


Abstract
Resveratrol, a grape polyphenol, is thought to have anti-inflammatory, cardioprotective, and cancer preventive properties. However, the mechanisms by which resveratrol might produce these effects are not clearly defined. A study was performed on whether resveratrol could prevent tumor cells from adhering to endothelial cells, which is an essential step during tumor metastasis. Phorbol 12-myristate 13-acetate (PMA) induced human fibrosarcoma HT1080 cells to adhere to endothelial ECV304 cells. Resveratrol inhibited PMA-induced HT1080 cells adhesion in a dose-dependent manner. To further study the mechanisms of this resveratrol-mediated blockade of tumor cell adhesion, the expression of the cell adhesion molecules intercellular adhesion molecule-1 (ICAM-1), vascular cell adhesion molecule-1 (VCAM-1) and E-selectin were examined. PMA induced ICAM-1 expression in HT1080 cells. In contrast, the expression of VCAM-1 and E-selectin were not altered by PMA treatment. The increase in tumor cell adhesion to endothelial cells following PMA treatment was partially inhibited by ICAM-1 siRNA or neutralizing antibodies. Resveratrol reduced the PMA-induced ICAM-1 expression in HT1080 cells as determined by RT-PCR, flow cytometry and ELISA. As the induction of ICAM-1 requires activation of the transcription factor NF-κB, the effects of resveratrol on the activation of this factor in HT1080 cells was also investigated. Resveratrol inhibited the PMA-induced NF-κB activation and NF-κB-dependent luciferase activity. These results suggest that resveratrol may exert an anti-metastatic effect by inhibiting NF-κB activation and ICAM-1 expression, leading to suppression of tumor cell adhesion to endothelial cells.
Cancer metastasis is a multi-step process beginning with the invasion of malignant cells from the primary tumor into the vascular or lymphatic circulation. Loss of cell adhesive molecules induces the disassembly of free cancer cells from the primary tumor masses, disseminating them to distant sites via blood vessels and lymphatics, and eventually elicits the transition from benign tumors to metastatic tumors (1, 2). The circulating tumor cells interact with the endothelial cells of blood vessels, and this is considered a preliminary event during tumor metastasis (3). The adhesive interactions between tumor cells and the capillary wall do not result from random adhesion of cancer cells but rather is a selection process favoring certain tumor cells with specific properties (4).
As with leukocytes, attachment of cancer cells to the vascular endothelium seems to be initiated by the interaction between sialyl Lewis X, a surface carbohydrate expressed on several cell types including many tumor cells (5). Various adhesive molecules, including intracellular adhesion molecule-1 (ICAM-1), vascular cell adhesion molecule-1 (VCAM-1), and E-selectin have been identified as being responsible for the endothelial adhesion of cancer cells. ICAM-1 and VCAM-1 are 95 and 110 kDa proteins, respectively, and both belong to the immunoglobulin superfamily. Besides endothelial cells, ICAM-1 is also expressed by monocytes, B and T cells, keratinocytes, chondrocytes, and epithelial cells. VCAM-1 is expressed in endothelial cells, monocytes, dendritic cells, myoblast, and bone marrow fibroblasts. The 115 kDa protein E-selectin is known to be expressed exclusively on endothelial cells (6). Some of these adhesive molecules are constitutively expressed while others are induced by many different environmental signals and distributed in a tissue-specific manner as well. Clinical studies show elevated serum levels of soluble ICAM-1 and VCAM-1 in patients with cancer and these levels correlate with tumor progression (7). Resveratrol (3,5,40-trihydroxystilbene), which was first isolated from the roots of white hellebore (Veratrum glandiflorum O. Loes), has been shown to inhibit or retard the growth of various cancer cells in culture and implanted tumors in vivo (8). Resveratrol was reported to target many components of intracellular signaling pathways including pro-inflammatory mediators, regulators of cell survival and apoptosis, and tumor angiogenic and metastatic switches by modulating a distinct set of upstream kinases, transcription factors and their regulators (9). However, the effect of resveratrol on tumor cell adhesion to endothelial cells is less well understood.
