A1, cyclosporine A, tamoxifen, DIDS, nigericin and edelfosine), have all been reported to exert their cellular effects, at least in part, through pHi-acidifying mechanisms (31). A decrease in pHi has been shown to sensitize cancer cells of diverse origins to apoptosis, chemotherapy and hyperthermia, or to induce apoptosis by themselves (4, 24, 25). Indeed, a reversal of MDR can be obtained by the pH-lowering effects of amiloride and/or its analogs in a variety of situations (23). In summary, a selective and concerted role for PTIs as chemotherapy adjuvants in MDR, as well as selective anticancer agents on their own, could well be a very successful strategy. This would decrease chemotherapy dosages and toxicity while at the same time increase therapeutic specificity and effectiveness regardless of tumor type and origin (26).
(D) Relationships between hydrogen ion dynamics, malignant neovascularization and the metastatic process. Neovascular growth and metastasis are direct consequences of the hostile environment of low extracellular pH as well as of low interstitial pO2 (3). Indeed, the high pHi-low pHe-proton gradient reversal factor by itself can induce vascular endothelial growth factor (VEGF) production, without the need of any other structural intermediate such as hypoxia-inducible factor 1 (HIF-1), and which, at the same time, can be inhibited by lowering intracellular pH and/or collapsing the proton gradient reversal with amiloride (42, 43) (Figure 1). This H+-gradient reversal has also been shown to induce not only the expression of VEGF but also of insulin-like growth factor 1 receptor (IGF1R), platelet-derived growth factor β-receptor, interleukin 8 and metalloproteases (33) (Figure 1). Recent research trends have mainly focused on tumoral hypoxia as a source of VEGF and has emphasized its role in the metastatic process (44-46). However, the proven role of relative hypoxia as a direct etiological factor in cell malignant transformation, as initially proposed by Warburg - what was previously called by us the Warburg-Goldblatt effect - still needs to be taken into account as an etiological factor of its own in cell malignant transformation in a manner similar to high pHi (low intracellular H+-concentration functionally mimicking low pO2, or para-hypoxia) (31).

Intracellular signalling factors and mechanisms targeting pHi and the Na+/H+ exchanger in apoptosis. Factors that induce apoptosis through intracellular acidification as its common final pathway. This integrated and homeostatic pH-related perspective can help to foretell pro-apoptotic and anti-apoptotic factors in order to find synergistic therapies and potential antagonisms (MDR) in anticancer treatment. Abbreviations: ↑: Stimulation; ↓: inhibition; SST: somatostatin; SHP1: protein tyrosine phosphatase; MDR: multiple drug resistance; GFs: growth factors; Cyt C: cytochrome C; NO: nitric oxide. TFWS: trophic factor withdrawal syndrome; αCD95 (Fas/Apo-1) death receptor; JNK: Jun-terminal kinase; MAPK: mitogen-activated protein kinase. (For further details, see text and refs. 27, 50); PTI: proton transport inhibitors; ICE: interleukin-1β-converting enzyme.
The role of NHE1 activity and/or an abnormally increased pHi in stimulating different steps of the metastatic process have been reviewed elsewhere (3, 4) (Figure 1). The activity of a significant number of proangiogenic factors and oncogenes has been shown to be directly related to NHE1 expression (47), while, on the contrary, a wide array of antiangiogenic drugs inhibit the NHE1 (4, 47). In addition, other pro-metastatic mechanisms are sensitive to inhibitors of NHE1 activity (Figure 1), such as the urokinase-type plasminogen activator (μPA) (3, 41), matrix metalloproteinase (MMP-9) (48) and the cathepsin B-dependent activation of MMP-2 and MMP-9 (49). It has long been known that amiloride can achieve a complete in vivo antimetastatic effect in different transplanted tumors (50). Thus, amiloride and, mainly, its more potent derivatives, have been increasingly considered as a novel, adjuvant and neoadjuvant treatment for cancer in order to reduce tumor growth and increase patient survival (2, 4, 6, 51, 52).
Role of Tumoral pHe in Invasion and Extracellular Protease Action
Multiple studies have strongly supported a pathogenic role of the acidic interstitial pHe of tumors by giving a selective advantage for tumor progression and metastasis. It has been shown to drive large changes in gene expression independently of hypoxia (53, 54) and has also been associated with tumor progression by impacting multiple processes including increased invasion (49, 54-56) and metastasis (57, 58). This can occur directly or through the alteration of the extracellular matrix (ECM) compartment through up-regulation of protease secretion/activation and in an altered tumor-stromal interaction via an inverse stimulation of pro-angiogenic factors paired with impaired immune functions (59) (Figure 1).
