


Abstract
Background: Conventional chemotherapy is ineffective for high-grade chondrosarcomas, highlighting the need for improved chemotherapies. Various clinical trials have been initiated using antiapoptotic agents and perifosine, and are truly in the experimental phases. Chondrosarcoma is still therefore considered a surgical disease despite its aggressive features of recurring locally and spreading to the lungs. Bisphosphonates inhibit growth of various cell types, including cancer cells and perhaps chondrosarcoma. Materials and Methods: The effect of different concentrations of alendronate on cell proliferation, migration, apoptosis and cytoskeleton reorganization as well as on the regulation of intracellular protein expression were analyzed for the high-grade chondrosarcoma cell line CS-1. Mevalonate pathway intermediates were used in some experiments to assess mechanistic aspects. Results: Alendronate decreased cell viability of CS-1 by inhibiting cell proliferation and cell migration. Alendronate-induced loss of cell viability led to a sequence of events including apoptosis and cytoskeletal rearrangements. Moreover, changes in the expression levels of various proteins involved in cell proliferation, migration, cell cycle, apoptosis and cytoskeleton reorganization were demonstrated. Conclusion: Alendronate exerts antiproliferative effects by perturbing various signaling pathways in CS-1 cells. These findings may lead to new treatment options for high-grade chondrosarcoma.
Resection with an adequate margin is the only effective standard option for the treatment of chondrosarcoma. Although less than 10% of grade I chondrosarcomas relapse, the 5-year survival rate for high-grade tumors remains below 60% (1). Conventional chemotherapy regimens for high-grade chondrosarcoma generally have no therapeutic response, and novel approaches are desperately needed to improve the prognosis for these unfortunate patients.
Bisphosphonates (BPs) such as alendronate, a second-generation nitrogen-containing BP (N-BP), bind strongly to hydroxyapatite in bone and induce apoptosis in osteoclasts (2). These drugs have direct effects on other malignant tumors such as myeloma (3) or breast cancer (4-6). There is some evidence that N-BPs can inhibit the growth of chondrosarcoma cell lines (7, 8), but the intracellular mechanisms have not been investigated. N-BPs may inhibit the mevalonate pathway, resulting in apoptosis, cell cycle arrest or cytoskeletal rearrangements in cells of osteoclast lineage as well as cancer cells (9, 10).
It is hypothesized that alendronate might have some direct inhibitory effects on chondrosarcoma cell metabolism via the mevalonate pathway. The proposed inhibitory effect might be related to cell cycle arrest, increased apoptosis, unusual cytoskeletal rearrangements, or significant changes in intracellular protein phosphorylation or kinase activations. BP administration may therefore have clinical utility for the treatment of chondrosarcoma.
Materials and Methods
Cell line. The chondrosarcoma cell line (CS-1) was established from a surgically resected human high-grade chondrosarcoma removed from a 62 year-old male with metastatic chondrosarcoma (11). The patient had no previous exposure to either radiation or chemotherapy. Assessment of type II collagen mRNA and cytogenetic analysis were performed for characterization. The cells were routinely cultured in RPMI 1640 with 10% FBS in a humidified atmosphere of 5% CO2.
Cell proliferation and migration assay. Cell proliferation and migration assay were performed as described previously (12, 13). For cell proliferation, cells were seeded onto 96-well plates at a density of 1×104 mL and incubated for 24 hours. Indicated concentrations of alendronate or EDTA as a control were added and the cultures were subsequently incubated for 96 hours. Several intermediates of the mevalonate pathway, including trans,trans-farnesol (FOH, 30 μM), geranylgeraniol (GGOH, 40 μM) or mevalonolactone (MA, 1 mM) were added. Following the incubation, viable cells were detected using a spectrophotometric CellTiter 96 Aqueous cell proliferation assay according to the manufacturer's instructions (Promega, Madison, WI, USA). For the cell migration assay, CS-1 cells were treated with EDTA or alendronate (100 μM) for 48 hours. The chambers were constructed with Falcon multiwell cell culture plates and Falcon cell culture inserts (Becton Dickinson, Franklin Lakes, NJ, USA). Each upper chamber was pre-coated with 1% gelatin. Cells were suspended (1×105 /mL) and applied (300 μL) into each well in the upper chamber. The assembled chambers were incubated for the indicated periods (1-3 h) and then pictures of the invaded cells at the lower surface of the filters were taken. The cell number was determined by manual counting facilitated by microscopic visualization. All data were derived from at least 3 independent experiments.
