


Abstract
The effects of sodium phenylacetate (NaPa), an antitumoral molecule, on cell death and matrix metalloproteinase (MMP) activities and synthesis were investigated in two metastatic breast tumour cell lines, MDA-MB-231 and MDA-MB-435, cultured on three-dimensional type I collagen gels (3-D cultures). In both cell lines, NaPa inhibited cell proliferation and induced apoptotic cell death as measured by TUNEL assay, with an IC30 of 20 mM and 10 mM for MDA-MB-231 and MDA-MB-435 cells, respectively. In MDA-MB-231 cells, NaPa also induced (i) an autophagic process evidenced by the appearance of autophagic vacuoles and an increased phosphatase acid activity, (ii) the formation of pseudopodia and (iii) an increase in MMP-1 and MMP-9 secretion without affecting MT1-MMP. In NaPa-treated MDA-MB-435 cells, no autophagic vacuoles were formed but F-actin depolymerisation was observed. MMP-1, MMP-9 and MT1-MMP levels were strongly enhanced in these cells but MMPs were not secreted and accumulated intracellularly. When breast cancer cells were treated with NaPa in the presence of an MMP inhibitor (GM6001), apoptotic cell death decreased and the induction of autophagic vacuoles in MDA-MB-231 cells was inhibited. Taken together, these data suggest that MMPs are involved in the autophagic cell death and/or apoptosis of breast tumour cells.
The term ‘apoptosis’ was originally described as the condensation of chromatin at the nuclear membrane, prominent condensation of cytoplasm and fragmentation of cell nucleus, while organelles were well preserved and autophagocytosis was absent (1). On the other hand, the cell suicide program has been found to involve the autophagic/lysosomal compartment (2).
Isoprenoid synthetic pathway inhibitors and farnesyl transferase inhibitors represent a new class of anticancer drugs, very promising in blocking tumour growth without inducing major toxicity. These antiproliferative molecules induce DNA fragmentation, cytochrome c release and caspase 3 activation preferentially in ras-transformed cells (3). Among these, sodium phenylacetate (NaPa), a product of phenylalanine metabolism, has been shown to induce cytostasis (4) and differentiation (5) in a variety of tumour models. Thus, it has been previously shown that NaPa treatment can induce tumour cell apoptosis alone (6-8) or in association with tamoxifen (9). In vivo studies on animal models demonstrated an antitumoral effect of NaPa and derivatives. Thus, MCF-7 ras tumour development in nude mice was blocked by NaPa treatment (6). This NaPa antitumoral activity was related not only to antiproliferative and proapoptotic effects on MCF-7 ras but also to stromal paracrine effects (10).
Invasion and metastasis of cancer cells are complex multistep processes that involve cell adhesion, proteolytic enzyme degradation of the extracellular matrix (ECM) and production of growth factors that influence cell migration (11). Several gene products are involved in these processes, among which matrix metalloproteinase (MMP) activities were shown to be increased during progression of breast cancer cells (12). The secretion of various MMPs in breast cancer cell lines of different tumorigenicities correlates with the biological behaviour of theses cells (13). The modulation of tumour cell adhesion to ECM proteins is also an important process in tumour invasion (14) and the regulation of both MMP activities and ECM remodeling has been shown to affect apoptosis in different systems (15).
In this study, the NaPa effect on the activities and/or synthesis of four major MMPs (MMP-1, MMP-2, MMP-9 and MT1-MMP) secreted by MDA-MB-231 and MDA-MB-435 cells grown in 3-D type I collagen gel cultures was investigated. MDA-MB231 and MDA-MB435 are oestrogen receptor (ER)-negative and hormone-independent mammary adenocarcinoma cell lines (16). MDA-MB-231 is an invasively growing cell line which expresses glandular and squamous cytokeratins. MDA-MB-435 is an invasive and highly-metastatic cell line which expresses only pancytokeratin markers and low level of MUC-1 as compared to MDA-MB-231. Whereas MDA-MB-231 cells xenografted in fat pads of nude mice give no metastasis, MDA-MB-435 cells proliferate and give rise to lung metastasis in almost all the mice (17). In parallel to measuring the activity and synthesis of secreted MMPs, this study investigated the NaPa effects on the proliferation, type I apoptosis and autophagic type II apoptosis of these two different tumour cell lines. Data that support the concept that the synthesis and/or secretion of MMPs could be responsible for the autophagic cell death and/or apoptosis of breast tumour cells are presented.
Materials and Methods
Reagents. All culture media and additives were purchased from Gibco Life Technologies (Invitrogen Corporation, Cergy-Pontoise, France). Sodium phenylacetate (NaPa) was from Seratec (Courville sur Eure, France), rat tail type I collagen (354236) from BD Biosciences (Le Pont de Claix, France) and collagenase H (1074) from Roche Diagnostics (Meylan, France).