In the present study, the effects of resveratrol on the adhesion of HT1080 fibrosarcoma cells to endothelial ECV304 cells were examined. It was found that resveratrol inhibits the PMA-induced adhesion of HT1080 cells to ECV304 cells by inhibiting NF-κB activation and expression of ICAM-1.
Materials and Methods
Cell lines and chemicals. Human endothelial ECV304 cells and human fibrosarcoma HT1080 cells were obtained from the American Type Culture Collection (VA, USA) and cultured in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum (FBS) and 1% penicillin-streptomycin at 37°C in 5% CO2. PMA and resveratrol were obtained from Sigma Chemical Co. (MO, USA).
Adhesion assay. HT1080 cells, grown in 6-well plates, were treated at 37°C with PMA for 4 hr after pretreatment with resveratrol for 1 h and then washed twice with phosphate-buffered saline (PBS). HT1080 cells were labeled for 30 min at 37°C with 10 ng/mL calcein-AM (Molecular Probes, The Netherlands) and then 2.0×105 of the labeled cells were added to the ECV304 monolayers, grown in 96 well plates, to a final volume of 100 μL and incubated in a CO2 incubator for 1 hr. Non-adherent cells were removed from the plate by gentle washing twice with PBS, and the fluorescent intensity of the monolayer-associated HT1080 cells was quantitated using a fluorescent plate reader. Calcein-AM is a cell-permeable, non-fluorescent and hydrophobic compound, which is rapidly hydrolyzed by cytoplasmic esterases, releasing the membrane-impermeable hydrophilic and intensely fluorescent calcein (10). The fluorescence was excited at 488 nm using an argon laser, and the evoked emission was filtered with a 515 nm long pass filter using a fluorometer (Fluoroskan; Labsystems, Finland). The adhered cells were observed and photographed under a fluorescence microscope (Olympus 1×50, Shibuya, Japan). To determine the role of ICAM-1 in the adhesion of HT1080 cells to ECV304 cells, HT1080 cells were pre-incubated with 10 μg/ml of anti-human ICAM-1 (CD54) antibody (BD Biosciences Pharmingen, CA, USA) for 1 h, or transiently transfected with the ICAM-1 siRNA oligonucleotides prior to adhesion assay.
RT-PCR. Total RNA was extracted from the HT1080 cells using TRIzol reagent (Invitrogen, CA, USA). One microgram of the total RNA was used for first strand cDNA synthesis using random primers and Superscript reverse transcriptase (Invitrogen) according to the manufacturer's protocols. The following primers with their predicted product size were used for amplification: GAPDH (836bp) 5′-TTGTTGCCATCAATGACCCC-3′ (forward) and 5′-TGA CAAAGT GGTCGTTGAGG-3′ (reverse), ICAM-1(289bp) 5′-CGACTGGA CGAGAGGGATTG-3′ (forward) and 5′-TTATGACTGCGGCTGC TACC-3′ (reverse), VCAM-1(336bp) 5′-GCAAGGT TCCTAGC GTGTAC-3′ (forward) and 5′-GGCTCAAGCTGTCAT ATTCAC-3′ (reverse), E-selectin (537bp) 5′-AACTTCCATGAG GCCAAACG-3′ (forward) and 5′-TTGTCGTTGCCAGTGTTCAG-3′ (reverse). The PCR conditions were as follows: denaturation at 94°C for 30 seconds, annealing at 58°C for 30 seconds and extension at 72°C for 30 seconds. The PCR products were subjected to 1% agarose gel electrophoresis.
Transfection of HT1080 cells with siRNA targeted ICAM-1. The HT1080 cells were transiently transfected with the siRNA oligonucleotides using Lipofectamine RNAiMAX (Invitrogen) according to the manufacturer's instructions. Starved HT1080 cells were transfected with 20 nM siRNAs for 5 h and stabilized for 48 hr in DMEM supplemented with 10% FBS and 1% penicillin-streptomycin at 37°C in 5% CO2 for further assays. The siRNA oligonucleotide sequences for ICAM-1 are as follows: 5′-CGGCUGGAGCUGUUUGAGATT-3′ and 5′-GGAGUGAUUUUU CUAUCGGTT-3′ (Invitrogen).