Proteome analysis and validation. CS-1 cells were treated with either alendronate or EDTA (100 μM) for 48 hours. Proteome analysis was performed utilizing the Becton Dickinson power blot system. A total of 351 antibodies were used for the procedure. In brief, after isolation of total protein, 80 μg of protein were separated by SDS-PAGE and Western blotting was performed according to standard techniques. The membranes were then washed, dried and scanned.
Apoptosis assay. To assess apoptosis, terminal dUTP nick-end labeling assay was performed. Apoptosis was quantified using the dye-uptake bioassay (Biocolor Ltd, Newtonabbey, UK) which uses the membrane alteration during apoptosis. Briefly, 1×104 CS-1 cells were plated in triplicate in 96-well plates and were allowed to attach for 24 hours. Each set was treated with 100 μM EDTA, 10 μM and 100 μM alendronate. Hydrogen peroxide was used as a positive control. Apoptotic cell number was determined by manual counting under microscopic visualization.
Immunofluorescence detection of actin. CS-1 cells were seeded onto 35 mm culture dishes on sterile coverslips in triplicate and allowed to attach and grow for 24 hours. Cells were then incubated for 48 hours in the presence of either alendronate or EDTA (100 μM). The medium was then removed and the cells fixed with 4% paraformaldehyde. The fixation buffer was removed and cells were washed with phosphate-buffered saline. For visualization of filamentous actin, the cells were exposed to rhodamine phalloidin (Invitrogen, Carlbad, CA, USA). Cover slips were mounted on the dishes using a 50% solution of glycerol in phosphate-buffered saline. The cells were examined under an Olympus Fluoview FV 1000 confocal microscope (Tokyo, Japan) for actin realignment.
