


Abstract
Background: We have previously screened many natural products for their cytotoxic effects on neuroblastoma cell lines, and two diarylheptanoids derived from Alpinia officinarum, 7-(4″-hydroxy-3″-methoxyphenyl)-1-phenyl-4E-hepten-3-one (Compound 1) and (5R)-5-methoxy-7-(4″-hydroxy-3″-methoxyphenyl)-1-phenyl-3-heptanone (Compound 2), were shown to have potent cytotoxicity. On this basis, a detailed study of the antitumor activities of these two diarylheptanoids in neuroblastoma cell lines was performed. Materials and methods: Cytotoxicity was assessed by 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide (MTT) assay. To measure apoptosis, nuclear shrinkage was monitored by Hoechst 33342 staining, and activation of caspases 3 and 9 was monitored by Western blotting. Cell cycle status was evaluated by flow cytometry with propidium iodide staining. Differentiated cells were photographed and counted in a randomized fashion. Results: Both diarylheptanoids showed significant cytotoxicity against neuroblastoma cell lines (IMR-32, SK-N-SH, NB-39). The compounds induced nuclear shrinkage and fragmentation, and activated caspase-3 and caspase-9. Flow cytometric analysis revealed induction of S-phase cell cycle arrest concurrently with an increased sub-G1 cell population. Moreover, a low concentration (10−8 M) of Compound 1 induced significant neurite branching in the NB-39 cell line. Conclusion: Diarylheptanoids derived from A. officinarum have marked activity against neuroblastoma cells, acting through multiple mechanisms. Our results suggest that the two compounds studied here may be useful for the treatment of patients with neuroblastoma.
Neuroblastoma is the most common solid tumor in children, and is a major cause of death from neoplasia in childhood. Most neuroblastomas are at an advanced stage at diagnosis, for example in the analysis of the Children's Oncology Group, 45% of patients with this disease had tumors of stage 4 (1). Neuroblastoma shows a wide range of clinical features. The five-year survival rate for patients with malignant neuroblastoma is less than 40%, despite intensive chemoradiotherapy and surgery (2). On the other hand, some neuroblastoma patients experience good survival without intensive therapy because of spontaneous regression of the tumor. In particular, stage 4S, a special stage for neuroblastoma, shows good prognosis despite the occurrence of distant metastasis to bone marrow, liver and/or skin, which appears to be involved in the process of spontaneous regression. Although the mechanisms of spontaneous regression are still unclear, some reports have suggested that apoptosis and/or differentiation might be involved (3, 4). In fact, treatment of 13-cis-retinoic acid, a differentiation inducing-compound, improved outcome among neuroblastoma patient without progressive disease (2).
Many clinically used anticancer drugs have been developed from natural plants, and thus we have screened many natural compounds to find cytotoxic agents against human neuroblastoma cell lines (5, 6). In our recent study, diarylhepta-noids derived from Alpinia officinarum, a plant used for gastric complaints and as an analgesic and anti-emetic in traditional oriental medicine, were revealed to have cytotoxic activity against neuroblastoma IMR-32 cells (7). In particular, 7-(4″-hydroxy-3″-methoxyphenyl)-1-phenyl-4E-hepten-3-one (Cpd 1) and (5R)-5-methoxy-7-(4″-hydroxy-3″-methoxyphenyl)-1-phenyl-3-heptanone (Cpd 2) showed potent cytotoxicity, with 50% effective concentration (EC50) values of the order of 10−7 M (7). There are also reports describing the anticancer activity of other diarylheptanoids, such as curcumin, a representative diarylheptanoid derived from turmeric (Curcuma longa), which induces apoptosis and neurite outgrowth in neuroblastoma cells (8-10). It has been reported that Cpd 1 inhibits proinflammatory mediators such as interleukin 1β and tumor necrosis factor-α induced by lipopolysaccharide (11), and we have previously reported that Cpd 1 and Cpd 2 inhibit 12-O-tetradecanoylphorbol-13-acetate (TPA)-induced inflammation (12). However, there are no reports pertaining to apoptosis-inducing and/or differentiation-inducing activity of either of these compounds.