Monoclonal mouse anti-MMP-1 (Ab-1, IM35), anti-MMP-9 (Ab-1, IM09) and anti-MT1-MMP (Ab-1, IM39) antibodies, and polyclonal rabbit anti-MMP-9 antibody (4444236) were from Calbiochem (VWR Intern., Strasbourg, France). Alexa 488-conjugated rabbit anti-mouse (A11059) and Alexa 488-conjugated goat anti-rabbit (A11037) secondary antibodies were from Molecular Probes (Invitrogen Corp.). HRP-conjugated goat anti-mouse secondary antibody (P0447) was from Dako (Trappes, France). FluoProbe 547-phalloidin (FP-AZ0330) was from FluoProbes (Interchim, Montluçon, France) and monodansylcadaverine (D4008) from Sigma (Saint Quentin Fallavier, France). All reagents used for electrophoresis were from Bio-Rad S.A. (Ivry sur Seine, France).
Cells. MDA-MB-231 and MDA-MB-435 cells were purchased from ATCC (Manassas, VA, USA). These two human breast carcinoma cell lines derived from two pleural effusions were routinely cultured in DMEM supplemented with 10% FCS (medium A). The absence of mycoplasms was routinely checked. For experiments, preparation of single cell suspensions was performed using 0.53 mM EDTA solution.
Three dimensional (3-D) type I collagen gels and 3-D cell culture. Gels were prepared according to the manufacturer's protocol (final type I collagen concentration: 2.4 mg/mL) and poured into 96-well plates to obtain approximately 2 mm thick gel layers. After gelling they were covered with serum-free medium B (DMEM-F12 medium with 0.2% BSA) for 3 hours and then used. Tumour cells to be tested were starved in medium B for 4 hours prior to the assay. At this time, cells (4×104/well) were seeded onto the top of the 3-D gels in medium B and cultured overnight. Culture media were then removed and NaPa (0 to 20 mM, 100 μL/well) was added in fresh medium B for 48 hours. At this time conditioned media were collected and their related cells lysed for further zymography and Western blot analysis. All culture conditions were performed in triplicate.
Zymography. MMP-1, MMP-2 and MMP-9 activities were analysed by zymography according to the method of Levkau et al. (18). Equal amounts of protein in the conditioned media were resolved under non reducing conditions on 10% polyacrylamide gels co-polymerized with 0.5 mg/mL type I collagen for MMP-1 activity or on 7.5% polyacrylamide gels co-polymerized with 1 mg/mL gelatin for gelatinase activities. After zymogram staining with Coomassie Blue G-250, lysis activities were quantified by densitometric scanning using an Image J processing program. Zymography was also performed in the presence of 15 mM EDTA to confirm metalloproteinase activity.
Western blots. After conditioned media were collected, cells were released from the top of the 3-D gels by complete collagenase digestion of the gels following the collagen manufacturer's protocol, then they were lysed in ice-cold lysis buffer as described previously (19). Standardized aliquots of conditioned media and of their related total cell lysates (20 μg protein per lane) were run on a 7.5% SDS-polyacrylamide gel. After protein transfer onto nitrocellulose membranes (0.2 μm porosity), membranes were incubated in TBS (0.05M Tris pH 7.4, 0.1 M NaCl) containing 0.05% Tween-20 and 5% non fat dry milk for 1 hour and then probed in the same buffer with the primary antibody, either anti-MMP-1 (1/75), anti-MMP-9 (Ab-1, 2 μg/mL) or anti-MT1-MMP (10 μg/mL), overnight at room temperature. After washes and 1 hour incubation with HRP-conjugated secondary antibody (1/1000) in complete TBS buffer, proteins were visualized using the chemiluminescence detection method (ECL plus Western Blotting Detection System from Amersham, Saclay, France). MMP levels were quantified by densitometric scanning using the Image J processing program.
Fluorescence microscopy. MDA-MB-231 and MDA-MB-435 cells prepared in medium B as described above were plated on the top of 3-D gels (105 cells/cm2 well) in Lab-Tek TC chamber slides (Nunc, Fisher Bioblock Scientific, Illkirch, France) and treated with 20 mM and 10 mM NaPa respectively in medium B for 48 hours. At the end of the incubation, the cells were washed in PBS and fixed on the slide with 4% paraformaldehyde (PFA) in PBS for 10 minutes at room temperature.
For actin and MMP staining, the fixed cells were successively permeabilized with 0.1% Triton® X-100 in PBS for 5 minutes, washed and incubated with 1% BSA for 30 minutes to reduce non-specific background staining. Specific staining was then run and after three washes and gasket removal, cultures were mounted with Vectashield mounting medium with DAPI to counterstain nuclei (Vectashield H-1200; Vector Laboratories). Immunostaining was observed under a DMRB fluorescence microscope (Leica Microsystèmes). Actin filaments (F-actin) were stained with FluoProbe 547-phalloidin (1/40 in PBS for 20 minutes). For MMP staining, slides were incubated with an anti-MMP-1 (1/125) or anti-MMP-9 (polyclonal, 1/150) primary antibody in PBS overnight at 4°C. After three washes with PBS, slides were incubated with Alexa 488-conjugated rabbit anti-mouse (1/800) or goat anti-rabbit (1/800) secondary antibodies respectively for 1 hour.
