


Abstract
We have previously reported that tropolone (T-3), 2,4-dibromo-7-methoxytropone (T-21), diethyl 2-chloroazulene-1,3-dicarboxylate (A-9), 1,3-difluoroazulene (A-11), 3-morpholino-1,5-azulenequinone (AQ-8) and 3,7-dibromo-1,5-azulenequinone (AQ-13) inhibited the nitric oxide (NO) production of lipopolysaccharide (LPS)-activated mouse macrophage-like RAW264.7 cells, with or without the inhibition of inducible NO synthase (iNOS) mRNA and protein expression. In order to confirm the anti-inflammatory potency, possible effects on prostaglandin (PG) E2 production and the expression of enzymes involved in the arachidonic acid pathway were investigated. Among these six compounds, only A-9 effectively inhibited the PGE2 production of the LPS-stimulated RAW264.7 cells. Western blot analysis demonstrated that A-9 inhibited phospholipase A2 (PLA2), cyclooxygenase (COX)-2 and iNOS proteins only by 12, 45 and 42%, respectively. These data demonstrate the lack of correlation between the extent of inhibition of iNOS protein expression by tropolone or azulene derivatives and that of PGE2, and suggest the possible anti-inflammatory potency of A-9.
- Diethyl 2-chloroazulene-1,3-dicarboxylate
- RAW264.7 cells
- macrophage
- PGE2
- COX-2
- PLA2
- anti-inflammatory effect
We have initiated the structure-activity study of tropolone and azulene derivatives. Among 107 compounds so far analyzed, tropolone (T-3), 2,4-dibromo-7-methoxytropone (T-21) (1), diethyl 2-chloroazulene-1,3-dicarboxylate (A-9), 1,3-difluoroazulene (A-11) (2), 3-morpholino-1,5-azulenequinone (AQ-8) and 3,7-dibromo-1,5-azulenequinone (AQ-13) (Figure 1) (3) inhibited the nitric oxide (NO) production of lipopolysaccharide (LPS)-activated mouse macrophage-like RAW264.7 cells (Table I), with (1, 3) or without (2) the inhibition of inducible NO synthase (iNOS) mRNA and protein expression. However, the extent of inhibition of iNOS expression differed considerably from compound to compound (1-3). Electron-spin resonance (ESR) spectroscopy showed that none of these compounds scavenged the NO released from 1-hydroxy-2-oxo-3-(N-3-methyl-3-aminopropyl)-3-methyl-1-triazene (NO generator) in the presence of 2-(4-carboxyphenyl)-4,4,5,5-tetramethylimidazoline-1-oxy1-3-oxide (a spin trap agent) (1-3). In order to confirm the anti-inflammatory potency of these six compounds, whether or not they inhibited the prostaglandin (PG) E2 production of LPS-stimulated RAW264.7 cells was first investigated. In the arachidonic acid (AA) cascade, two enzymes are involved in the production of PGE2. PG synthesis begins with the liberation of AA, the prime precursor, from membrane phospholipids by phospholipase A2 (PLA2). Subsequently, cyclooxygenase (COX) catalyzes the rate-limiting reactions for PG synthesis comprising the bis-cyclooxygenation of AA to form PGG2 and the peroxidative reduction of this intermediate to PGH2 (4). The cPLA2α is activated by proinflammatory cytokines or growth factors and catalyzes the AA from the cell surface membrane (5, 6). It is known that COX has two isoforms, COX-1 (constitutive isoform) and COX-2 (inducible isoform) (7). Therefore, the possible effects on the expression of these enzymes were also investigated.
Materials and Methods
Materials. The following chemicals and reagents were obtained from the indicated companies: Dulbecco's modified Eagle's medium (DMEM) (Invitrogen Corp, Carlsbad, CA, USA); fetal bovine serum (FBS) (Gemini Bio-Products, Woodland, CA, USA), LPS from Escherichia coli (Serotype 0111:B4) (Sigma Chemical Ind., St. Louis, MO, USA) and tropolone (T-3) (Tokyo Kasei Kogyo Co., Ltd., Tokyo, Japan).
