


Abstract
Prostate cancer (CaP) is a significant cause of death in American men. While men residing in Asia show a lower incidence of hormone-refractory prostate cancer (HRPC) compared to Caucasian males, Asian men who move to and live in the United States and adopt a western lifestyle have HRPC rates indistinguishable from Caucasian males. These findings suggest that Asian diets contain ingredients that might protect against the development of HRPC. The identity and mechanisms of such HRPC protective agents remain to be elucidated. An Asian diet may confer protection against HRPC owing to functional synergy between bioactive dietary agents, thus broadening the chemopreventive index, with increased distinct anticancer properties and decreased untoward effects. Here, whether or not a combination of epigallocatechin gallate (EGCG), genistein and quercetin, phytochemicals present in a traditional Asian diet, might exert synergy in controlling proliferation and gene expression was investigated in CWR22Rv1 CaP cells, an in vitro model mimicking CaP transition from AD (androgen dependence) to HRPC. Cell proliferation was inhibited ~40%, ~15% and ~20%, respectively by 2.5 μM EGCG, genistein and quercetin used alone. The co-administration of 2.5 μM of these phytochemicals suppressed proliferation synergistically in the CWR22Rv1 cells maintained in RPMI-1640 supplemented with 10% fetal bovine serum, but not in the cells maintained as serum-free cultures. Synergy in the expression of androgen receptor, tumor suppressor p53 and detoxification enzyme quinone reductase type 1, denoted NQO1, was also observed for the combined phytochemicals. These results demonstrate the feasibility of developing a diet-based combinatorial approach for CaP prevention and treatment and raise the possibility that serum added to culture medium might affect uptake, bioavailability and biological efficacy of dietary phytochemicals.
Prostate cancer (CaP) is the most commonly diagnosed non-cutaneous cancer in males and a leading cause of death in American men (1-3). Due to an ageing population worldwide and increased vigilance in prostate screening, trends of sustained rise in both incidence and mortality from CaP have also become evident in developing countries (2, 4-7).
Although latent CaP occurs with similar frequency in men with different ethnic backgrounds and residing in culturally diverse geographical locations, the incidence and mortality rates of clinical CaP are significantly higher for men living in the United States and many developed countries, compared to men residing in Asia, e.g. China and Japan (2, 8, 9). However, when Chinese and Japanese men move to and live in North America and adopt western diets and lifestyles, they show clinical CaP at rates comparable to Caucasian men (10-12). These data are supported by epidemiological studies showing that men residing in Asia have an inverse relationship between HRPC risk and consumption of traditional Asian diets rich in tea, soybean products etc. (2, 9, 13, 14) and suggest that traditional Asian diets confer protection against CaP transition to a hormone-refractory state denoted HRPC. HRPC is the most devastating form of CaP and represents the terminal stage of transition from androgen dependence (AD). At present, no curative therapy exists for HRPC, therefore, prevention, postponement or aversion of HRPC, or the maintenance of AD, would produce chronic instead of terminal CaP. Typical Asian diets are low in animal fat and high in fish, soy, fruits and vegetables and green tea considered healthy since they are rich in polyphenols, flavonoids and other phytochemicals with chemopreventive activities. As an example, genistein from soy has demonstrated breast and prostate cancer preventive activities (15-18). However, for the most part, the reported anti-tumor activities of phytochemicals are primarily based on in vitro studies. The purported benefits of healthy dietary agents are challenged by the uncertain results regarding the lowering of cancer risk that were obtained in large-scale intervention studies using specific single dietary ingredients at supraphysiological doses (19-23).
Most mechanistic studies on food-derived chemopreventive agents to date have used single dietary ingredients at concentrations unlikely to be achieved by food consumption (21, 24-27). We surmise that cancer protection by healthy diets may be attributable to synergy between low dose phytochemicals, for which little information or evidence currently exists (28, 29). In this study, the concept of functional synergy was tested by investigating the combination of EGCG (epigallocatechin gallate) and genistein, derived from tea and soy products commonly found in a traditional Asian diet, with quercetin, present in abundance in fruits and vegetables (30-32), for efficacy against CaP. Each chosen agent reportedly has shown anti-CaP activities, with overlapping and distinct molecular actions and targets. For example, EGCG acts at G1/S whereas genistein affects the G2/M checkpoint of the cell cycle (33, 34). EGCG exerts epigenetic control by inhibiting DNA methyl-transferases (35), an effect not reported for genistein or quercetin. Because these phytochemicals have non-overlapping activities, whether or not they exert synergistic interactions was investigated by exposing CWR22Rv1 cells to a suboptimal dose of each phytochemical, alone or in combination, followed by the determination of cell proliferation, colony formation and the expression of specific genes.