Statistical analysis. Student's t-test was used to compare the differences between groups. Results are given as mean±SD and results with p<0.05 were considered statistically significant.
Results
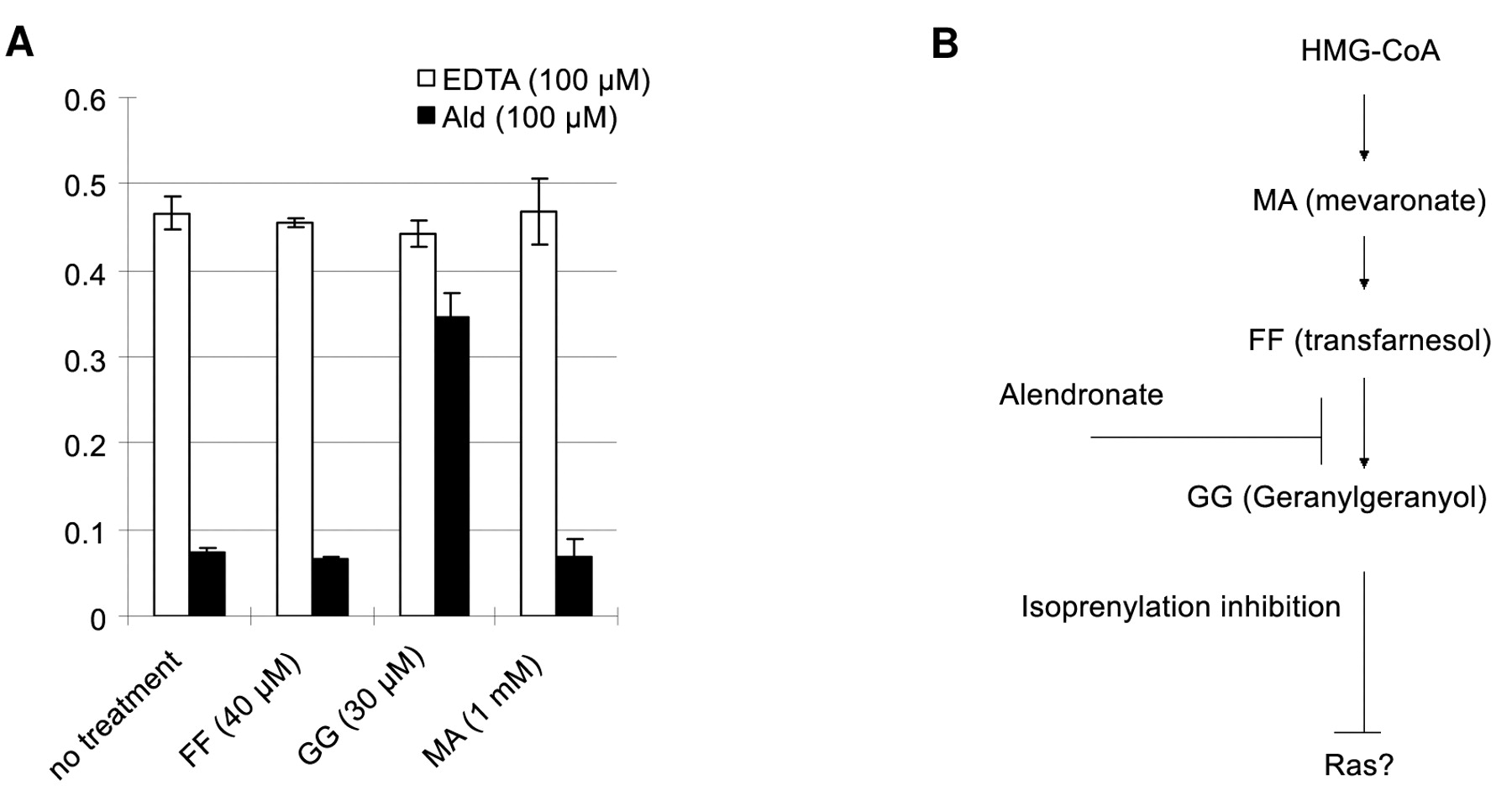
Alendronate inhibited the proliferation and migration of human chondrosarcoma cells in a dose-dependent manner (Figures 1, 2). The addition of GGOH prevented alendronate-induced loss of cell viability, but MA and FOH had no effect (Figure 3). The inhibitory effects of alendronate on chondrosarcoma cell proliferation through the mevalonate pathway were confirmed.
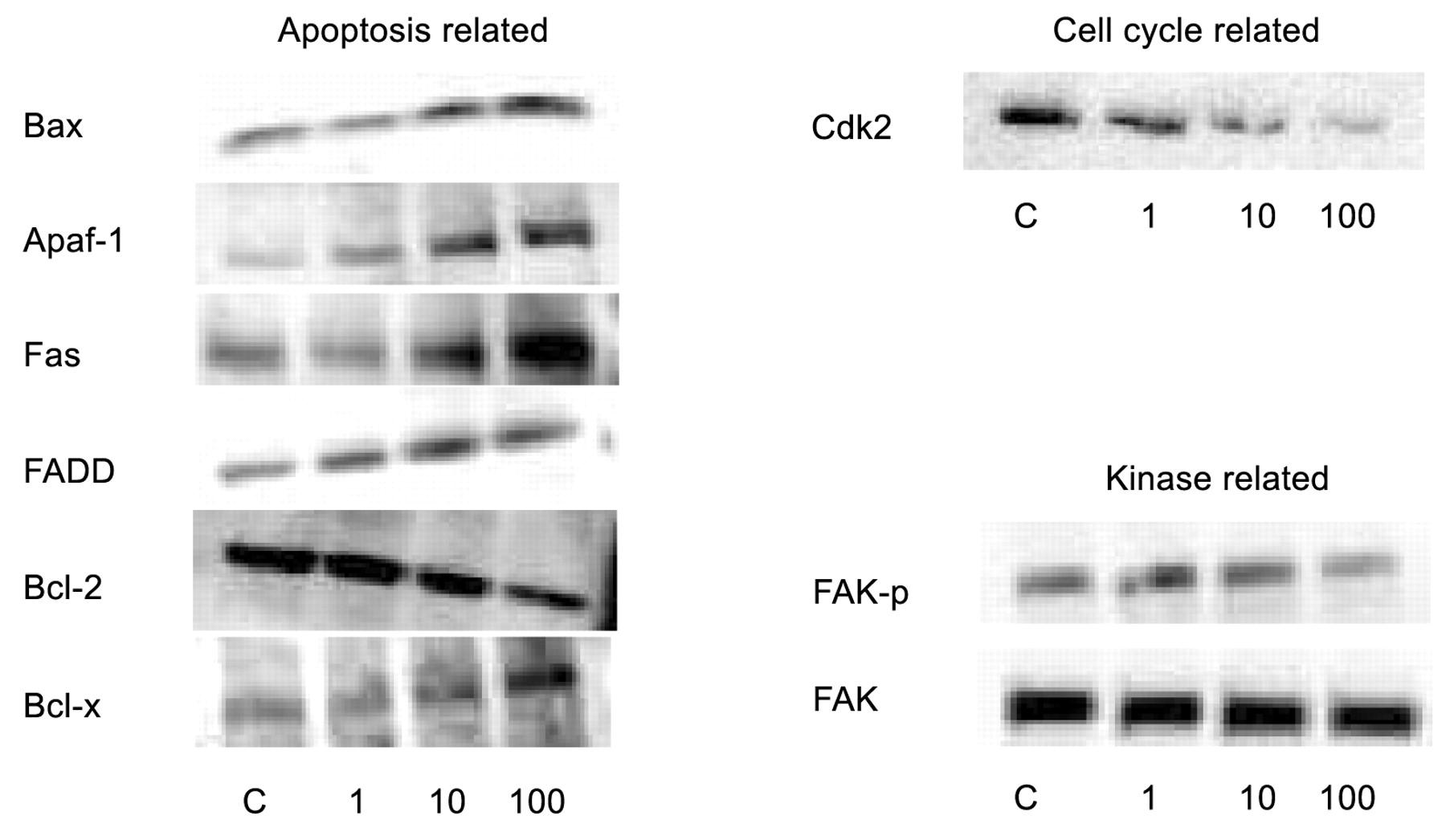
In order to elucidate the mechanism of alendronate on CS-1 cell growth and migration, high-throughput analysis of 351 proteins was performed using the power blot assay. In total, 31 proteins (8.8%), including KIP1/P27, Bax, Fas, FADD, Apaf-1 and hRad9 were up-regulated by over 2-fold. In contrast, a total of 27 proteins (7.7%), including bFGF, EGF receptor, RAS, GAP, cdk2, erbB and FAKphospho were down-regulated more than 2-fold (Table IA-F). These data suggest various pathways that alendronate acts upon, such as apoptosis, cell cycle regulation, cellular phosphorylation and cytoskeletal rearrangements. The regulation of a wide variety of proteins was confirmed by Western blot assay (Figure 4).
Proteome analysis suggested that one of the mechanisms in which alendronate exerted its action on CS-1 cells might be through apoptosis. Apoptosis was detected by the TUNEL assay and also quantified utilizing the phosphatidylserine flip of the CS-1 membrane. Microscopic examination revealed that apoptotic cells shrunk and were separated from the neighboring cells by a surrounding halo-like ring. Nuclear chromatin condensation was also observed (Figure 5). Quantification of these findings revealed alendronate induced apoptosis in a dose-dependent manner (Figure 6).