Synthesis of tropolone, azulene and azulenequinone derivatives. T-21 (8), A-9 (9), A-11 (9), AQ-8 (10, 11) and AQ-13 (10, 12, 13) were prepared, according to the reports in the references cited. Cell culture. The RAW264.7 cells, established from the peritoneal fluid of BALB/c mice and showing the phenotypic characteristics of monocytes and macrophages (14), were subcultured in DMEM supplemented with 10% heat-inactivated FBS under a humidified 5% CO2 atmosphere and incubated with various concentrations of test compounds in the presence or absence of LPS (100 ng/ml).

Structure of compounds used in this study.
Assay for cytotoxic activity. The cytotoxic activity of the azulenes was determined by the MTT method, and expressed as absorbance at 540 nm of the MTT-stained cells. The 50% cytotoxic concentration (CC50) was determined from the dose-response curve (1-3).
Assay for NO concentration. Near confluent RAW264.7 cells were incubated for 24 hours with each test sample in phenol red-free DMEM supplemented with 10% FBS, and the NO production was quantified by Greiss reagent, using the standard curve of NO -2. To eliminate the interaction between the sample and the Greiss reagent, the NO concentration in the culture medium without cells was also measured, and subtracted from the value obtained with the cells. The concentration that inhibited the LPS-stimulated NO production by 50% (50% inhibitory concentration: IC50) was determined from the dose-response curve (1-3). The efficacy of inhibition of NO production was evaluated by the selectivity index (SI), which was calculated by the following equation: SI=CC50 / IC50.
Measurement of PGE2 production. RAW264.7 cells were subcultured in 24-well plates and incubated with various concentrations of test compounds in the presence of LPS (100 ng/ml). The culture supernatant was collected by centrifugation and the PGE2 concentration was determined by EIA kit (Cayman Chemical Co, Ann Arbor, MI, USA).
Inhibition of NO production by LPS-stimulated RAW264.7 cells by tropolone, azulene and azulenequinone derivatives.
Western blotting. The cell pellets were lysed with 100 μl of lysis buffer (10 mM Tris-HCl [pH 7.6], 1% Triton® X-100, 150 mM NaCl, 5 mM EDTA-2Na and 2 mM phenylmethylsulfonyl fluoride (PMSF) for 10 minutes in ice water, and then incubated for 50 minutes at 4°C with RT-5 ROTATOR (Titec, Saitama, Japan). The cell lysates were centrifuged at 16,000 ×g for 20 minutes at 4°C to remove the insoluble materials and the supernatant was collected. The protein concentrations of the supernatant were measured by Protein Assay Kit (Bio Rad, Hercules, CA, USA). An equal amount of the protein from the cell lysates (10 μg) was mixed with 2x sodium dodecyl sulfate (SDS)-sample buffer (0.1 M Tris-HCl [pH 6.8], 20% glycerol, 4% SDS, 0.01% bromphenol blue, 1.2% 2-mercaptoethanol), boiled for 10 minutes, applied to SDS-8% polyacrylamide gel electrophoresis and then transferred to polyvinylidene difluoride (PVDF) membrane. The membranes were blocked with 5% skim milk in phosphate-buffered saline (PBS(-)) plus 0.05% Tween 20 for 90 minutes and incubated for 90 minutes at room temperature with anti-PLA2, anti-COX-2 or anti-iNOS antibody (1:1,000; Santa Cruz Biotechnology, Delaware, CA, USA) or anti-actin antibody (1:2,000-4,000; Sigma) and then incubated with horseradish peroxidase-conjugated anti-mouse or goat IgG for 90 minutes at room temperature. The immunoblots were developed with a Western Lightning™ Chemiluminescence Reagent Plus (Perkin Elmer Life Sciences, Boston, MA, USA) and analyzed on a Macintosh (Power Macintosh 7600/120) computer using the public domain NIH Image program (National Technical Information Service, Springfield, Virginia, USA, part number PB95-500195GEI).