Materials and Methods
Reagents. EGCG, genistein and quercetin were purchased from LKT Laboratories (St Paul, MN, USA). The primary antibodies anti-androgen receptor (AR), anti-estrogen receptor β (ERβ), anti-p53, anti-NQO1 (detoxification enzyme quinone reductase type 1), anti-NQO2 (a structural homolog of NQO1 thought to be involved in detoxification but may have other cellular functions), anti-actin and secondary antibodies were purchased from Santa Cruz Biotechnology Inc. (Santa Cruz, CA, USA). Fetal bovine serum (FBS), RPMI-1640, penicillin and streptomycin were purchased from Cellgro Inc. (Herndon, VA, USA). All other chemicals and solvents used were of analytical grade.
Cell culture. Human CWR22Rv1 cells were obtained from the American Tissue Culture Collection (Manassas, VA, USA) and maintained in RPMI-1640 supplemented with penicillin, streptomycin and 10% heat inactivated FBS in a humidified atmosphere of 5% CO2 in air at 37°C, as previously described (36-38).
Cell growth inhibition assay and colony formation assay. For the cell proliferation assay, the CWR22Rv1 cells were plated in 6-well plates at a seeding density of 5×104 cells/ml. EGCG, genistein or quercetin dissolved in DMSO, added to the culture media to the final concentration specified. Vehicle added alone served as the untreated control. After 72 h, the untreated control and treated cells were harvested by trypsinization and cell numbers were determined using a hemocytometer. Cell viability was assayed by trypan blue dye exclusion (36-38). The harvested cells were washed twice with PBS, and pellets were stored at −80°C for further analysis. The anchorage dependent clonogenicity assay was performed as described previously with some modification (37). Cells (800-1200 cells/ml, 2 ml/well) were added to 6-well tissue culture plates containing varying concentrations of EGCG, genistein and quercetin, alone or in combination, or vehicle alone as the control followed by an additional 8-day incubation to allow colonies to form. The colonies were fixed and stained with 1.25% crystal violet, washed extensively to remove excess dye and imaged using a Hewlett-Packard scanner. Quantitative changes in clonogenicity were determined by extraction colonies in individual tissue culture plates with 0.5 ml 10% acetic acid and measuring the absorbance of the extracted dye at 450 nm. The experiments were performed in triplicate.
Immunoblot analysis. For Western blot analysis, the cells collected by centrifugation were lysed in ice-cold radio immunoprecipitation assay (RIPA) buffer (50 mM Tris, pH 7.4, 150 mM NaCl, 1 mM EDTA, 1% Triton X-100, 1% deoxycholate, 0.1 % SDS, 1 mM dithiothreitol and 10 μl/ml protease inhibitor cocktail). The extracts were centrifuged and the clear supernatants were stored in aliquots at −80°C for further analysis. The protein content of the cell lysates was determined by a Coomassie protein assay kit (Pierce, Rockford, IL, USA) using BSA as standard. Aliquots of the lysates (20 μg of protein) were boiled with sample buffer for 5 min and resolved by 10% SDS-PAGE. The proteins were transferred to a nitrocellulose membrane and blocked in TBST buffer (10 mM Tris, pH 7.5, 100 mM NaCl and 0.05% Tween 20) containing 3% nonfat dried milk, overnight at 4°C. The blots were incubated with various primary antibodies, followed by incubation for 1 h with appropriate secondary antibodies conjugated to horseradish peroxidase in TBST. Actin expression was used as the loading control. The intensity of the specific immunoreactive bands was detected by enhanced chemiluminescence (ECL), using the manufacturer's protocol (KPL Inc., Gaithersburg, MS, USA), quantified by densitometry and normalized against the loading control actin, as previously described (39).
Statistical analysis. The results were expressed as mean±standard deviation (SD). The significance between the control and treated groups was determined by Student's t-test and p-values less than 0.05 were taken as significant in all the cell growth and colony formation experiments.