Cytotoxic effects of diarylheptanoids on neuroblastoma (IMR-32, SK-N-SH and NB-39) cell lines as shown by the MTT assay. A, The chemical structures of the two diarylheptanoids used in the present study. B, Cells were exposed to Cpd 1 and Cpd 2 for 48 h. Each plot shows the mean±S.E.M. survival rate relative to the control group.
To clarify their cytotoxic mechanisms and to evaluate their potential antitumor activity, we examined the effects of these two diarylheptanoids on apoptosis, cell cycle regulation and differentiation in neuroblastoma cell lines in this study.
Materials and Methods
Materials. Cpd 1 and Cpd 2 (Figure 1A) were isolated from a methanolic extract of the rhizome of A. officinarum as described in our previous report (7) and were stored as 50 mM stock solutions in 100% dimethyl sulfoxide (DMSO) (final concentration of DMSO 0.2%). Curcumin and 13-cis-retinoic acid were purchased from Wako (Osaka, Japan) and dissolved in DMSO (final concentration of DMSO 0.2%). 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide (MTT), Hoechst 33342 and anti-β-tubulin were purchased from Sigma (St. Louis, MO, USA). Rabbit polyclonal cleaved caspase-3 (Asp175) antibody and rabbit polyclonal caspase-9 antibody were purchased from Cell Signaling (Beverley, MA, USA).
Cell culture. Human neuroblastoma cells are classified into three subtypes: N-type (neuroblastic), S-type (non-neuronal) and I-type (intermediate between the N and S). N-type cells are highly invasive, while S-type cells are noninvasive. Some I-type cells behave as stem cells. IMR-32 (N-type) and SK-N-SH (interconversion between the N and S (13)) cells were purchased from Riken cell bank, Japan. NB-39 cells (N-type with undifferentiated cell state (14)) were provided by Dr. Toshimitsu Suzuki (Fukushima Medical University, Japan). IMR-32, SK-N-SH and NB-39 cells were maintained in RPMI-1640 medium (Invitrogen, Carlsbad, CA, USA) supplemented with 100 U/mL penicillin, 100 μg/mL streptomycin and 10% fetal bovine serum (FBS) (Invitrogen). The cells were maintained at 37°C/5% CO2 in a humid environment.
MTT assay. IMR-32, NB-39 cells (1×104 cells/well) and SK-N-SH (2×104 cells/well) were spread onto a 96-well culture plate with phenol red-free RPMI-1640 medium (with 10% FBS) and maintained for 24 h. Cpd 1, Cpd 2 (final concentration 10−8-10−4 M) or vehicle were added and the cells further incubated for 48 h. After addition of 0.5% MTT solution as a 1/10 volume of the medium in the well, incubation was continued for a further 3 h at 37°C/5% CO2. An equal volume of stop solution (0.04 N HCl in isopropanol) to that of the culture medium was then added to each well and the absorbance at 570 nm (peak) and 655 nm (trough) was measured after thorough pipetting to disperse the generated blue formazan.

Diarylheptanoids induce apoptosis through mitochondrial pathway in neuroblastoma cells. A, IMR-32 cells were treated with 10−5 M diarylheptanoids for 24 h. Nuclei of the cells were stained with Hoechst 33342. Upper panels show the phase-contrast image and lower panels show the fluorescent image from the same view. B, Western blotting analysis for caspase-3 and caspase-9 activation. IMR-32 cells were treated with 10−6 M diarylheptanoids for 0-48 h, and then active caspase-3 and active caspase-9 was assayed by Western blotting. β-Tubulin was used as a loading control.
Hoechst 33342 staining. IMR-32 cells (2×105 cells/well) were spread onto 6-well cell culture plates with RPMI-1640 medium and incubated at 37°C/5% CO2 for 24 h. Cpd 1 or Cpd 2 (final concentration 1×10−6-1×10−4 M) were added and cells were incubated for 24 h. Hoechst 33342 solution (final concentration 0.001% of medium) was added to the wells 15 min before the end of exposure. Morphological changes were observed with a fluorescence microscope (BH2-RFL-T3; Olympus, Tokyo, Japan).