Proteome analysis was consistent with the morphological changes of CS-1. After incubation of CS-1 cells with alendronate for 48 hours, a reorganization of the cytoskeleton was observed under confocal microscopy (Figure 7).
Discussion
There are sporadic reports of possible adjuvant therapy using antiangiogenic chemicals for chondrosarcoma, but it is unclear at this time whether adjuvant therapy is a realistic proposition for high-grade chondrosarcoma.
In this study, a direct effect of alendronate on chondrosarcoma cells demonstrated that this drug may have clinical efficacy. Xenograft studies will be performed to confirm the in vitro results. This cell line in has been previously used in xenograft evaluation of drug efficacy.
One argument against alendronate therapy is that it is unlikely that an effective concentration of BP can be achieved at the site of tumor growth. BPs rapidly disappear from the circulation and are primarily taken up by bone due to the strong binding to exposed hydroxyapatite crystals. It is possible that the local concentration of BP that is capable of inducing apoptosis may be achieved at sites of active bone resorption because the drug is liberated by osteoclast action. Sato et al. concluded that local concentrations of BP released from hydroxyapatite into the resorption space would be substantially higher than concentrations in the circulation and could achieve a concentration of 1,000 μM (2). This concentration is high enough for inducing an antitumoral response against chondrosarcoma.