Results
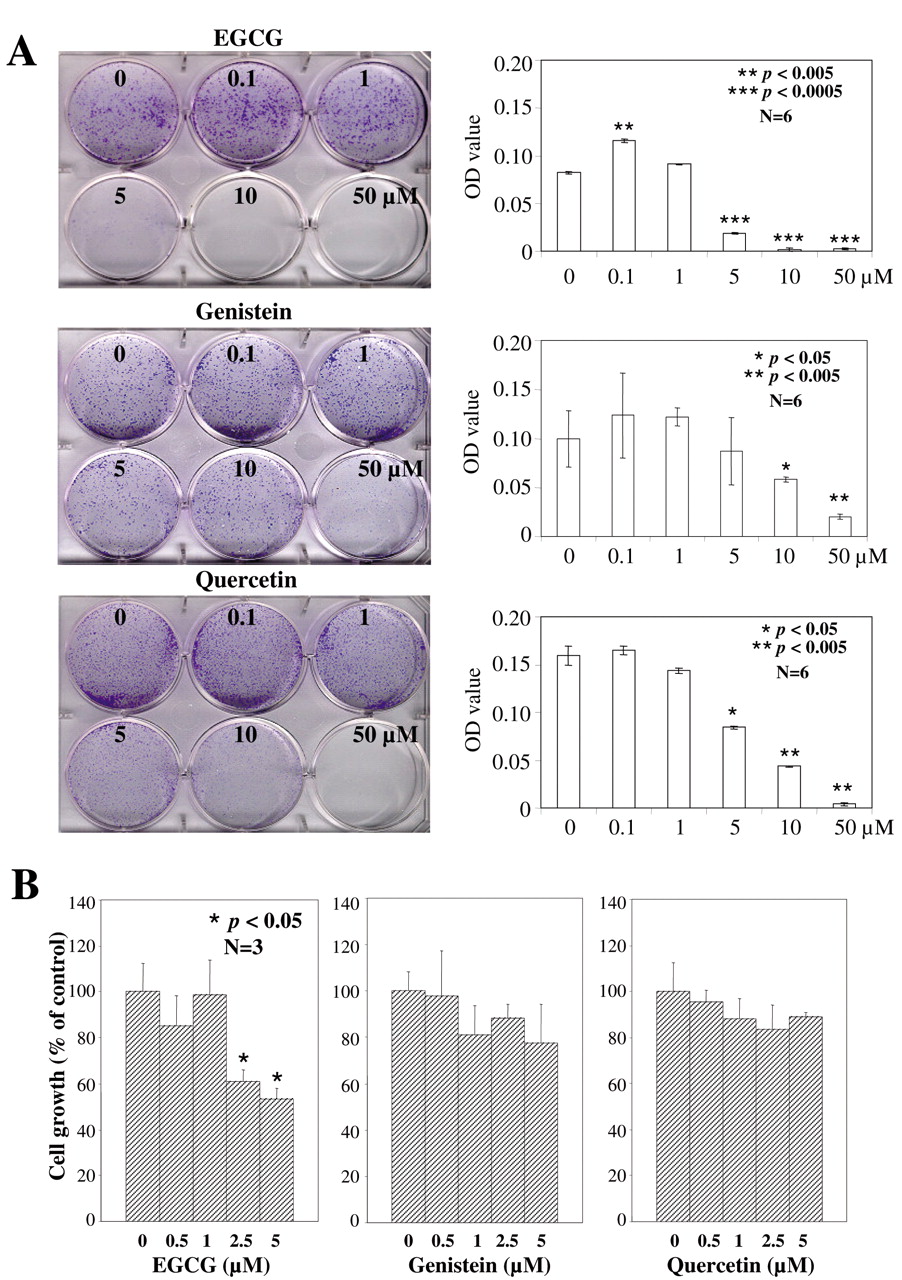
Effect of EGCG, genistein, or quercetin on colony formation, and cell proliferation. The CWR22Rv1 cells were incubated with 0, 0.1, 1, 5, 10 or 50 μM EGCG, genistein or quercetin. The phytochemicals exerted different activities on colony formation. EGCG and quercetin as single agents significantly inhibited the ability of the CWR22Rv1 cells to form colonies at doses ≥5 μM, whereas 5 μM genistein produced affects not significantly different from the vehicle treated control cells (Figure 1A). Using this as the upper limit of the dose of phytochemical added to the culture medium, the ability of EGCG, genistein and quercetin to suppress CWR22Rv1 cell proliferation was tested. Following incubation with 0, 0.5, 1, 2.5 or 5 μM of EGCG, genistein or quercetin for 72 h, dose-dependent cell growth suppression was observed with all three agents. In the EGCG-treated cells, 2.5 μM EGCG elicited a >40% suppression in proliferation, p<0.05 compared to the control (Figure 1B) while identical concentrations of genistein and quercetin were ineffective (Figure 1B). Analyzing the results from colony formation and cell proliferation together, the relative potency and efficacy of the three phytochemicals was ranked as EGCG>quercetin>genistein. The results also suggested, as hypothesized, that each phytochemical acted by a different mechanism and therefore when combined might be functionally complementary and generate synergy. Equally plausibly antagonism might result from the addition of these phytochemicals as a group to cultured CWR22Rv1 cells.