Effect of six selected compounds on PGE2 production by LPS-activated RAW264.7 cells. RAW264.7 cells were incubated for 24 hours without (control), or with the indicated concentrations of tropolone (T-3), 2,4-dibromo-7-methoxytropone (T-21), diethyl 2-chloroazulene-1,3-dicarboxylate (A-9), 1,3-difluoroazulene (A-11), 3-morpholino-1,5-azulenequinone (AQ-8), or 3,7-dibromo-1,5-azulenequinone (AQ-13), and the extracellular concentration of PGE2 was then measured.
Results
Among the six test compounds, only A-9 inhibited the PGE2 production of the LPS-stimulated RAW264.7 cells (Figure 2). A-9 inhibited the PGE2 production dose-dependently. A-9 at 2 or 20 μM inhibited the PGE2 production by 53.2 and 96.5%, respectively (Figure 3). Western blot analysis (Figure 4) demonstrated that A-9 inhibited PLA2, COX-2 and iNOS proteins by only 12, 45 and 42%, respectively.
Discussion
The present study demonstrated that although tropolone (T-3), 2,4-dibromo-7-methoxytropone (T-21), 1,3-difluoroazulene (A-11), 3-morpholino-1,5-azulenequinone (AQ-8) and 3,7-dibromo-1,5-azulenequinone (AQ-13) inhibited NO production by LPS-stimulated mouse macrophage-like cells, these failed to inhibit the PGE2 production of the same cells. This suggested that the inhibition of NO production by macrophages is not an appropriate parameter for evaluating anti-inflammatory activity. On the other hand, diethyl 2-chloroazulene-1,3-dicarboxylate (A-9) inhibited PGE2 production (IC50<2 μM) more efficiently than NO production (IC50=17 μM). These data suggested the importance of further investigating A-9 for its possible anti-inflammatory potential. It was unexpected that A-9 inhibited COX-2 protein expression only by 45% . It also inhibited the iNOS protein expression only partially and only marginally affected the PLA2 protein expression. The binding assay with fluorescence-labeled LPS demonstrated that A-9 did not affect the binding of LPS to the cell surface receptor of the RAW264.7 cells (Nishishiro, unpublished data), eliminating the possibility that the inhibitory effect of A-9 was due to the inhibition of LPS binding to its receptor. It has been reported that PLA2α (15) and COX-2 (16) were activated by S-nitrosylation caused by iNOS. Therefore, it remains to be investigated whether A-9 inhibits the nitrosylation of PLA2α and COX-2 proteins leading to the lower production of PGE2.