The effect of alendronate on the proliferation of CS-1 cells was analyzed. Microscopic analysis of cell proliferation after incubation with 100 μM EDTA (A) or alendronate (B 3 μM, C 10 μM, D 30 μM, E 100 μM, F 300 μM) for 24 hours. Alendronate inhibited the proliferation of human chondrosarcoma cells in a dose-dependent manner (G).

The motility of the cell is associated with its grade of malignancy. CS-1 cells were incubated with either EDTA (A-C) or alendronate (D-F) for 48 hours. Cells were suspended (1×105 cells /mL) and applied (300 μL) to a Boyden chamber system precoated with 1% gelatin. Images of the migrated cells were taken at 1 h (A, D), 2 h (B, E) and 3 h (C, F) and the cell number was determined. Cell migration was inhibited by treatment with alendronate (G).

Targeting of various proteins to the membrane is mediated by hydrophobic lipids such as farnesyl pyrophosphate and geranylgeranyl pyrophosphate produced via the mevalonate pathway. The addition of GGOH prevented alendronate induced loss of cell viability, but MA nor FOH were without effect (A). Thus, the results suggest that the target of alendronate is geranylgeraniol synthesis rather than farnesyl pyrophosphate synthase or isopentenyl pyrophosphate isomerase in the current study (B).

The results from high-throughput analysis were confirmed using by Western blot assay. The analysis was repeated three times.
The possible roles of protein prenylation by which alendronate might affect the function of cell types such as macrophages, osteoclasts, endothelial cells and cancer cells in vitro have been reported (14, 15). The question was raised whether the mechanism previously reported on various cell lines would be applicable to chondrsarcoma cells in vitro. Farnesyl pyrophosphate synthase and/or isopentenyl pyrophosphate isomerase are crucial intracellular targets of N-BP action (9, 15). Geranylgeraniol is also capable of partially preventing apoptosis and blocking caspase-3 like activity in J774 macrophages (15). Therefore, various intermediates of the mevalonate pathway, including FOH, GGOH and MA were tested in the cell proliferation system, and the data demonstrated that the target of alendronate was GGOH synthesis rather than farnesyl pyrophosphate synthase or isopentenyl pyrophosphate isomerase in the current study.
In order to elucidate mechanistic aspects pertaining to alendronate's effects on CS-1 cells, expression of 351 proteins were investigated using the power blot assay. Proteins related to apoptosis (A, B), cell cycle (C, D) and kinase pathways (E, F) that were up-regulated or down-regulated over 2-fold are listed.
The pathways downstream of small guanosine triphosphate-binding proteins such as Ras have not been elucidated. To investigate the mitogenic and survival pathways downstream of Ras and the mechanism by which N-BPs induce antitumor effects, high-throughput analysis of the expression of 351 proteins was performed. Because of the sporadic changes in the expression levels of proteins involved in apoptosis regulation, focus was initially on whether apoptosis actually is the mechanism by which alendronate exerts its effect.
There are two major intracellular cascades important for the induction of apoptosis, the death receptor pathway and the mitochondrial-related pathway. For the death receptor pathway, the up-regulation of Fas, the initiator of this signal transduction pathway which results in the induction of apoptosis, as well as FADD, the adapter protein of Fas, was confirmed, suggesting a plausible hypothesis for the action of alendronate on chondrosarcoma. Alendronate treatment increased the Fas/FADD interaction, which in turn allowed for the recruitment of the death-effector domain-containing initiator caspase-8, initiating apoptosis.
Regarding the second cascade, members of the Bcl-2 family play a critical role in regulating the response of cells to a wide variety of apoptotic signals. Bcl-X, Bax, Bad and Bak also regulate this pathway. Bax homodimers are thought to promote cell death, whereas heterodimers with Bcl-2 or Bcl-X appear to block cell death. Up-regulation of Bax and down-regulation of Bcl-2 were observed following alendronate treatment. These findings support the already confirmed results of protein-protein interactions that lead to apoptosis. Up-regulation of Smac/Diablo and Apaf 1, both key proteins downstream of cytochrome c which are released from mitochondria after being triggered by certain stimuli (in this case, alendronate), were also observed. These data suggest, but do not confirm, the close relationship between the antitumor effect of alendronate and apoptosis mechanisms. As judged by two different methods, the dye-uptake bioassay and TUNEL assay, apoptosis was involved in the antitumor effect produced by alendronate. Other in vitro studies have shown that N-BPs exert proapoptotic effects on a variety of human tumor cell lines (myeloma, breast cancer, prostate and pancreas) in a concentration- and time-dependent manner (2, 3, 5, 16). Although these studies were able to confirm the process of apoptosis by various methods, none of the above studies were able to elucidate the mechanisms by which BPs exert their proapoptotic effects.