TUNEL labeling assay was performed to detect apoptotic cells through their fragmented DNA according to the manufacturer's instructions (MEBSTAIN Apoptosis kit Direct, IM3171; Immunotech, Beckman-Coulter, Roissy, France).
Labeling of autophagic vacuoles with monodansylcadaverine (MDC) was performed according to Biederbick et al. (20). Cells were cultured and treated with NaPa for 48 hours in Lab-Tek TC chamber slides as described above. At the end of the assay, cells were incubated with 0.05 mM MDC in free DMEM/F-12 medium for 20 minutes in the dark. After 3 washes in PBS, cells were fixed with 4% PFA. The slides were immediately mounted with Vectashield mounting medium with or without DAPI (H-1000) and observed under a fluorescence microscope (excitation filter: 340-380 nm, barrier filter: 425 nm). In some experiments, the MMP inhibitor GM6001 (364205, Calbiochem) was added (25 μM) together with NaPa and controls received equivalent dilution of DMSO vehicle alone.
Results
Antiproliferative and proapoptotic effects of NaPa on MDA-MB-231 and MDA-MB-435 cells in 3-D culture. Initially, in 3-D culture conditions, a dose-dependent inhibition of MDA-MB-231 and MDA-MB-435 cell proliferation by NaPa was shown. After 48 hours of treatment, the IC30s were approximately 20 mM and 10 mM NaPa for MDA-MB-231 and MDA-MB-435 cells, respectively. Therefore, all following assays were run using these concentrations, except where otherwise stated.
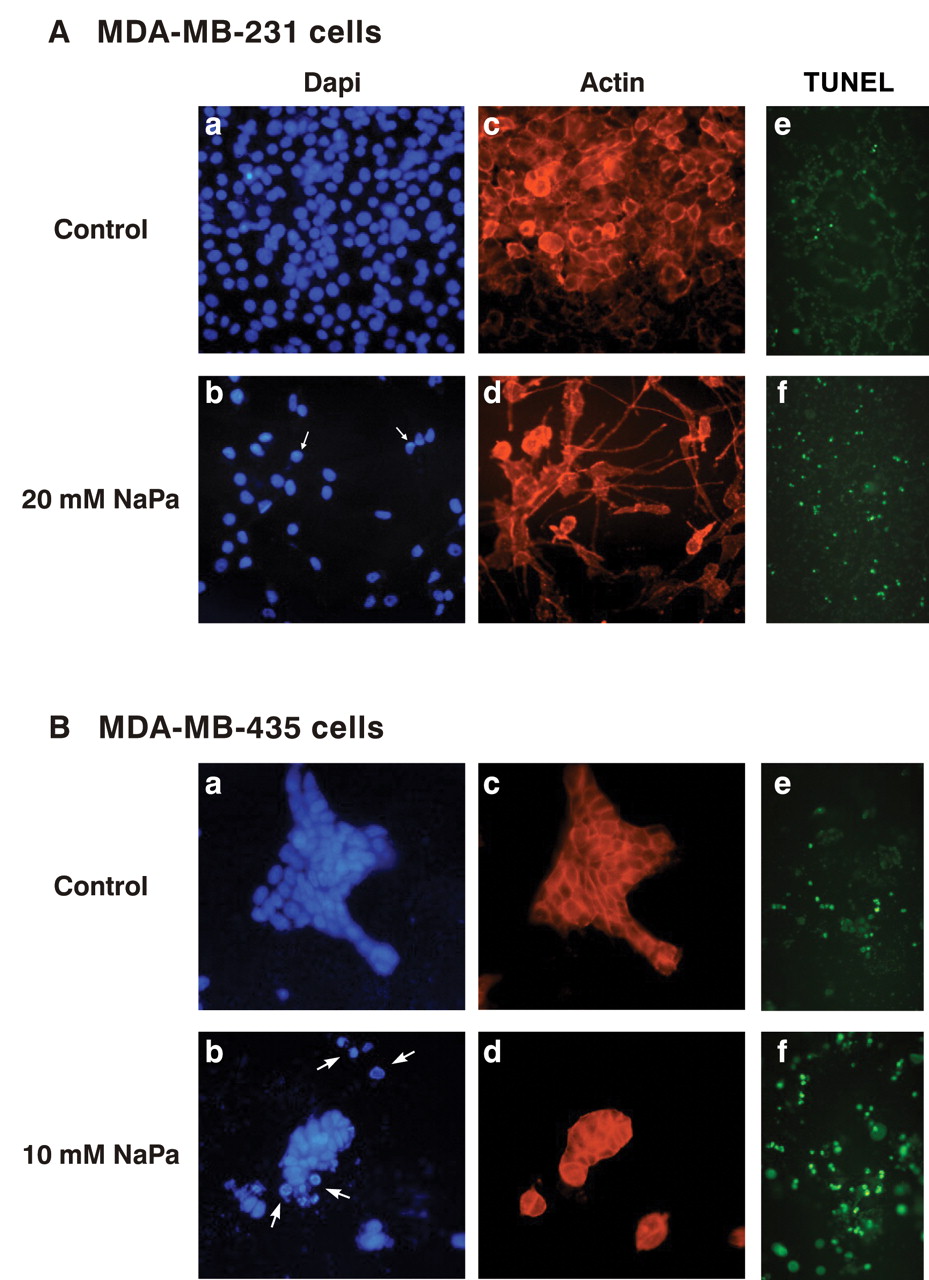
As changes in tumour cell morphology were observed upon NaPa addition, further examination of the effect of NaPa on cell cytoskeleton using actin network staining was performed. Nucleus staining with DAPI was concomitantly performed (Figure 1A, B) to assess cell density and nuclear morphology and confirmed that NaPa induced a loss in cell number and evidenced the presence of numerous apoptotic nuclei in both NaPa-treated cell lines. Treated MDA-MB-231 cells mainly exhibited nuclei with a condensed heterogeneous chromatin (Figure 1A a, b) whereas apoptotic bodies were also frequently observed in treated MDA-MB-435 cells (Figure 1B a, b).
