


Abstract
Background: 5-Hydroxymethylfurfural (HMF) is produced in large quantities during the processing of food containing carbohydrates and can be metabolised to 5-sulfooxymethylfurfural (SMF), a reactive intermediate that can bind to DNA and cause mutagenic effects. Materials and Methods: Three to six days after birth, multiple intestinal neoplasia (Min/+) mice were given a single subcutaneous injection of either 500 mg/kg body weight (bw) HMF, 25 mg/kg bw SMF or vehicle (0.9 % NaCl), and were euthanised at 12 weeks of age. The number and size of adenomas and flat aberrant crypt foci (ACF) were counted in the intestine. Results: HMF increased the number of small intestinal adenomas (p=0.033), whereas SMF increased the flat ACF number in the large intestine (p=0.025). Treatment with HMF and SMF had no effect on the size of the adenomas. Conclusion: These results show that both HMF and SMF are weak intestinal carcinogens in Min/+ mice.
- 5-Hydroxymethylfurfural
- 5-sulfooxymethylfurfural
- Min/+ mice
- aberrant crypt foci
- intestinal carcinogenesis
Abbreviations: HMF, 5-hydroxymethylfurfural; SMF, 5-sulfooxymethylfurfural; Min/+ mice, multiple intestinal neoplasia mice; ACF, aberrant crypt foci; HMFA, 5-hydroxymethyl-2-furoic acid; SULT, sulfotransferase; Apc, adenomatous polyposis coli gene (murine); FAP, familial adenomatous polyposis; wt, wild type; PAPS, 3′-phosphoadenosine-5′-phosphosulfate.
5-Hydroxymethylfurfural (HMF) is one of many compounds formed during the heating of food containing carbohydrates. It is formed by acid-catalyzed dehydration and in the Maillard reaction from reducing sugars. HMF is a small molecule consisting of three structural components, the furan ring, the allylic alcohol group and the aldehyde group. In humans and rodents, most of the HMF will be converted by oxidation to the metabolite 5-hydroxymethyl-2-furoic acid (HMFA) and excreted (4, 8, 11). However, HMF is also a good substrate for sulfotransferases (SULTs) and HMF might be converted to 5-sulfooxymethylfurfural (SMF), a reactive metabolite that can bind to DNA and cause mutagenic effects in vitro (1, 2). SMF has also been detected in vivo in the blood of mice treated with HMF (3). Although the acute toxicity of HMF is relatively low, HMF appears both to initiate and promote large intestinal ACF in rats (4). In a 2-year study, oral administration of HMF enhanced the incidence of hepatocellular adenomas in female mice, but not in male mice nor in rats (5). In addition, induction of skin papillomas has been described after topical application to mice (6).
Compared to acrylamide, HMF is produced in large quantities and the levels can exceed 1g/kg in several food items, such as dried fruits, caramel products, plum juice and instant coffee powder (7, 8). Although lower levels of HMF are present in bread (7, 9), honey (10) and milk (11), these foods contribute considerably to the HMF intake because they are consumed in high quantities. The average daily intake of HMF in humans is in the range of 30-150 mg (7, 12).
The Min/+ mouse is heterozygous for a mutation in the tumour suppressor gene Apc, which leads to spontaneous development of numerous adenomas, mainly in the small intestine, but also in the large intestine. This mutation is analogous to that seen in humans with the familial adenomatous polyposis (FAP) syndrome, although in humans the adenomas are mainly localized in the large intestine. It has been previously shown that the Min/+ model is most susceptible to intestinal carcinogens early in life (13, 14). This model has been used to test several possible human carcinogens, such as heterocyclic amines (15-17), chlorination by-products in drinking water (18), acrylamide (19) and the experimental carcinogen azoxymethane (AOM) (4, 20, 21). The Min/+ mice also develop flat dysplastic lesions in the large intestine, so called flat ACF, which are believed to be precancerous lesions (22); similar precancerous lesions have been identified in the large intestine of patients with colorectal cancer and FAP (23, 24).
The objective of the present study was to examine whether HMF and SMF could affect intestinal carcinogenesis in Min/+ mice and in their wild type (wt) litter mates.