Western blotting. IMR-32 cells (3×106 cells/dish) were spread onto 60-mm cell culture dishes with RPMI-1640 medium and incubated at 37°C/5% CO2 for 24 h. Cpd 1 or Cpd 2 (final concentration 1×10−6 M) were added and cells were incubated for 2, 4, 8, 24 or 48 h. Cells were collected and lysed with lysis buffer. Protein concentration was determined with a Protein Assay Rapid Kit (Wako). Cell lysates containing 20 μg of total protein were loaded onto 15% sodium dodecyl sulfate (SDS)-polyacrylamide gels with Tris/glycine running buffer and transferred to polyvinylidene difluoride (PVDF) membranes. Each membrane was blocked with blocking buffer (5% skim milk in Tween-TBS) for 1 h at room temperature and incubated with the primary antibody (anti-cleaved caspase-3 1:1,000, anti-caspase-9 1:1,000, β-tubulin 1:10,000; diluted in Tween-TBS (TTBS) containing 5% bovine serum albumin (BSA)) at 4°C overnight. After being washed with TTBS, the membrane was incubated with a secondary antibody conjugated with horseradish peroxidase (1:10,000, diluted in 5% skim milk) at room temperature for 1 h, and the signal was detected with an ECL Western blotting detection system (GE Healthcare, Buckinghamshire, UK).

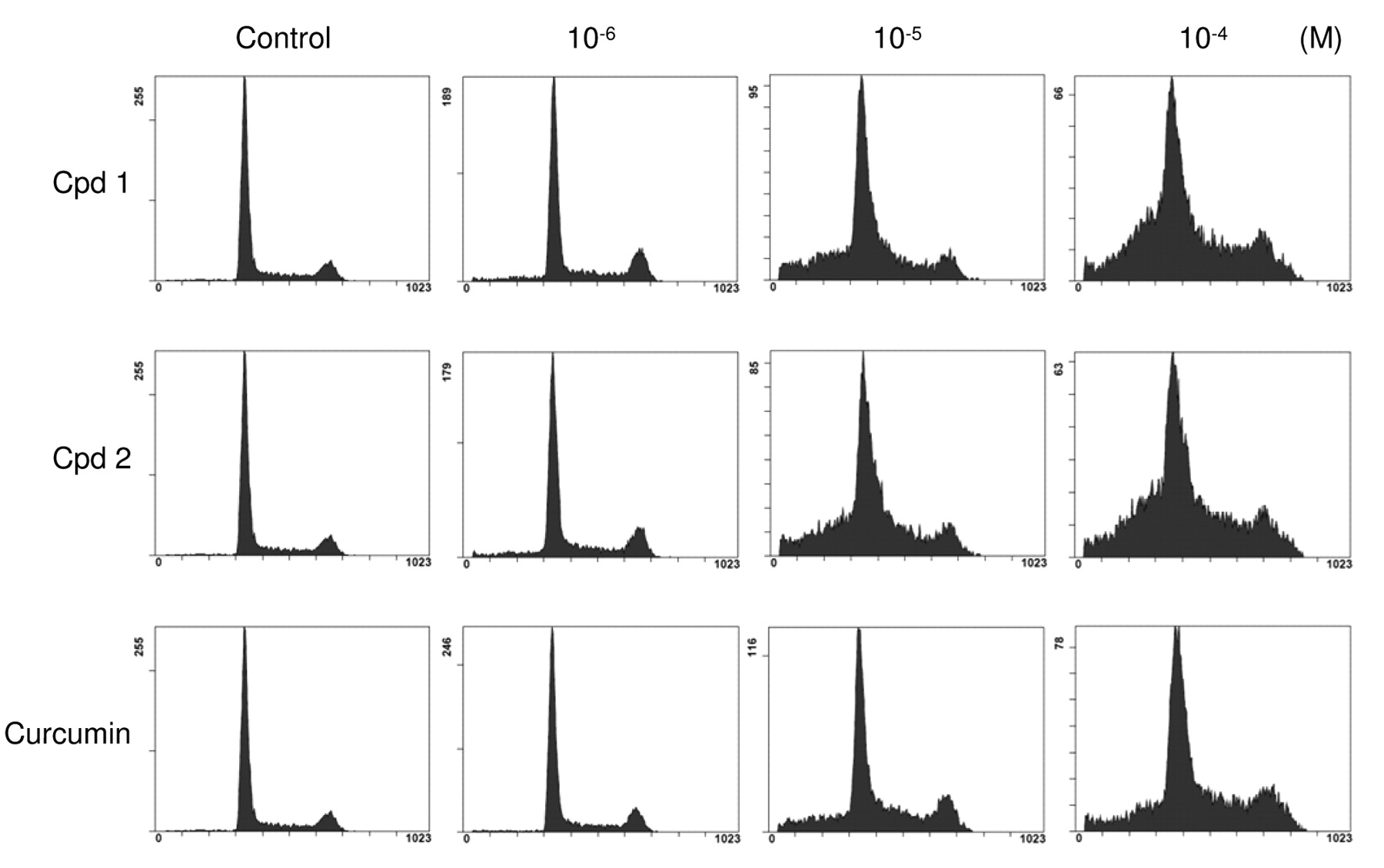
DNA content of IMR-32 cells after application of diarylheptanoids as shown by flow cytometric analysis. IMR-32 cells were treated with Cpd 1, Cpd 2, and curcumin (10−6-10−4 M) for 48 h and stained with PI. Vertical axes indicate cell number and horizontal axes indicate DNA content.
Flow cytometry. IMR-32 cells (2×106 cells/dish) were spread onto 60-mm cell culture dishes with RPMI-1640 medium and incubated at 37°C/5% CO2 for 24 h. Cpd 1, Cpd 2, curcumin (final concentration 1×10−6-1×10−4 M), or vehicle were added and cells were incubated for 48 h. Cells were collected by trypsinization and washed with PBS. Cells were then fixed with ice-cold 70% ethanol for 2 h. After washing with PBS, cells were treated with 0.25 mg/mL RNase solution for 30 min at 37°C. A total of 500 μg/mL propidium iodide (PI) solution was added and cells were incubated for 30 min at 4°C in the dark. The cell samples were analyzed with a FC500 flow cytometer (Beckman Coulter, Fullerton, CA, USA) using the FL4 range.