Cellular actin staining confirmed cell cytoskeleton modifications under NaPa treatment. Thus, MDA-MB-231 cells developed a thick and very long pseudopod and F-actin was observed in the cortical area as in control cells (Figure 1A c, d). MDA-MB-435 cells grown under control conditions formed large islets of pavimentous cells with a cortical distribution of F-actin. Under NaPa treatment, islets broke up giving rise to smaller islets and numerous isolated cells which were only detected under the DAPI illumination as they lost their cortical F-actin (Figure 1B c, d). Moreover it was observed that all cells negative for F-actin exhibited morphologies of apoptotic bodies.
Apoptosis, first assessed by cell nuclei condensation and the presence of apoptotic bodies, was confirmed by showing DNA fragmentation using a TUNEL assay (Figure 1A e, f and 1B e, f). Estimate of the percentage of apoptotic MDA-MB-231 cells in control cultures was less than 4% and reached approximately 10% of cells after 48 hours of 20 mM NaPa treatment. In control cultures, TUNEL-positive MDA-MB-435 cells were observed (approximately 10% cells) and were located at the edge of the islets. Most of the 10 mM NaPa-treated MDA-MB-435 cells became TUNEL positive (60 to 80% of cells) and this result was consistent with the observation of many apoptotic bodies.
Taken together these results showed that NaPa induced type I apoptosis associated with morphological changes in the two breast adenocarcinoma cell lines.
Concomitant autophagic cell death (type II apoptosis) induced in MDA-MB-231 cells by NaPa treatment. The presence of autophagic vacuoles characterizes autophagic cells. Monodansylcadaverine (MDC), an autofluorescent compound, selectively accumulates in autophagic vacuoles which appear as distinct dot-like structures under fluorescence microscopy. Whereas few MDC-positive cells (less than 2%) were present in control MDA-MB-231 cell cultures, 25 to 30% of these cells accumulated MDC after NaPa treatment (Figure 2A a, b). MDC staining was observed in isolated cells as well as in cell clusters. In MDC-positive cells, large fluorescent spots were present in the perinuclear region as well as all over the cytoplasm as described in CHO cells by Munafo et al. (21). In contrast, control and treated MDA-MB-435 cells exhibited very few or no MDC-positive cells (Figure 2B a, b).
Because it was demonstrated that MDC-labeled structures contained the lysosomal acid phosphatase enzyme (22), its activity in control and NaPa-treated cells was measured. In MDA-MB-231 cells, acid phosphatase activity was stimulated about 2-fold by a 48 hour NaPa treatment whereas in MDA-MB-435 cells NaPa strongly inhibited this activity (data not shown), likely by inducing complete apoptosis of these cells. These results are consistent with those based on MDC incorporation. Further investigation of a possible involvement of MMPs in the NaPa induced type I and/or type II cell death was performed. For this purpose, MDA-MB-231 and MDA-MB-435 cells were concomitantly treated with NaPa and the broad-spectrum MMP inhibitor, GM6001, for 48 hours. The double (NaPa+GM6001)-treated MDA-MB-231 cells exhibited nearly normal proliferation and only a few condensed nuclei were observed as in the control culture (data not shown). Moreover, GM6001 reduced the autophagic death process induced by NaPa as only few cells (8 to 11%) exhibited autophagic vacuoles in their cytoplasm (Figure 2A c). As seen in Figure 2B c, GM6001 also prevented disruption of MDA-MB-435 cell islets into isolated cells. These results showed that MMPs could play a role in the cell death processes induced by NaPa in MDA-MB-231 and MDA-MB-435 cells.