Materials and Methods
Animals and chemicals. C57BL/6J Min/+ male and C57BL/6J +/+ (wt) female mice were bred at the Norwegian Institute of Public Health, Oslo, Norway. The mice were originally purchased from the Jackson Laboratory (Bar Harbor, ME, USA), and were housed in plastic cages with a 12 h light/dark cycle. There were one female and one male in each cage. Water and feed were given ad libitum. The mice were fed a breeding diet, Harlan Teklad (E) (Harlan Teklad, UK), during gestation and until 4 weeks of age, then the litter was separated from the mother and the Min/+ and wt pups were identified by an allele-specific PCR-assay, as described previously (25). Both genotypes were used in the experiment. Thereafter they were given a standard maintenance diet, SDS RM1 (E) (Special Diets Services Ltd., Witham, UK). HMF of 99% purity was purchased from Sigma-Aldrich Corp. (St.Louis, MO, USA). SMF was synthesized in the laboratory of Albrecht Seidel (Grosshansdorf, Germany), as described previously (3).
Treatment. Pups were given one subcutaneous injection of 500 mg/kg bw HMF in 0.9 % NaCl, 25 mg/kg bw SMF in 0.9 % NaCl or vehicle (0.9% NaCl) 3-6 days after birth (Min/+ mice control n=19, 6 males and 13 females; Min/+ mice treated with HMF n=24, 13 males and 11 females; Min/+ mice treated with SMF n=21, 8 males and 13 females; wt mice control n=16, 9 males and 7 females; wt mice treated with HMF n=18, 7 males and 11 females; wt mice treated with SMF n=15, 8 males and 7 females). Because of the short half-life of SMF in vehicle, 120 min in water at 37°C and approximately 4 min in blood plasma in the mouse in vivo (3), the solutions were made directly before administration. HMF has been shown to be stable in solution for 2 weeks at room temperature (unpublished data, H.R. Glatt, Germany). The mice were euthanised by cervical dislocation at 12 weeks of age.
Scoring of flat ACF and adenomas. The small and large intestine were removed separately and rinsed in cold PBS and slit open longitudinally. The small intestine was then divided into proximal, middle and distal parts. They were then fixed flat between wet (PBS) filter papers for 48 hours in 10% neutral buffered formalin prior to 5 seconds staining with 0.2% methylene blue (George T. Gurr Ltd., London, United Kingdom) dissolved in 10% neutral buffered formalin solution. The small and large intestine was examined by transillumination in an inverse light microscope. The number, size and location of the lesions in the small and large intestine were scored. The sizes of the adenomas were scored as mm2, and the location in the small and large intestine was described as cm distal from the ventricle and the caecum, respectively.
Number of flat ACF in each group and incidence in wild type mice after one subcutaneous injection 3-6 days after birth of HMF (500 mg/kg bw), SMF (25 mg/kg bw), or vehicle (0.9 % NaCl).
Histopathological examination of kidney. Kidneys were removed and put on 10% neutral buffered formalin. After 2-7 days, the kidney was embedded in paraffin wax, cut in thin slices and stained with haematoxylin and eosin.
Statistical analysis. One Way ANOVA on Ranks (SigmaStat software; Jandel Scientific, Erkrath, Germany) was used to test for the differences between the groups of Min/+ mice. For isolation of the groups that differed from the control group the Dunn's Method was used. Z-test was used to test for the differences in proportions between groups of wt mice. A p-value of <0.05 was considered significant.
Results
In the wt litter mates, the number of intestinal lesions was low, with no induction of adenomas in either the small or large intestine. However, treatment with HMF and SMF induced flat ACF in the large intestine of 22 and 13% of the animals, respectively, whereas these lesions were absent in control mice only treated with vehicle (Table I). Nevertheless, the difference between treated and control animals was not statistically significant.