Flow cytometry. ICAM-1 expression in HT1080 cells was measured using flow cytometry. The HT1080 cells were incubated in the absence or presence of PMA and/or resveratrol. After incubation, 1×106 cells/ml were stained with PE-conjugated mouse anti-human ICAM-1 antibody (BD Biosciences PharMingen) for 20 min on ice. The samples were washed three times with PBS and then analyzed using flow cytometry (Becton Dickinson Labware, NJ, USA). The sample data were acquired on a FACS Calibur cell sorter (Becton Dickinson) and analyzed with CellQuest software (Becton Dickinson).
ELISA for ICAM-1. The concentration of ICAM-1 in the culture supernatants was measured using a commercially available ELISA kit (R&D system, MN, USA). Briefly, the samples were incubated in microtiter plates precoated with a monoclonal antibody specific to ICAM-1. After incubation at room temperature for 2 hr and washing, the substrate solution was added. Color development was stopped after 30 min at room temperature, and the color intensity was read at 450 nm on a spectrophotometer microplate reader (μQuant; Bio-Tek Instrument, Inc., VT, USA).
Extraction of nuclear proteins. Eighty to ninety percent confluent HT1080 cells were incubated in a medium containing 1% FBS in the presence or absence of PMA and/or resveratrol. The cells were resuspended in 500 μl of cold buffer A (50 mM Tris [pH 7.4], 150 nM NaCl, 0.2 mM EDTA, 3% [v/v] glycerol, and 1.5 mM MgCl2) on ice. Subsequently, cells were lysed with 500 μl of buffer B (identical to buffer A but containing 0.05% Nonidet P-40, Sigma). The cell lysate was gently layered onto an equal volume cushion of buffer C (10 mM Tris [pH 7.4], 25% [v/v] glycerol, and 1.5 mM MgCl2) and centrifuged for 5 min at 200 ×g. The white nuclear pellet was resuspended in 75 μl of a cold high-salt lysis buffer (20 mM Hepes [pH 7.9], 400 mM NaCl, 1 mM EDTA, 1 mM dithiothreitol, and 1 mM phenylmethylsulfonyl fluoride). This suspension was agitated for 30 min and microcentrifuged for 15 min at 4°C. The resulting supernatant was stored in aliquots at -80°C. The protein was quantified spectrophotometrically using the BCA assay (Pierce, IL, USA) with bovine serum albumin as the standard.
Electrophoretic mobility shift assay (EMSA). EMSA was performed with a gel shift assay system (Promega, WI, USA). Briefly, oligonucleotides containing the consensus sequences for NF-κB (5′-AGT TGA GGG GAC TTT CCC AGG-3′) were end-labeled with [γ-32P] adenosine triphosphate (3000 μCi/mmol; Amersham Pharmacia Biotech, UK) using T4 polynucleotide kinase. The labeled oligonucleotides were then purified in Microspin G-25 columns (Sigma) and used as probes for EMSA. The nuclear extract proteins (10 μg) were pre-incubated with the binding buffer (10 mM Tris-HCl [pH 7.5], 50 mM NaCl, 0.5 mM EDTA, 1 mM MgCl2, 0.5 mM dithiothreitol, 4 % [v/v] glycerol, and 0.05 mg/mL poly[deoxyinosine-deoxycytosine]) for 5 min and then incubated with the labeled probe for 15 min at 37°C. Each sample underwent electrophoresis in a 5 % non-denaturing polyacrylamide gel in 0.5 × Tris-borate-EDTA buffer (pH 7.4) at 150 V for 4 h. The gel was dried and subjected to autoradiography. In the competition study, a 50-fold excess of unlabeled oligonucleotide was included in the reaction mixture along with the radiolabeled probe.