Antiproliferative and proapoptotic effects of NaPa on MDA-MB-231 (A) and MDA-MB-435 (B) cells in 3-D culture. MDA-MB-231 and MDA-MB-435 cells (4×104) were cultured onto the top of 3-D type I collagen gels in serum-free medium in the absence (control cells in a, c, e) or in the presence of 20 mM and 10 mM NaPa respectively (treated cells in b, d, f) for 48 hours. Nuclei were counterstained with DAPI: a,b. Arrows and arrowheads point to condensed nuclei and apoptotic bodies respectively. c,d: the same fields observed under TRITC-illumination.e,f: cell apoptosis was also assessed by using TUNEL labeling assay. TUNEL-positive cells appear as bright green cells.

Autophagic cell death (type II apoptosis) induced by NaPa treatment. MDA-MB-231 and MDA-MB-435 cells were cultured as described in Figure 1. After 48 hours of NaPa-treatment, labeling of autophagic vacuoles with monodansylcadaverine (MDC) was performed as described in the Materials and Methods. After fixation with 4% PFA, cells were immediately analyzed by fluorescence microscopy (excitation filter: 340-380 nm, barrier filter: 425 nm). a: Control cells; b: NaPa-treated cells; c: in some experiments, the MMPs inhibitor GM6001 (364205, Calbiochem) was added (25 μM) together with NaPa, MDC-labeling was then run as indicated above.
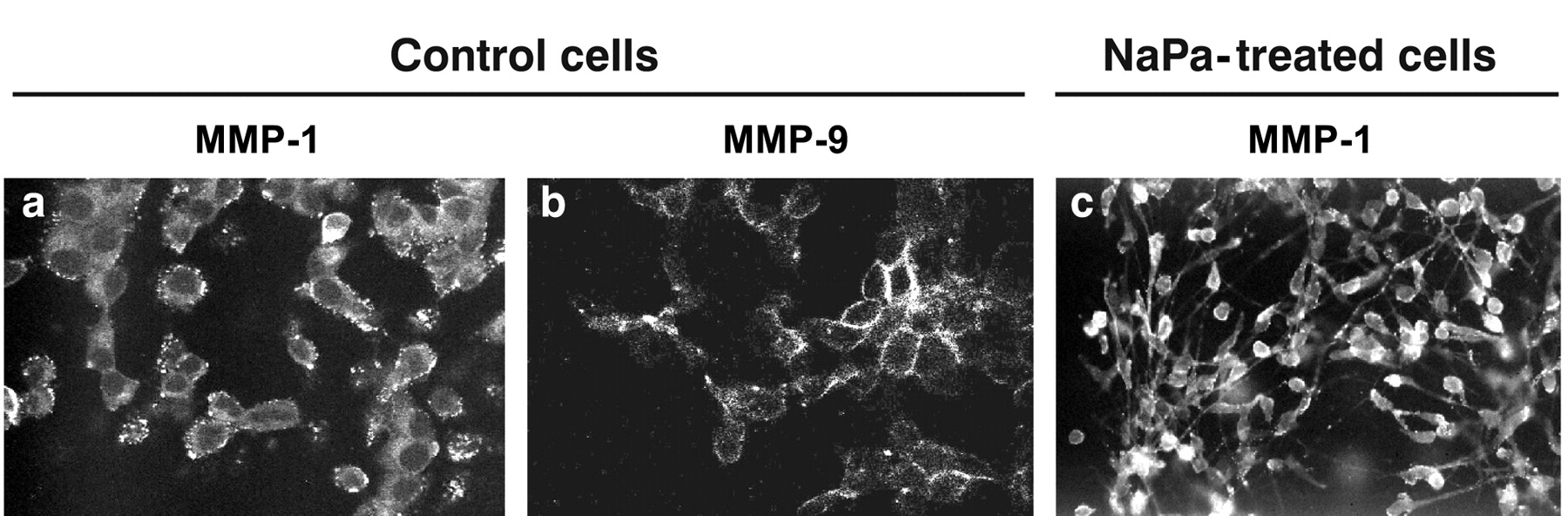
NaPa effects on MMP-1, MMP-2 and MMP-9 expression in MDA-MB-231 and MDA-MB-435 cells. Immunocytochemistry was initially used to evaluate the MMP expression in these two cell lines when cultured on 3-D type I collagen gels and showed the presence of MMP-1 and MMP-9 in control MDA-MB-231 cells (Figure 3). MMP-1 staining appeared as small dot-like structures uniformly distributed in the cytoplasm (Figure 3a) whereas that of MMP-9 mostly exhibited a cortical distribution (Figure 3b). NaPa treatment induced the large recruitment of these MMPs in the pseudopod which was concomitantly induced by NaPa, as shown for MMP-1 (Figure 3c). The presence of both MMP-1 and MMP-9 was also observed all over the cytoplasm of control MDA-MB-435 cells and no noticeable staining rearrangement was evidenced after NaPa treatment (data not shown). The presence of MMP-2 in the cytoplasm of both control or treated cell lines was not observed.