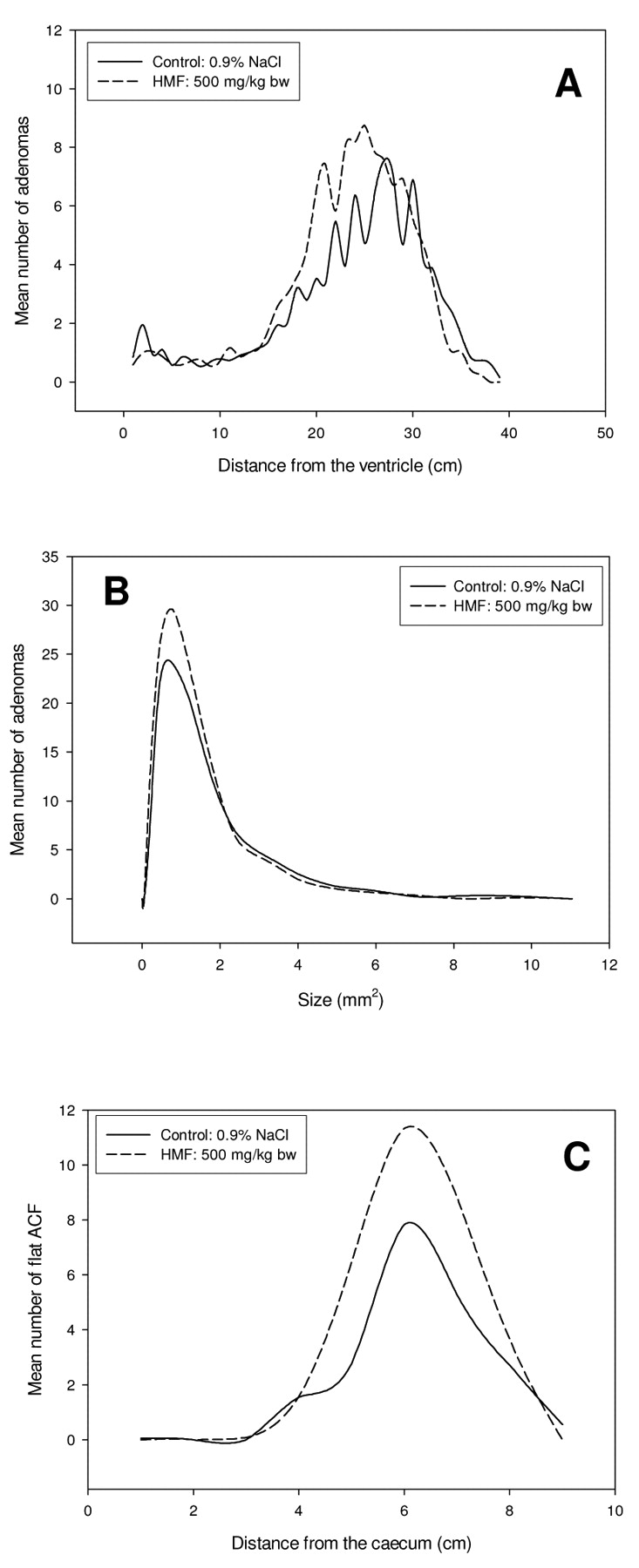
The Min/+ mice treated with HMF had a higher number of adenomas in the small intestine compared to control mice, with a mean of 119 and 102, respectively. A significant increase in the number of adenomas in the small intestine was found in the middle/distal part in the area 15-26 cm from the ventricle (p=0.033) (Figure 1A). This is also the region where the spontaneous adenoma development in Min/+ mice is highest. The increase in small intestinal adenomas due to treatment with HMF was in the range of 0.25-1.2 mm2 in size (Figure 1B). The average size of the adenomas in HMF treated mice was slightly lower compared with the control group, but the difference was not statistically significant. The number of flat ACF in the large intestine increased with a mean of 21 and 32 for control and HMF treated mice, respectively, although the difference did not reach statistical significance (Figure 1C). Treatment with HMF did not give a statistically significant increase in the number of adenomas in the large intestine (data not shown).
SMF treatment increased the number of adenomas in the small intestine with a mean of 102 and 136 for control and SMF treated mice, respectively, although the difference did not reach statistical significance (Figure 2A). The majority of small intestinal adenomas in both groups were in the range of 0.25-1.5 mm2 in size (Figure 2B). The average size of adenomas was slightly lower in SMF-treated animals than in the control group, but the difference was not significant. There was a significant increase in the number of flat ACF in the large intestine of Min/+ mice treated with SMF compared to the control group, in the middle/distal part, in the area 3-7 cm from the caecum (p=0.025) (Figure 2C), the mean number of flat ACF was 21 and 32 for the control and SMF treated mice, respectively. Treatment with SMF did not give a statistical increase in the number of adenomas in the large intestine (data not shown).

Increased development of intestinal lesions in Min/+ mice treated with HMF. Adenoma and flat ACF development in control Min/+ mice versus Min/+ mice treated with 500 mg/kg bw HMF. (A) The mean number of adenomas along the small intestine. (B) The mean size distribution of adenomas in the small intestine. (C) The mean number of flat ACF along the large intestine.