Colony formation and cell growth in CWR22Rv1 cells treated with EGCG, genistein or quercetin in RPMI-1640 containing 10% FBS. A, Colony formation assay, quantified by spectrophotometer at O.D. 450. B, Cell numbers determined by hemocytometer after 72 h incubation. Values are expressed as mean±SD from three experiments and the asterisk indicates statistical significance compared to the control.
Effect of EGCG, genistein, or quercetin, alone or combined on cell proliferation and specific gene expression in cells cultured in 10% FBS. To test whether functional synergy or antagonism exists between EGCG, genistein and quercetin, experiments were performed at a suboptimal dose of each phytochemical, used alone or in combination. Since 2.5 μM EGCG sufficed to significantly inhibit proliferation of the CWR22Rv1 cells, this suboptimal dose was chosen for each phytochemical for the combination study. Compared to single phytochemical alone (Figure 1B), no potentiation of suppression of cell proliferation was evident in the groups combining two phytochemicals at the 2.5 μM dose (Figure 2A), while a substantially more pronounced antiproliferative effect was observed in the cultures exposed to all three phytochemicals (Figure 2A).
Western blot analysis was performed to investigate changes in gene expression. It was observed that addition of 2.5 μM EGCG or quercetin (2.5 μM) separately, inhibited AR expression by 67% and 47%, respectively compared to the control, compared to 22% reduction in AR levels for the cells treated with 2.5 μM genistein (Figure 2B). Surprisingly, the cells treated with groups of two or three phytochemicals increased AR expression by 2- to 4-fold without inducing a commensurate change of ERβ expression (Figure 2B).
Minimum changes of p53 expression were observed when the cells were treated with the single phytochemicals while combinations of two or three phytochemicals markedly up-regulated p53 expression (Figure 2B). Results in Figure 2B show augmented induction of NQO1 expression in cells exposed to two or three phytochemicals together. In contrast, the expression of NQO2 was minimally affected by the same treatment conditions. Taken together, these results support the existence of synergy between EGCG, genistein and quercetin in the control of AR, p53 and NQO1 expression.

Cell growth and specific gene expression in CWR22Rv1 cells treated for 72 h with 2.5 μM EGCG, genistein and quercetin in RPMI-1640 containing 10% FBS. A, Growth response with 2 or 3 agents together. B, Western blot analysis of AR, ERβ, p53, NQO1 and NQO2 protein expression. Actin was used as a loading control. The intensity of the specific immunoreactive bands was quantified by densitometry and expressed as a fold difference against actin. Values are mean±SD from three experiments.
Effect of EGCG, genistein, or quercetin alone or in combination on cell proliferation, cell viability and specific gene expression in cells cultured in serum-free conditions. To test whether culture conditions might affect synergy or antagonism between the phytochemicals, the influence of FBS was investigated. Cells continuously maintained in media supplemented with 10% FBS were switched to serum-free conditions and cultured for an additional 72 h, with or without the addition of 2.5 μM of each phytochemical alone or in combination. A more substantial inhibition in proliferation was observed in the serum-free cultures in response to treatment with each phytochemical individually (Figure 3A). However, no synergistic suppression of proliferation was found in the serum-free cultures exposed to combined phytochemicals (Figure 3A). Interestingly, when cell viability was used as an assay, synergy was observed in the combination of EGCG with either genistein or quercetin, but not in the combination of genistein with quercetin (Figure 3B). Moreover, no additional synergy on cell viability occurred when the cells were exposed to all three phytochemicals combined (Figure 3B).

Effects of EGCG, genistein and quercetin on cell growth, cell viability and protein expression in CWR22Rv1 cells cultured in serum-free RPMI-1640 medium. Cells were treated with 2.5 μM of EGCG, genistein or quercetin alone or in combination for 72 h. A, Cell numbers determined by hemocytometer. B, Cell viability measured using the trypan blue dye exclusion assay. C, Western blot analysis of protein expression. Actin was used as a loading control. The intensity of the specific immunoreactive bands was quantified by densitometry and expressed as a fold difference against actin.