As it was observed by immunocytochemistry that both cell lines constitutively expressed MMP-1 and MMP-9 in the 3-D culture system, MMP-1 and MMP-9 expression patterns were further evaluated in control and NaPa-treated cells using zymography and Western blot analysis. After a 48 hour 3-D culture in the presence of increasing concentrations of NaPa (0 to 20 mM), the activity of MMPs secreted by MDA-MB-231 and MDA-MB-435 cells in their conditioned media were analysed by zymography. Results presented in Figure 4A show that increasing concentrations of NaPa enhanced the amounts of MMP-1 and MMP-9 secreted by MDA-MB-231 cells (lanes 1 to 4). By densitometry scanning these stimulations were estimated to be 3-fold for MMP-1 and 2.4-fold for MMP-9 in the presence of 20 mM NaPa (lane 4 vs. 1). The same NaPa concentrations did not induce the secretion of MMP-1 or MMP-9 by MDA-MB-435 cells nor that of MMP-2 by the two cell lines (lanes 5 to 8).

MMP-1 and MMP-9 expression in MDA-MB-231 cells. MDA-MB-231 cells were cultured as described in Figure 1. After fixation in 4% PFA and permeation with 0.1% Triton® X-100, cells were incubated with an anti-MMP-1 (1/125) or anti-MMP-9 (polyclonal, 1/150) primary antibody followed by Alexa 488-conjugated rabbit anti-mouse (1/800) or 488-conjugated goat anti-rabbit (1/800) secondary antibody, respectively, as described in the Materials and Methods. Cells were observed using fluorescence microscopy. a, b: Control cells, MMP-1 and MMP-9 expression respectively; c: MMP-1 recruitment in the pseudopod which was induced in NaPa-treated cells.
Western blot analyses were further performed on the same conditioned media and on their related cell lysates to confirm MMP-1 and MMP-9 expressions evidenced by immunocytochemistry and zymography data. Results are shown in Figure 4B. Western blot analysis of MMP-1 protein in control MDA-MB-231 cell lysates (lane 2) showed the presence of two bands, one of about 55 kDa and a main band of about 110 kDa which could correspond to a dimer of MMP-1. Only the 55 kDa was detected in the corresponding control conditioned medium (lane 1). Under NaPa treatment (20 mM), the level of this secreted form was enhanced by about 2.6-fold (lane 3 vs. 1), similar to the enhancement of MMP-1 activity measured by zymography (Figure 4A). A concomitant decrease (about 2.2-fold) of the intracellular amount of MMP-1 110 kDa form was observed (lane 4) whereas the intracellular 55 kDa form did not seem to vary. The presence of MMP-1 was not detected in conditioned medium of control (lane 5) or treated (lane 7) MDA-MB-435 cells, in agreement with the zymography data (Figure 4A). However, two forms of MMP-1, one of about 55 kDa and a main form of about 110 kDa, were present in control lysates (lane 6) as observed in control MDA-MB-231 cells. NaPa did not induce any detectable change in this intracellular MMP-1 distribution (lane 8 vs. 6).
In the 3-D assay, MMP-9 protein was not detected in the conditioned media of control (lane 1) or 20 mM NaPa-treated (lane 3) MDA-MB-231 cells although the presence of MMP-9 in the two conditioned media was evidenced by zymography (Figure 4A). MMP-9 protein was also not evidenced in the conditioned media of control or 10 mM NaPa-treated MDA-MB-435 cells (lanes 5 and 7). Nevertheless, NaPa induced changes in MMP-9 cytoplasmic levels, as attested by a decrease of the MMP-9 content in MDA-MB-231 cells (about 2.6-fold, lane 4 vs. 2) versus an increase in MDA-MB-435 cells (lane 8 vs. 6).
Because membrane-bound MT1-MMP was shown to be involved in MMP activation, the MT1-MMP level in both cell lysates was also measured. NaPa did not induce a noticeable change in the MT1-MMP content of MDA-MB-231 cells (lane 4 vs. 2) whereas it significantly increased it in MDA-MB-435 cells (lane 8 vs. 6).
These results show that NaPa enhances MMPs expression, i.e. MMP-1 in MDA-MB-231 and MMP-9 and MT1-MMP in MDA-MB-435, suggesting that MMP activities might be contributing to the NaPa-induced cell death that was suppressed in the presence of an MMP inhibitor.
Discussion
In the present study, the effects of sodium phenylacetate (NaPa), an antiproliferative and proapoptotic molecule (5, 23) on two human breast adenocarcinoma cell lines (MDA-MB-231 and MDA-MB-435) were investigated. For this purpose, 3-D cultures on type I collagen gels that are more relevant to the in vivo situation than 2-D cultures were used. In 3-D culture conditions, a 48 hour NaPa treatment induced an antiproliferative effect in both the MDA-MB-231 and MDA-MB-435 cell cultures. However, the inhibition of growth observed with the MDA-MB-231 cell line (IC30 of 20 mM) was less important than that previously observed when the assay was performed in 2-D cultures (IC30 of 5 mM) (8). Thus, 3-D cultures on type I collagen gels were able to protect MDA-MB-231 cells from apoptotic death induced by NaPa as has been observed by others in a different model (24). However, this survival effect did not seem to affect the MDA-MB-435 phenotype.