Increased development of intestinal lesions in Min/+ mice treated with SMF. Adenoma and flat ACF development in control Min/+ mice versus Min/+ mice treated with 25 mg/kg bw SMF. (A) The mean number of adenomas along the small intestine. (B) The mean size distribution of adenomas in the small intestine. (C) The mean number of flat ACF along the large intestine.
Since kidney toxicity has been reported after SMF exposure in mice (26), kidneys were sampled for histopathological examination. An abundant acute necrosis was observed in several of the Min/+ and wt mice which had received SMF (Figure 3). There were no signs of gross morphological changes in mice treated with vehicle or HMF.
Discussion
Both HMF and SMF induced an increase in the number of adenomas and flat ACF in the intestine. HMF gave a significant increase in the number of adenomas in the middle/distal part of the small intestine, while treatment with SMF gave a significant increase in the number of flat ACF in the middle/distal part of the large intestine. Additionally, treatment with HMF and SMF induced flat ACF in 22 and 13% of the wt mice, whereas none of the wt control mice developed lesions. These findings are consistent with earlier studies, where 250 mg/kg bw HMF dose given twice by gavage has been shown to induce large intestinal ACF in 45% of the rats (4).
Because of the small size and high water solubility of the molecule, the majority of HMF is eliminated in the urine following oxidation to the corresponding HMFA. It has been shown that through both oral intake and intravenous injections of 14C-labelled HMF, the majority of the radioactivity is found in the kidney, bladder and liver, but also in the intestine and various other organs (27). In another study, where mice were given a single oral dose of 5 to 500 mg/kg bw 14C HMF, the elimination in the faeces was determined to be 8.5-25.5% of the dose (28). Thus, it is likely that a considerable amount of HMF will reach the intestine with a subcutaneous injection, supported by the increase in adenomas and flat ACF number observed after HMF injections.
HMF is converted to SMF in the presence of rat hepatic cytosolic fraction with PAPS, the cofactor for SULTs (2, 6, 29). SMF is much more reactive than HMF, the sulfate group located in the allylic position may be spontaneously cleaved off resulting in an electrophilic compound, the kidneys being the major target for acute toxicity. Due to the high reactivity of SMF towards proteins and DNA it was expected that SMF would induce more adenomas and flat ACF than HMF in Min/+ mice. The results show that SMF does induce more adenomas than HMF in Min/+ mice, with adenoma numbers of 136 and 119, respectively. However, SMF did not induce a higher number of flat ACF than HMF, although due to its toxicity it had to be used at much lower doses than HMF (20-fold). Moreover, the SMF dose could have been further reduced due to the short half-life and/or the compound reacting to surrounding tissue and components in the bloodstream before reaching the intestine. Another important issue is that the cellular uptake of SMF may be low because of its negative charge (3). However, the increase in the number of adenomas and flat ACF in the intestine compared with the control group after treatment with SMF, shows that the compound did reach the intestine.
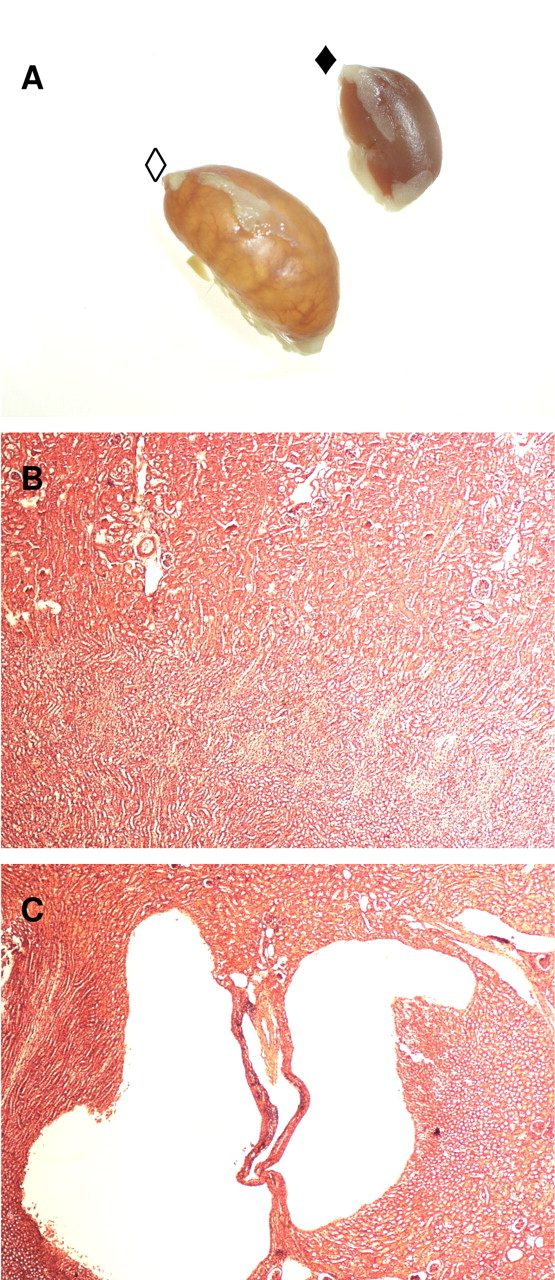
Abundant kidney necrosis in mice treated with SMF. (A) Pictures of kidneys from control mouse (◆) and mouse treated with SMF (◇). (B) H&E stained section of kidney from mouse treated with vehicle. (C) H&E stained section of kidney from mouse treated with SMF.