Proteolytic ECM remodeling is a prerequisite for the invasive process. Indeed, the proteolytic breakdown of proteins of the ECM is one of the first steps in invasion in primary cancer lesions (60). During invasion, cancer cells use secreted, surface-localized and intracellular cathepsins, serine proteases and MMPs to proteolytically cleave, remove and remodel different types of ECM substrates at the cell surface, including collagens, laminins vitronectin, and fibronectin (61). While tumor-driven extracellular acidification of the tumor pericellular space can directly drive the destruction of the surrounding normal limitrophic tissue (62, 63), a large body of work has demonstrated that the acid pHe of tumors can also indirectly drive ECM proteolysis by increasing protease production and secretion of the active forms of the cathepsin family of proteases, such as cathepsin D (53, 64), cathepsin B (49, 65, 66), cathepsin L (58), MMP-9 (48, 49, 55, 58, 67, 68) and MMP-2 (49, 58). There is evidence demonstrating that NHE1 and its associated extracellular acidification is necessary for the (i) cathepsin B-dependent ECM proteolytic activity and invasion of breast cancer cells in which the ECM receptor, CD44, was activated by hyaluronan (66), and (ii) MMP-9 activation and invasion in non-small lung cancer cells in which alpha1-adrenergic receptor was stimulated by phenylephrine (48). Interestingly, one study observed that the low pHe-driven activation of MMP-9 and MMP-2 was dependent on the up-stream activation of cathepsin B and all three proteases were located on small vesicles shed from the tumor cell (49). This increased secretion and activity of proteases is congruent with the known increased invasive capacity at acid pHe (48, 55, 66).
The acid pHe of tumors has also been shown to alter the interactions between tumor cells and the cells of both the stromal compartment and the immune antitumoral defense system. On the one hand, acidic pHe has been demonstrated to increase the expression and secretion of angiogenesis promoting and metastatic factors such as VEGF (58, 69, 70-72) and interleukin-8 (IL8) (58, 73, 74). On the other hand, there is evidence that the acid component of the tumor microenvironment also directly reduces/impairs the function of the antitumoral immune system, thus contributing to the known in vivo immunosuppression. Exposure to increasingly acidic pHe has been shown to reduce tumor cell-induced cytolytic activity of lymphokine-activated killer (LAK) cells (75), to play a role in down-regulating cytolytic activity of tumor-infiltrating lymphocytes with natural-killer (NK) phenotype (76) and to inhibit the non-major histocompatibility complex (MHC)-restricted cytotoxicity of immunocompetent effector cells (53, 58, 77, 78).
Altogether, these studies indicate that an acidic tumoral interstitial pHe promotes invasion and metastasis by a reciprocal mechanism involving acidity-induced up-regulation of proteolytic enzymes and pro-angiogenic substances together with an acidity-induced down-regulation or impairment of the organisms antitumoral immune defense. One consequence of this situation is that treatment strategies should be aimed by all means at collapsing the intracellular/extracellular H+-gradient inhibiting PTs in order to increase selective intracellular acidification and apoptosis, plus (an apparent paradox) alkalinizing the tumor interstitial space by blocking the mechanisms driving its acidification, while avoiding any therapy that could involve deliberate tumor extracellular acidification.
H+-related Mechanisms in the Spontaneous Regression of Cancer (SRC): Food for Thought
The favorable influence of acidification on complete cancer regression in a wide array of transplanted animal tumors has also been recognized over the years (79, 80). Severe metabolic acidosis induced by some surgical procedures, such as ureterosigmoidostomy, infections and febrile processes, was initially considered to be the main and ultimate underlying mechanism behind some spontaneous regressions of malignant tumors in animals and human beings (31, 79-83). Recently, a graded metabolic acidosis associated with mild renal failure was claimed to reduce, and even reverse, the rates of tumor growth and invasion in cancer patients (84).
Conclusions
Etiopathogenesis-based therapeutics. PTIs as potential and selective anticancer agents in the treatment of human malignant diseases.
While proton research in cancer cannot yet be considered to be within the mainstream of modern oncology research, the increasing evidence accumulated during the last few years points to the fact that the dynamics and metabolism of the hydrogen ion are becoming a subject of growing interest as a potential key target in selective therapeutic intervention in leukemias, solid tumors and other chronic degenerative diseases (33, 85, 86).