NaPa treatment also induced cell shape modifications of the surviving cells. Thus, MDA-MB-231 cells developed a large pseudopod without modification of the F-actin, which remained in a cortical position. In contrast, MDA-MB-435 cells, which grow as islets of pavimentous cells under control conditions, lost their cell-cell contacts together with their cortical F-actin when they were isolated from the islets. These results are in agreement with previous studies showing a morphological differentiating effect of NaPa and derivatives (23).
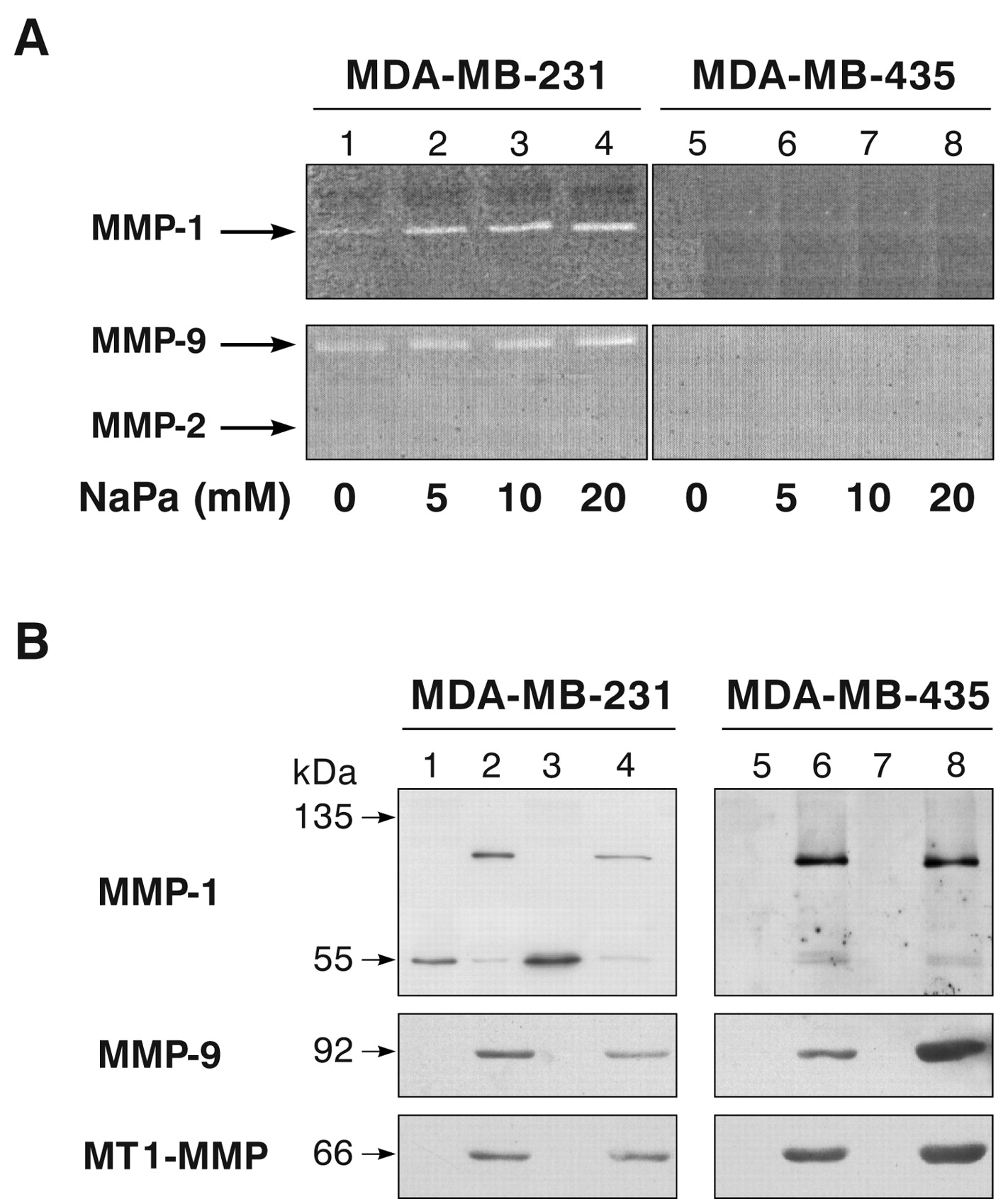
NaPa effects on MMP expression in MDA-MB-231 and MDA-MB-435 cells. (A) After 48-hour 3-D culture in the presence of increasing concentrations of NaPa (0 to 20 mM), the activity of MMP-1, MMP-2, MMP-9 secreted by MDA-MB-231 and MDA-MB-435 cells in their conditioned media were analysed by zymography as described in the Materials and Methods. (B) Western blot analysis was performed on the same conditioned media and their related cell lysates as described in the Materials and Methods. Immunoblots were run using either anti-MMP-1 (1/75), anti-MMP-9 (Ab-1, 2 μg/mL) or anti-MT1-MMP (10 μg/mL) as primary antibodies and HRP-conjugated secondary antibody (1/1000). Proteins were visualized using the chemiluminescence detection method. Lanes 1, 5: control conditioned medium; lanes 3, 7: conditioned medium of NaPa-treated cells (20 mM for MDA-MB-231 cells and 10 mM for MDA-MB-435 cells); lanes 2, 6: corresponding control cell lysates; lanes 4, 8: corresponding treated cell lysates.