The control Min/+ mice in this study spontaneously developed a high number of small intestinal adenomas with a mean of 102, whereas these mice normally develop 30-60 (30). The effect of HMF and SMF might have been camouflaged by the high background of adenomas. In addition, there were high individual differences concerning both the adenoma and flat ACF number, which resulted in low statistical power. This may explain why the increase in adenoma and flat ACF number after treatment only reached statistical significance in parts of the small and large intestine.
A slight reduction in adenoma size was observed in the small intestine. It appears that HMF and SMF lead to an increase in the number of small adenomas, reducing the mean value for adenoma size in the two treatment groups. This might indicate either an induction at a later point in time and/or slower growth than spontaneously occurring adenomas.
Since HMF is found at high levels in numerous food items and humans are exposed on daily basis, it would have been interesting to see if repeated subcutaneous injections would have increased the effect. Mice, both wt and Min/+, who received an additional dose of 500 mg/kg bw HMF, developed strong convulsions and clear apnoea after a few minutes. For this reason, the experiment was terminated. It has been shown that the aldehyde group of HMF is able to bind to proteins (28) and might therefore serve as a hapten upon subcutaneous injection, the observed reaction could possibly be an anaphylactic shock caused by an allergic reaction. With oral treatment of the animals, this problem would likely have been avoided.
HMF has been shown to be negative in several conventional genotoxicity tests (2, 6, 8, 29), but these test systems lack the enzymes necessary for HMF activation. In fact, HMF was shown to be mutagenic in tests using Salmonella typhimurium together with rat hepatic cytosol enriched with 3′-phosphoadenosine-5′-phosphosulfate (PAPS) (29), indicating that SULT is essential for the activation of HMF. In addition, HMF induced mutations and sister chromatid exchange in human SULT1A1 expressing Salmonella typhimurium and V79 cells, respectively, but not in the parental cells (1).
It has been hypothesized that humans may be more sensitive to HMF exposure than rodents because they express SULTs in several different tissues, while in rodents most SULTs are mainly expressed in the liver. For example, the human gut is a rich source of many different SULTs (31), whereas this enzyme class is very low in the gut compared to the liver in rats (32).
Human intake of HMF is 103-105 higher than for other hazardous compounds formed during heat treatment of food, such as acrylamide, furan, heterocyclic amines or polycyclic aromatic hydrocarbons. Although the acute and chronic toxicity of HMF is very low, it has been shown that HMF can be activated by SULTs, and it has been hereby demonstrated that both HMF and SMF have a weak tumourigenic activity in the intestine of Min/+ mice even after a single subcutaneous injection. Thus, there is a concern for the mutagenic and carcinogenic capabilities of HMF, and further studies which take into consideration the difference in metabolism between humans and rodents are necessary.
Acknowledgements
We thank Ewa Andruchow, Hege Hjertholm, Andre Larsen, Tone Rasmussen and Victor Labay Ong for excellent technical assistance. This study was supported with grants from the Norwegian Research Council.
- Received January 30, 2009.
- Revision received March 17, 2009.
- Accepted April 8, 2009.
- Copyright© 2009 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.