NF-κB-reporter assay. The NF-κB-reporter construct was purchased form Clontech (CA, USA). Once the cells had reached 60-70% confluence, they were washed with DMEM and incubated with DMEM without serum and antibiotics for 18 h. The cells were then transfected with 1 μg NF-κB-reporter containing the pGL3 vector using Lipofectamine 2000 (Invitrogen). In order to determine the inhibitory effects of resveratrol on NF-κB by PMA, the transfected cells were pretreated with 0-50 μM resveratrol for 1 h and incubated with 5 nM PMA for 4 h. After incubation, the cells were lysed and the luciferase activity was measured using a luminometer.

Effect of PMA on the adhesion of HT1080 cells to ECV304 cells. HT1080 cells were treated with 0-50 nM PMA for 4 h and then labeled with 10 ng/mL calcein-AM. The labeled HT1080 cells were seeded onto the ECV304 cell monolayers and co-cultured for 1 h. After removing the nonadherent cells, the remaining adherent cells were visualized as bright dots by fluorescent microscope (A) and quantitated using a fluorometer (B). Data are expressed as the mean±SEM of averages from at least three experiments.
Statistical analysis. Data are expressed as the mean±SEM of averages from at least three experiments. ANOVA was used to assess the statistical significance of the differences, and a value of p<0.05 was considered statistically significant.
Results
Effect of resveratrol on the adhesion of PMA-treated fibrosarcoma cells to endothelial cell monolayers. In order to quantify the adhesion of the fibrosarcoma HT1080 cells to endothelial ECV304 cells, HT1080 cells were treated with different pharmacological doses of PMA (0-50 nM) and then labeled with calcein-AM. The labeled HT1080 cells were seeded onto the ECV304 cell monolayers, and co-cultured for 1 h. After removing the non-adherent cells, the remaining adherent cells were evaluated by the quantification of fluorescent calcein. As shown in Figure 1A and B, PMA treatment of HT1080 cells induced increased adherence to endothelial ECV304 cells in a dose-dependent manner. However, PMA-treated ECV304 cells, independent of the concentration and similar to untreated cells, did not adhere to HT1080 cells (data not shown). To study the effect of resveratrol on the adhesion of HT1080 cells to ECV 304 cells, HT1080 cells were pre-treated with different concentrations of resveratrol in the presence or absence of PMA. As shown in Figure 2A and B, resveratrol inhibited the adhesion of HT1080 cells to ECV304 cells in a dose-dependent manner. Cell viability at the concentrations of resveratrol used in these experiments was not affected.
Role of ICAM-1 in PMA-induced HT1080 cell adhesion to ECV304 cells. To determine which adhesion molecules are involved in this cellular adhesion event, the expression status of various adhesion molecules and their possible association with the adhesion capability of the cells was analyzed. Cells were treated with PMA (50 nM) for 4 h and then RT-PCR was performed for the expression of the adhesion molecules ICAM-1, VCAM-1 and E-selectin. Whereas the ECV304 cells expressed all the adhesion molecules examined, HT1080 cells expressed only ICAM-1, but not VCAM-1 or E-selectin. Furthermore, the expression of ICAM-1 in HT1080 cells was significantly increased following PMA treatment (Figure 3A). To examine the specific role of ICAM-1 in the adhesion of HT1080 cells, anti-ICAM-1 antibodies and ICAM siRNA were used. Pre-treatment of HT1080 cells with anti-ICAM-1 antibodies prior to the assessment of binding dose-dependently inhibited the cells from binding to the ECV304 cells. However, non-specific IgG did not alter the adhesiveness of the cells. Additionally, siRNA targeting ICAM-1 also inhibited HT1080 cell adhesion (Figure 3B), suggesting that ICAM-1 is important for the adhesion of HT1080 cells to ECV304 endothelial cell monolayers.