The intracellular mechanism by which alendronate exerts its action on CS-1 cells appears to be apoptosis, based on the results of the proteome data. By microscopic examination, CS-1 cells were observed undergoing apoptosis. Apoptotic cells shrank and were separated from the neighboring cells by the surrounding halo-like ring. Nuclear chromatin condensation was also observed. A, EDTA 100 μM; B, alendronate 10 μM; C, alendronate 100 μM; D, H2O2.
In the process of apoptosis, there are characteristic morphological changes including loss of adhesion, actin-cytoskeletal structural reorganization into cortical rings, cell rounding and contracting, and generation of apoptotic bodies. After the initial phase of contraction and blebbing, caspase-mediated cleavage of actin monomers probably induces disassembly of actin filaments (17). Caspase-mediated proteolysis of structural and adhesion proteins reportedly plays an important role in the morphological changes that characterize apoptotic cell death (18). Tassone et al. showed that the pancreatic cancer cell line PANC-1 PC underwent significant actin architecture rearrangements after a 48-hour exposure to zoledronic acid (3). They concluded these modifications indicated N-BPs induced actin rearrangements into cortical rings and that these events drive cells into apoptosis. Thus it was important to determine if morphological and cytoskeletal changes were induced during apoptosis induction by alendronate in the CS-1 cells. After 48-hour treatment with alendronate, significant actin-cytoskeletal reorganization was observed in the chondrosarcoma cells as demonstrated by actin staining. These modifications indicate alendronate induces actin rearrangements and these events may lead the cells into the apoptotic process.

Induction of programmed cell death was detected using the TUNEL assay. After incubation with 100 μM alendronate (A), fragmented DNA was detected with this assay system. 100 μM EDTA was used as a control (B). Apoptosis was also confirmed by a dye-uptake bioassay that is based upon membrane alteration. After incubation with either 100 μM EDTA (C) or alendronate (D) for 48 hours, cell number was determined and apoptosis was detected in a concentration-dependent manner (E).
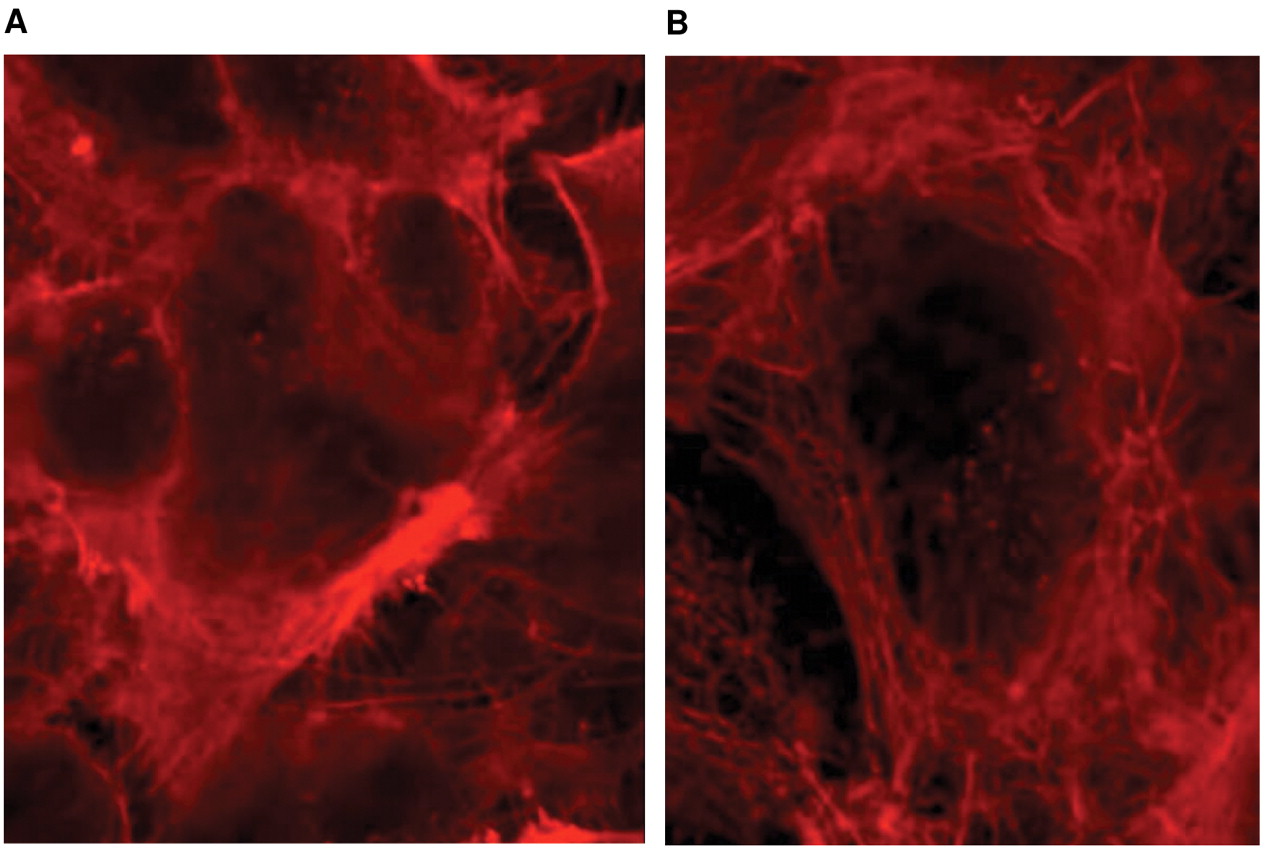
Proteome analysis suggested alterations in the cytoskeleton and associated morphological changes in the CS-1 cells. In fact, after incubation of CS-1 cells with alendronate for 48 hours, reorganization of the cytoskeleton was observed under confocal microscopy.