The H+-related perspective briefly reviewed here suggests a new paradigm able to encompass an enormous and scattered bulk of information in the main areas of cancer research as has been advanced in recent reports (4, 33). An advantage of such a unified basic approach is the possibility of integrating what previously were considered to be non-interrelated areas of research, in order to translate their data and interrelationships into a more complete and encompassing integrated synthesis and, from there, into clinical therapeutics. Besides improving our basic understanding, this paradigm could stimulate further integrations between biochemical and metabolic cancer research to molecular biology and cancer immunity. The latter relationship is exemplified by the fact that therapeutic cell death in lymphomas depends on NHE1 inhibition by IGM-mediated cell death through intracellular acidification (87).The negative effects of tumoral interstitial acidification in reducing cellular immunity are also well known (75-78).
In spite of the fact that transmembrane H+ gradient reversal appears to be the single most differential molecular characteristic setting apart cancer cells and tissues from normal ones, this feature still remains to be exploited in the treatment of human cancer. Taking into account the theoretical background available and the results of different cell studies, animal experimentation and occasional reports in cancer patients that justify this H+-dependent approach, the situation is difficult to understand. This is probably due to the fact that the most active agents that could revert the abnormal H+-gradient situation, such as the potent amiloride-derivatives like HMA, cariporide and zoniporide (Figure 1), are still waiting to be included in pre-clinical or clinical trials. In this respect, all the available data would suggest the importance of undertaking prospective studies in different human malignancies in order to test the therapeutic, pro-apoptotic, antimetastatic and MDR-overcoming concerted effects of PTI drugs, from the more potent derivatives of the amiloride series to other NHE1, V-ATPase, MCT1, HCO3-/Cl- exchangers and CA inhibitors (4, 7, 9, 11, 12) (Figure 1).
In summary, the main bulk of both seminal and emerging data briefly reviewed in this contribution leads to three consistent conclusions: i) Cell alkalinization constitutes an initial and fundamental event in the transformation process of normal cells and tissues regardless of their origin; ii) The overexpression/activation of a number of membrane-bound proton transporters plays a positive key role in later neoplastic and metastatic progression and a negative role in the host defense mechanisms (e.g. antiangiogenesis, spontaneous regression, therapeutics) by severely disrupting intracellular/extracellular proton gradients and inducing a subsequent alteration in cell thermodynamics; iii) Targeted inhibition of the different proton transporters is a promising area in seeking selective anticancer treatments useful in preventing, retarding, or counteracting the neoplastic process at different levels (Figures 1 and 2). The concerted utilization of PTIs, alone or in combination with other forms of chemotherapy, may prove fundamental in primary, adjuvant and/or neoadjuvant treatment of different solid tumors in humans, as well as in the overcoming of MDR. We have to agree with other leading authors in the field who recently advanced that this pH-targeted therapy approach “will lead to the collapse and massive shrinkage of solid tumors” (7). All the available evidence seems to indicate that this would take place regardless of pathogenic differences or genetic origin.
Acknowledgements
This work was supported by a grant of the Mercedes Castresana Foundation, Vitoria, Spain, and the Italian Association for Cancer Research (l'AIRC). We thank Jose Maria Bakero for informatic assistance. The authors also want to apologize to all investigators who have significantly contributed to the different research fields reviewed here, past and present, but whose work is not specifically cited in this contribution.
- Received February 19, 2009.
- Revision received April 8, 2009.
- Accepted April 23, 2009.
- Copyright© 2009 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
Jump to section
- Article
- Abstract
- H+-related Etiopathogenic and Therapeutic Aspects of Cancer
- The Thermodynamic Neostrategy of Cancer Cells and Tissues: Order within Chaos
- Role of Tumoral pHe in Invasion and Extracellular Protease Action
- H+-related Mechanisms in the Spontaneous Regression of Cancer (SRC): Food for Thought
- Conclusions
- Acknowledgements
- References
- Figures & Data
- Info & Metrics
Related Articles
Cited By...
- Structure-Activity Relationship Study Identifies A Novel Lipophilic Amiloride Derivative That Induces Lysosome-Dependent Cell Death in Therapy-Resistant Breast Cancer Cells
- A novel tescalcin-sodium/hydrogen exchange axis underlying sorafenib resistance in FLT3-ITD+ AML
- Autophagy Is a Protective Mechanism for Human Melanoma Cells under Acidic Stress