By Western blot analysis, EGCG and quercetin used alone inhibited the AR expression by 55% and 75%, respectively, compared to the control, whereas genistein surprisingly elevated the AR levels by ~2.45-fold (Figure 3C). This induction of AR by genistein was completely abrogated in the cells exposed to genistein combined with either EGCG or quercetin (Figure 3C). As with the FBS-supplemented cell cultures, the ERβ expression was minimally affected in the serum-free cultures by the phytochemicals, alone and in combination (Figure 3C).
Only minimum changes in p53 levels were observed in the cells treated with single phytochemicals (Figure 3C). The combination of EGCG and quercetin resulted in ~69% reduced expression of p53, whereas the combination of genistein and quercetin increased the p53 levels by ~42% (Figure 3C). With all three phytochemicals combined, no additional synergy on p53 expression was evident (Figure 3C). Genistein and quercetin, as individual phytochemicals, increased NQO1 expression by ~3.78- and ~6.42-fold, respectively, but EGCG had the minimum effect (Figure 3C). In contrast, a more prominent increase was found in the induction of NQO2 in the cells exposed to the combination of two and three phytochemicals (Figure 3C).
Discussion
The transition from AD to HRPC involves a myriad of genetic and epigenetic changes, which might be optimally sensitive to a combination of dietary ingredients with distinct as well as overlapping activities. While the low dose of the bioactive agents chosen for the present studies still may not be easily achievable physiologically, it was nevertheless much lower than the efficacious dose used for most in vitro studies to date (40-44). Since, in addition to the choice of dietary agents, other factors such as culture conditions, stability and uptake of these bioactive dietary ingredients might also influence their efficacy, the effects of culturing cells in serum-free media versus media containing 10% FBS were compared. When all three phytochemicals were co-administered, functional synergy on growth suppression was only observed in the cells maintained in 10% FBS and not in the cells cultured in serum-free conditions. Nevertheless, a significant reduction in cell proliferation was observed in the cells treated with the single phytochemicals and when two phytochemicals were used together. It is possible that the presence of serum interferes with the binding or uptake of phytochemicals, particularly at the low dose used thereby lowering the bioavailability and hence efficacy of each phytochemical whether used alone and in combination. One might extrapolate from these findings that challenges remain for a priori determination of the factors possibly impinging on phytochemical interactions and bioavailability.
Both AR and ERβ play important roles in the control of prostate cancer cell proliferation and the culture conditions, as reflected by the presence of serum, had a critical role in modulating AR expression in the CWR22Rv1 cells. For instance, when all three phytochemicals were co-administered at 2.5 μM, a 3.89-fold up-regulation of AR expression was observed in the cells cultured in 10% FBS, which starkly contrasted with the 70% suppression of AR expression under the same treatment conditions using serum-free cell cultures. In contrast, the co-administration of all three phytochemicals suppressed proliferation in the cells under both culture condition, suggesting that different mechanisms were at play regarding the control of AR and cell proliferation. Because studies have shown that the overexpression of wild-type p53 reduces androgen function (45) and since the stability of p53 is subject to regulation by the phase II detoxification enzymes NQO1 and NQO2 (46, 47), changes in the levels of these three genes were also assessed. Synergy was observed between EGCG, genistein and quercetin in regards to the control of AR, p53 and NQO1 in the 10% FBS and serum-free cultures. One might speculate that the combined phytochemicals might affect the balance between AR, p53, NQO1 and NQO2 and disrupt or even obliterate the interplay between this group of genes during AD to HRPC progression such as to deter AR-independent cell proliferation and gene expression, a hypothesis under active consideration and experimentation in our laboratory.
Acknowledgements
We thank Dr. Elangovan Selvakumar for his help with the Western blot analysis. This research was supported in part by NIH RO3 121364 to J.M.W.
- Received April 10, 2009.
- Revision received June 22, 2009.
- Accepted July 27, 2009.
- Copyright© 2009 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Quercetin Targets hnRNPA1 to Overcome Enzalutamide Resistance in Prostate Cancer Cells
- Effect of Combined Treatment with Ursolic Acid and Resveratrol on Skin Tumor Promotion by 12-O-Tetradecanoylphorbol-13-Acetate
- The Molecular Basis for the Pharmacokinetics and Pharmacodynamics of Curcumin and Its Metabolites in Relation to Cancer