Differentiation. NB-39 cells are suitable for observation of differentiated cell morphology, since the cells maintain an undifferentiated state (14) and show separated round-cell shapes on the culture dish. NB-39 cells (1×105 cells/dish) were spread onto a human fibronectin Cellware 60-mm dish (BD Bioscience, Bedford, MA, USA) with low serum (1% FBS) RPMI-1640 medium and incubated at 37°C/5% CO2 for 24 h. Cpd 1, Cpd 2, 13-cis-retinoic acid (final concentration 1×10−9-1×10−6 M), or vehicle were added and cells were incubated for 96 h. The cells were observed and photographed under a microscope (BH2-RFL-T3, Olympus). The images were randomized and the number of total cells, cells with neurite outgrowth (>2x soma length), cells with branched neurites and Schwann-like cells were counted in a blinded fashion.
Statistical analysis. Data are given as means±S.E.M (n=3). Significance testing was performed using one-way analysis of variance (ANOVA) followed by Bonferroni's test for comparing three or more data, or Student's t-test in comparing two data.
Results
Cytotoxicity. Previously, we have shown that diarylheptanoids derived from A. officinarum, including Cpd 1 and Cpd 2, have cytotoxic effects on human neuroblastoma IMR-32 cells (7). In this study, we examined in detail the cytotoxic effects of Cpd 1 and Cpd 2 on human neuroblastoma SK-N-SH and NB-39 cell lines as well as on IMR-32 cells (Figure 1B). Consistent with our previous report, Cpd 1 and Cpd 2 showed potent cytotoxicity against IMR-32. For SK-N-SH and NB-39, these diarylheptanoids also showed significant cytotoxicity although their activities were weaker than that against IMR-32. The cytotoxicity of Cpd 1 was comparable to that of Cpd 2 in all cell lines.