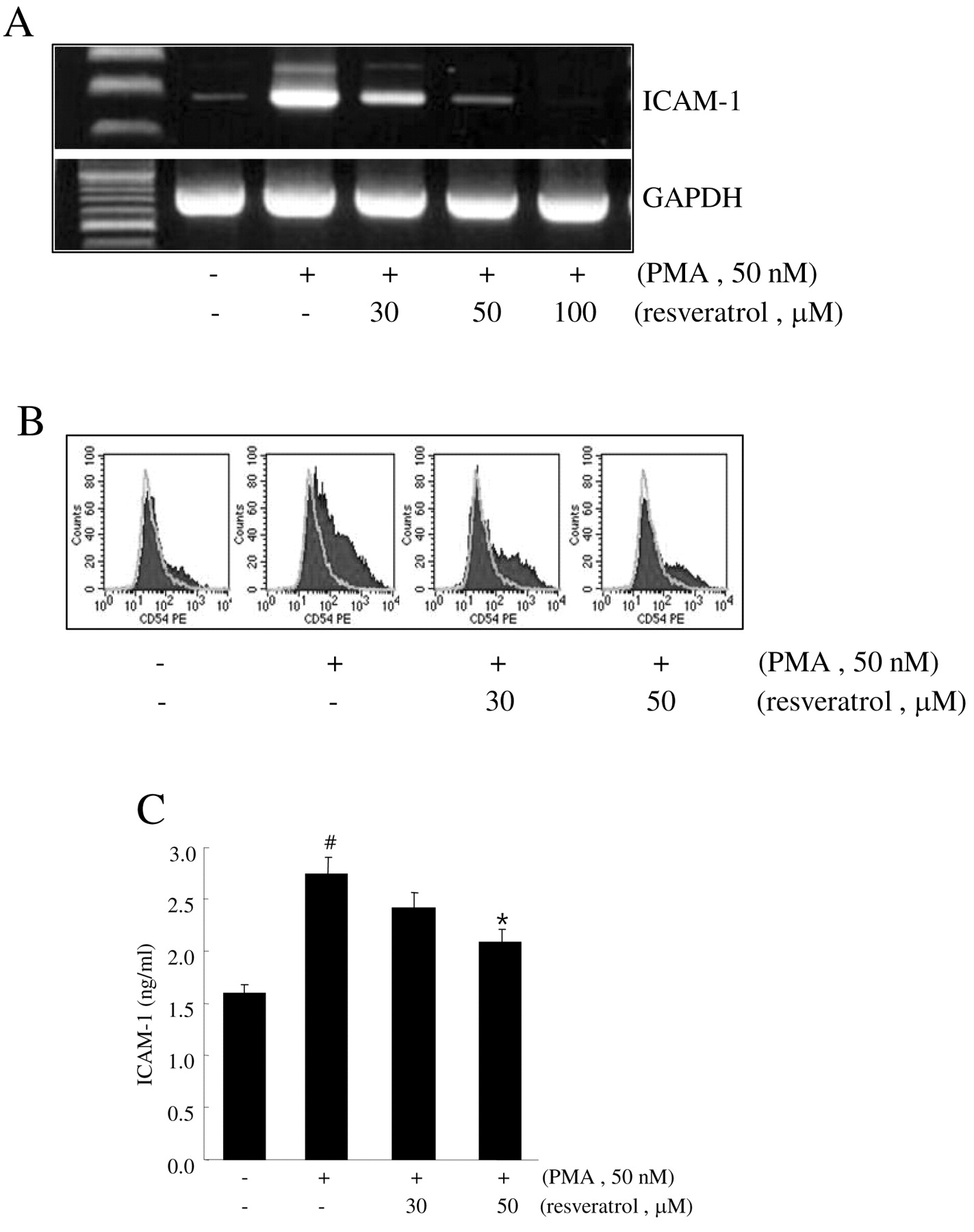
Effect of resveratrol on the expression of ICAM-1 in HT1080 cells. To determine whether ICAM-1 is an important target for the resveratrol-mediated blockade of tumor cell adhesion to endothelial cells, the effect of resveratrol on the expression of ICAM-1 was examined. ICAM-1 expression was upregulated in the presence of PMA compared to ICAM-1 levels in control cells, and the introduction of resveratrol attenuated this effect as determined by RT-PCR and flow cytometry (Figure 4A and B). Additionally, the cell culture supernatant of PMA-treated HT1080 cells showed an increase in ICAM-1 concentration, and this increase was also abrogated by resveratrol treatment in a dose-dependent manner (Figure 4C). These results suggest that the ability of resveratrol to inhibit PMA induction of ICAM-1 could prevent tumor cell adhesion to endothelial cell monolayers.