Finally, because there were considerable changes in the levels of proteins involved in the cell cycle, flow cytometric analysis of CS-1 cells following treatment with alendronate was performed. N-BP reportedly inhibits cell cycle progression and increases the proportion of cells arrested in the S-phase in rat and human osteosarcoma cells (19). In these experiments using human chondrosarcoma cells, the proportion of cells arrested in the S-phase was not increased (data not shown). This observation may be explained by the slow proliferation rate of chondrosarcoma cells as compared with osteosarcoma cells.
BPs are considered safe agents that are widely prescribed for patients with osteoporosis. There are sporadic reports that BPs exhibit antiangiogenic activities against various tumors. This suggests that in addition to its anticancer activities against chondrosarcoma that were examined in this report, BPs may be employed as an adjuvant therapy for conventional chemotherapy. A synergistic effect of BP with anticancer drugs has in fact been demonstrated in human breast cancer (5, 20) and osteosarcoma cell lines (21). An additional topic of current investigation is interaction of BPs with various signal transduction inhibitors. However, the precise mechanism of interaction between BPs and other agents warrants further investigation.
Acknowledgements
The authors wish to thank Miyuki Murayama and Takefumi Yamaguchi for their technical assistances.
- Received December 19, 2008.
- Revision received February 9, 2009.
- Accepted March 16, 2009.
- Copyright© 2009 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Anti-miRNA Oligonucleotide Therapy for Chondrosarcoma
- Development of Stemness in Cancer Cell Lines Resistant to the Anticancer Effects of Zoledronic Acid
- miR-181a Targets RGS16 to Promote Chondrosarcoma Growth, Angiogenesis, and Metastasis
- Lung Cancer Cell Line Sensitivity to Zoledronic Acid Is BAX-dependent
- Pro-inflammatory and anti-angiogenic effects of bisphosphonates on human cultured retinal pigment epithelial cells
- The Potential for Isocitrate Dehydrogenase Mutations to Produce 2-Hydroxyglutarate Depends on Allele Specificity and Subcellular Compartmentalization
- Inhibition of Heat-shock Protein 27 Expression Eliminates Drug Resistance of Osteosarcoma to Zoledronic Acid