Morphological differentiation of neuroblastoma NB-39 cells induced by diarylheptanoids. A, Phase-contrast images of NB-39 cells treated with Cpd 1 (10−8 M), Cpd 2 (10−8 M), 13-cis-retinoic acid (10−7 M, positive control), and DMSO (vehicle control) for 96 h. B, Cells with neurite outgrowth, branching neurites, and Schwann cell-like morphology were counted in a blind study. Vertical axes indicate numbers expressed as a % of total cell number. N: Normal control, V: vehicle control, P: positive control (13-cis-retinoic acid, 10−7 M); *p<0.05, **p<0.01 versus vehicle control.
Apoptosis. Subsequently, we analyzed the apoptosis-inducing activity of diarylheptanoids in IMR-32 cells, which are highly sensitive to these compounds. Both compounds induced cell condensation and nuclear shrinking and fragmentation, morphological features of apoptosis (Figure 2A). In Western blot analysis, we examined activation of caspase-3, a pivotal protease in apoptosis induction, and caspase-9, a protease involved in mitochondrial apoptosis (Figure 2B). Both Cpd 1 and Cpd 2 activated caspase-3 and caspase-9 after application for >24 h at a concentration of 10−6 M.
Cell cycle arrest. Since conventional anticancer agents act on cell cycle-controlling mechanisms concurrently with induction of apoptosis, we assayed the DNA content of IMR-32 cells treated with Cpd 1, Cpd 2, and curcumin, a well-characterized representative diarylheptanoid, for 48 h using flow cytometry to clarify the cell cycle status. Cpd 1 and Cpd 2 increased the S-phase cell population concurrently with the appearance of a sub-G1 cell population in a concentration-dependent manner (Figure 3). Consequently, treatment of IMR-32 neuroblastoma cells with these diarylheptanoids induces both S-phase cell cycle arrest and apoptosis. Curcumin also increased populations of S-phase and sub-G1 cells, but its activity was relatively modest compared with those of Cpd 1 and Cpd 2 (Figure 3). In addition, flow cytometry at 24 h after application showed that Cpd 1 and Cpd 2, but not curcumin, at concentrations of 10−6 M induced a significant increase in the S-phase cell population (data not shown).
Differentiation. The data for cell cycle arrest led us to hypothesize that diarylheptanoids derived from A. officinarum cause differentiation of malignant neuroblastoma cells to mature neural or glial cells. To examine this possibility, we assessed the morphological features of neuroblastoma NB-39 cells after 96 h of treatment with Cpd 1, Cpd 2, and with 13-cis-retinoic acid as a positive control (Figure 4). NB-39 cells have a rounded cell shape and are suitable for cell shape assessment by counting, and have a relatively low sensitivity to these compounds (Figure 1B). As shown in Figure 4, Cpd 1 at 10−8 M showed a tendency to cause elongation of neurites and also caused a significant increase in neurite branching, the latter effects comparable to that of 13-cis-retinoic acid. Cpd 2 also showed a tendency to cause neurite outgrowth and branching with a peak at 10−8 M, but the effects were not significant. On the other hand, Schwann cell-like differentiation, a feature of treatment with 13-cis-retinoic acid, was unaffected by treatment with either Cpd 1 or Cpd 2. It is likely that the reduction of parameters at 10−6 M shown in Figure 4B is a result of the cytotoxicity of the test compounds at this concentration.