Effect of resveratrol on the PMA-induced HT1080 cell adhesion to ECV304 cells. HT1080 cells pretreated with the indicated concentrations of resveratrol for 1 hr were treated with 50 nM PMA for 4 h and then labeled with 10 ng/mL calcein-AM. The labeled HT1080 cells were seeded onto the ECV304 cell monolayers and co-cultured for 1 h. After removing the non-adherent cells, the remaining adherent cells were visualized as bright dots by fluorescent microscope (A) and quantitated using a fluorometer (B). Data are expressed as the mean±SEM of averages from at least three experiments.
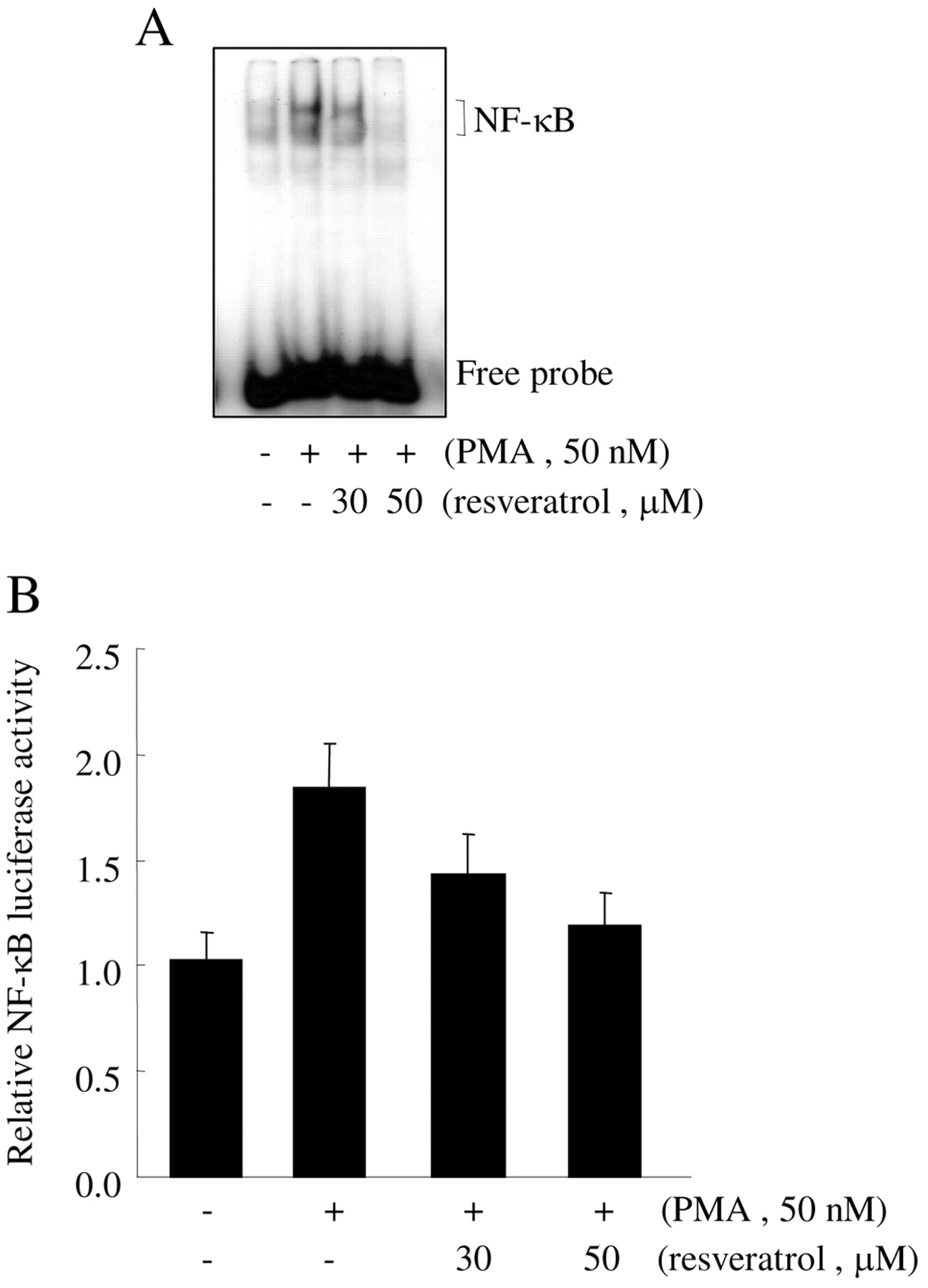
Effect of resveratrol on the PMA-induced NF-κB activation. While the expression of ICAM-1 requires the transcription factor NF-κB, it is not known whether resveratrol inhibits the expression of ICAM-1 through suppression of NF-κB activation in HT1080 cells. The NF-κB activities of the cells treated with resveratrol before PMA treatment were examined. As shown in Figure 5A, PMA treatment caused a remarkable increase in the amount of NF-κB that could form a complex with a radiolabeled oligonucleotide probe as determined by EMSA. However, the binding of NF-κB, from the cells pre-treated with resveratrol, to the DNA probe was decreased in a dose-dependent manner. The effect of resveratrol on NF-κB-dependent transcriptional activity was examined to gain further insight into the mechanism of resveratrol-mediated regulation of NF-κB. Consistent with the EMSA results, resveratrol treatment decreased the PMA-induced NF-κB-dependent transcriptional activity in a dose-dependent manner (Figure 5B). This suggests that resveratrol decreased the PMA-induced expression of ICAM-1 through the suppression of NF-κB activation.

Involvement of ICAM-1 in PMA-induced HT1080 cell adhesion to ECV304 cells. HT1080 cells and ECV304 cells were treated with the indicated concentrations of PMA for 4 h and then total RNA was extracted. ICAM-1, VCAM-1 and E-selectin mRNA expression was determined by RT-PCR. GAPDH was used as a normalization control (A). HT1080 cells treated with 50 nM PMA for 4 h were incubated with 10 μg/mL of anti-human