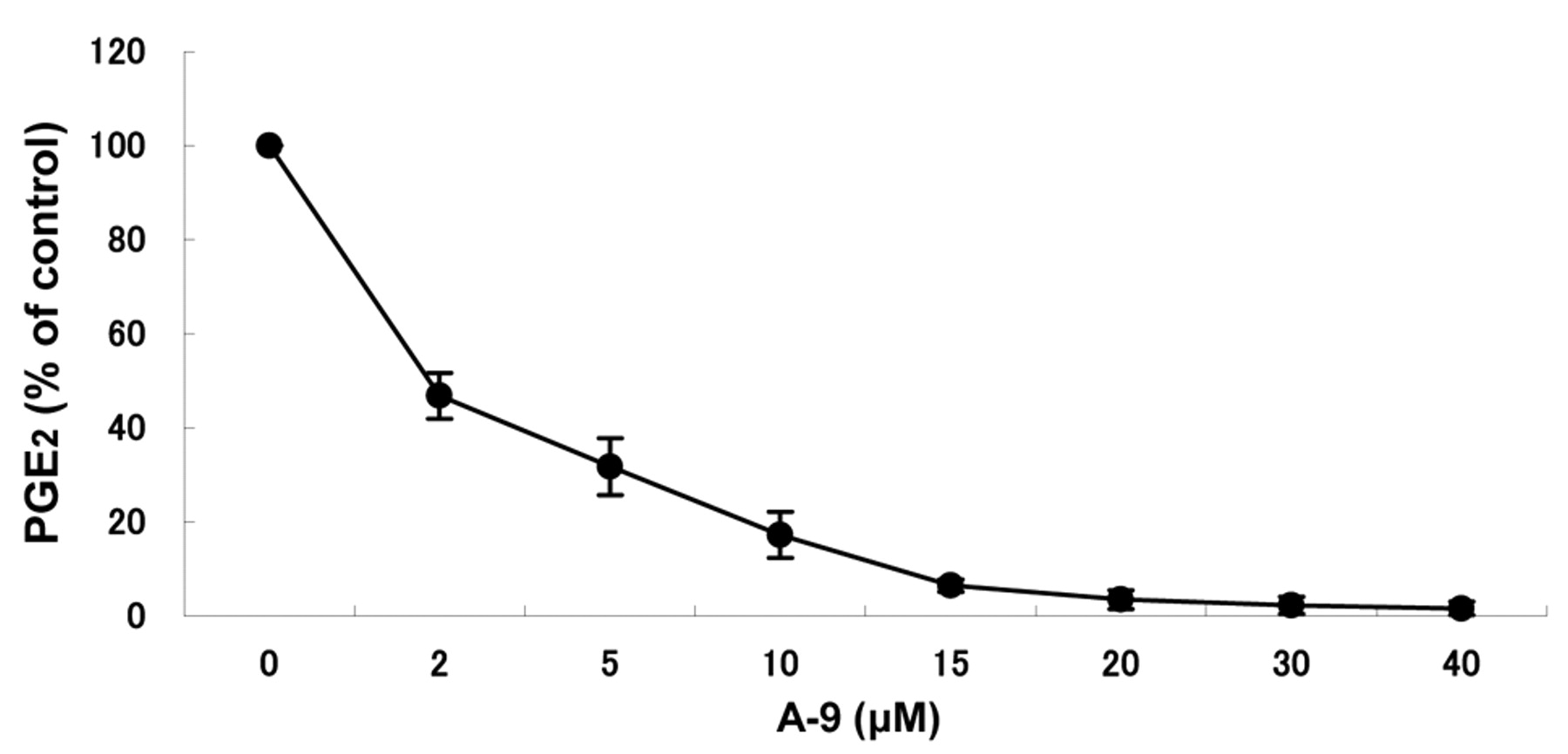
Dose response of A-9 in the inhibition of PGE2 production by LPS-stimulated RAW264.7 cells. RAW264.7 cells were incubated for 24 hours without (control), or with the indicated concentrations of diethyl 2-chloroazulene-1,3-dicarboxylate (A-9), and the extracellular concentration of PGE2 was then measured. Each value represents the mean and SD from three independent experiments.

Effect of diethyl 2-chloroazulene-1,3-dicarboxylate (A-9) on the intracellular concentration of PLA2, iNOS and COX-2. RAW264.7 cells were incubated for 24 hours with the indicated concentrations of (A-9) in the absence (-) or presence (+) of 0.1 μg/ml LPS, and PLA2 (A), COX-2 (B) or iNOS (C) protein levels were determined by western blot analysis. Each value represents mean and SD from three independent experiments.
We have previously reported that a lower concentration (16.3 μM) of A-9 slightly reduced the viable cell number of RAW264.7 cells with or without LPS stimulation (as shown by the dose-response curve in Figure 2) (2). This slight reduction of viability might overestimate the extent of the inhibition of NO production. However, such a minor growth inhibitory effect of A-9 at higher concentrations does not affect the inhibition of PGE2 production observed at much lower concentration. Further study is required to elucidate the mechanism of the possible anti-inflammatory potency of A-9.
Footnotes
- Received May 19, 2008.
- Revision received November 20, 2008.
- Accepted December 1, 2008.
- Copyright© 2009 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Cytotoxic Activity of Benzo[b]cyclohept[e][1,4]oxazines
- Anti-UV Activity of Newly-synthesized Water-soluble Azulenes
- Effect of Sasa senanensis Rehder Extract on NO and PGE2 Production by Activated Mouse Macrophage-like RAW264.7 Cells
- Inhibition of NO Production in LPS-stimulated Mouse Macrophage-like Cells by Benzo[b]cyclohept[e] [1,4]oxazine and 2-Aminotropone Derivatives
- Tumor-specific Cytotoxicity and Type of Cell Death Induced by Benzo[b]cyclohept[e][1,4]oxazine and 2-Aminotropone Derivatives