Previous studies evidenced that programmed cell death (PCD) comprises at least (i) apoptosis, or type I PCD, characterized by a chromatin margination and condensation followed by nucleus and cell fragmentation and (ii) autophagic cell death, or type II PCD, which exhibits an extensive organite degradation in the cytoplasm preceding nuclear destruction. These PCD also differ in cytoskeleton fates. Early stages of apoptosis are associated with F-actin depolymerization and cytokeratin degradation whereas an intact cytoskeleton is necessary for autophagic cell death (25).The presented results are in agreement with these findings as NaPa-treated MDA-MB-435 cells lost their cortical F-actin and could only undergo type I apoptosis whereas NaPa-treated MDA-MB-231 cells, in which the actin cytoskeleton appeared well preserved, could undergo an apoptotic death as well as an autophagic process, the latter being much more prominent. The results also demonstrated (i) that the two cell lines respond differently to NaPa treatment though both are wild type for the tumour suppressor PTEN (26) and (ii) that apoptosis and autophagy can arise at the same time in NaPa-treated MDA-MB-231 cell cultures, in agreement with previous studies showing that apoptosis and autophagy may occur in the same tissue as well as in the same cell as reported in mammary gland epithelial cells (27).
Numerous studies have suggested that MMPs are involved in the metastatic progression of human cancer (28) but the abundance of the latent proMMPs in normal tissues and fluids suggest a complex regulation in tumours. NaPa treatment was first shown to decrease MDA-MB-231 cell invasiveness through Matrigel (29). Here it is shown that the levels of MMP-1 and MMP-9 activities present in the conditioned medium of NaPa-treated MDA-MB-231 cells were nevertheless highly increased and that NaPa enhanced the secretion of these two MMP proteins from their cytoplasmic stocks. In MDA-MB-435 cells, NaPa increased the level of MMP-9 alone which, in contrast, was not secreted and remained cytoplasmic. This absence of secretion could be due to the presence of collagen I in a 3-D polymerized form as it was evidenced that NaPa induced the secretion of MMP-1 and MMP-9 in conditioned medium of MDA-MB-435 cells in 2-D culture system (data not shown). This is consistent with previously reported data showing that 3-D collagen I gels specifically modulate the MMP expression in various cells (19, 30).
NaPa treatment induced a morphological change of MDA-MB-231 cells which adopted a spindle shape. It cannot be excluded that this reorganization of the cytoskeleton as well as modifications of cell adhesion molecules could be responsible for the increase of MMP-1 and MMP-9 since it was reported that paclitaxel treatment of breast cancer cells induce a change in cytoskeleton organization associated with a concomitant increase of MMP-9 secretion (31). On the other hand, the NaPa antitumoral activity is related not only to antiproliferative and proapoptotic effects on breast tumor cells but also to stromal paracrine effects. Thus, it has been shown that NaPa treatment modulated the synthesis and secretion of autocrine and paracrine growth factors secreted by MCF-7 ras cells (10) and in this context, it cannot be excluded that these factors could also be involved in MMP modulation.
It is well known that MMPs play an important role in the tissue remodeling of healthy tissues and diseases by degrading stroma proteins and ECM components as well as by interacting with cell surface receptors (28, 32). By these ways, they may promote cell migration as well as apoptosis. Thus, study of MMPs and their inhibitors in breast cancer evidenced high levels of MMP-1 and MMP-9 in tumours associated with a high rate of distant metastases (33) and there is increasing in vitro evidence for the involvement of MMP-9 in the apoptosis phenomena (34). The presented results suggest a direct relationship between NaPa-induced MMP activities and PCD. Thus, inhibition of MMP activity by GM6001, a broad-spectrum MMP inhibitor, prevented the production of autophagic vesicules induced by NaPa treatment in MDA-MB-231 cells, and nuclei integrity was mostly preserved in MDA-MB-231 and MDA-MB-435 showing an involvement of MMPs in these processes. These results are in agreement with previous reports showing that MMP-2 and MMP-9 activities increased in regressing tumours, coinciding with the peak of apoptosis (35), and that other MMP inhibitors, such as BB-94, protect cells against apoptosis (35, 36).
In conclusion, the autophagic cell death of certain metastatic breast tumour cells could be induced indirectly by/through MMP activation/synthesis and secretion. This would challenge the principle of using MMP inhibitors in cancer treatment and requires further in vitro and in vivo experimental studies.
Acknowledgements
This work was supported by a grant of the Institut National du Cancer, France.
- Received September 30, 2009.
- Revision received December 17, 2009.
- Accepted January 19, 2009.
- Copyright© 2009 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved





{kind=link}
{kind=link}
{kind=link}
{kind=link}