Discussion
Both Cpd 1 and Cpd 2 induced significant cell death in all neuroblastoma cell lines, although IMR-32 cells were more sensitive to diarylheptanoids than were the other cell lines (SK-N-SH, NB-39). Even though the differences in sensitivity among the cell lines should be taken into consideration, Cpd 1 and Cpd 2 were more potent cytotoxins than the other diarylheptanoids derived from A. officinarum (7, 15). The mechanism of cell death involves induction of apoptosis via the mitochondrial pathway. Recently, loss of caspase-8 expression was reported in patients with highly malignant neuroblastoma and in established neuroblastoma cell lines, including IMR-32 (16, 17). Caspase-8 is involved in the mechanism of apoptosis induced by curcumin in other cancer cells (18, 19). On the other hand, activation of p53 and inhibition of nuclear factor (NF)-κB have been reported to contribute to curcumin-induced apoptosis in human neuroblastoma cells (8, 9). Inhibition of proinflammatory mediators by Cpd 1 occurs by reduction of NF-κB activation and phosphorylation of extracellular signal-regulated kinase (ERK) (11). Taken together, the apoptosis induced by Cpd 1 in caspase-8-lacking IMR-32 cells is due, at least in part, to inhibition of NF-κB and activation of mitochondrial apoptotic pathway via caspase-9 cleavage. Diarylheptanoids derived from A. officinarum would be expected to induce apoptosis in clinically malignant neuroblastomas lacking caspase-8.
S-Phase cell cycle arrest was observed in IMR-32 cells after application of Cpd 1 and Cpd 2, concurrently with apoptosis induction. Tian et al. also reported the apoptosis-inducing and S-phase-arresting effects in neuroblastoma cells of a diarylheptanoid derived from A. officinarum, although with a chemical structure different from that of Cpd 1 and Cpd 2, and they suggested that activating transcription factor 3 (ATF3) up-regulation and p53 stabilization may contribute to these effects (20). The findings of our study in regard to cell cycle arrest corroborate those of Tian et al. Curcumin shows various patterns of effects on cell cycle, arresting human mantle cell lymphoma cells in G1/S-phase (21) and human bladder cancer and glioma cells in G2/M-phase (22, 23). Moreover, it was reported that a methanolic extract of A. officinarum inhibited H2O2-induced apoptosis and concurrently arrested the cell cycle in G2/M-phase, but not in S-phase, in Chinese hamster lung fibroblast V79-4 cells (24). These results indicate that compounds possessing analogous diarylheptanoid structures do not necessarily induce the same effects on the cell cycle, and that effects may also differ among different kinds of cancer cells.
Recently, a clinical trial in malignant neuroblastoma reported the efficacy of 13-cis-retinoic acid as differentiation-inducing therapy (2, 25). In our study, Cpd 1 induced cell branching, as a differentiation feature, in the NB-39 neuroblastoma cell line, comparable to that induced by 13-cis-retinoic acid. On the other hand, unlike 13-cis-retinoic acid, differentiation into Schwann-like cells was not increased by diarylheptanoids. This discrepancy may be the result of differences in the differentiation-inducing mechanisms of Cpd 1 and 13-cis-retinoic acid, suggesting that more effective differentiation-inducing therapy may result from the combined application of Cpd 1 and 13-cis-retinoic acid. Contrary to the differentiation-promoting activity of retinoic acid, fenretinide, a synthetic retinoid, induced apoptosis in neuroblastoma cells (26). In our study, Cpd 1-treated cells showed both aspects of apoptosis and differentiation, suggesting that Cpd 1 may be able to control malignant neuroblastoma by multiple mechanisms.
In conclusion, we examined the antitumor effects of two diarylheptanoids, Cpd 1 and Cpd 2, against neuroblastoma cells. Both compounds induced mitochondrial apoptosis and S-phase cell cycle arrest, and Cpd 1 caused significant neural differentiation when applied at 10−8 M. These results indicate that diarylheptanoids isolated from A. officinarum have multiple antitumor effects in neuroblastoma cell lines. In consequence, diarylheptanoids appear to be useful candidate drugs for chemotherapy of advanced neuroblastoma.
Acknowledgements
We thank Dr. Toshimitsu Suzuki, Fukushima Medical University School of Medicine, for providing NB-39 cells. This work was supported by the ‘Academic Frontier’ Project for Private Universities: matching fund subsidy from MEXT (Ministry of Education, Culture, Sports, Science and Technology) 2007-2009, and a Joint Research Grant from Nihon University College of Pharmacy 2005-2009.
- Received June 24, 2009.
- Revision received November 13, 2009.
- Accepted November 23, 2009.
- Copyright© 2009 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.