ICAM-1 antibodies or nonspecific IgG for 1 h and then labeled with 10 ng/mL calcein-AM, or HT1080 cells transfected with 20 nM ICAM-1 siRNA were incubated with 50 nM PMA for 4 h and then labeled with 10 ng/mL calcein-AM. The labeled HT1080 cells were seeded onto the ECV304 cell monolayers and co-cultured for 1 h. After removing the non-adherent cells, the remaining adherent cells were quantitated using a fluorometer (B). Data are expressed as the mean±SEM of averages from at least three experiments. #p<0.05 versus none (without PMA); *p<0.05 versus PMA alone.
Discussion
The results demonstrated that resveratrol decreased the attachment of fibrosarcoma HT1080 cells to endothelial ECV304 cells. Due to the apparent importance of cell adhesion in cancer progression and metastasis, it is considered that blocking the interaction of cancer cells with endothelial cells might be an important therapeutic target.
The anti-cancer property of resveratrol has been extensively studied in vitro and in vivo. Interest in resveratrol as a chemopreventive agent in humans has intensified for several reasons. First, preclinical studies suggested that resveratrol was effective in blocking tumor initiation, promotion and progression. According to Jang et al. (11), topically applied resveratrol significantly reduced 7,12-dimethylbenz[α]anthracene-initiated and 12-O-teradecanoyl-phorbol-13-acetate-promoted skin tumors in female CD-1 mice. Subsequent studies demonstrated that resveratrol exhibited strong chemopreventive effects in various experimentally induced tumor models (12). Several recent studies reported that intratumoral, peritumoral or intraperitoneal administration of resveratrol significantly arrested tumor growth in vivo and induced apoptosis in xenografted tumors in athymic nude mice (13, 14). Second, the mechanistic findings with resveratrol support this phytoalexin as a chemopreventive and chemotherapeutic agent. Resveratrol was reported to prevent oxidative damage of target cell DNA and to modulate various intracellular signal transduction pathways, which often go awry during the course of carcinogenesis. Blockade of angiogenic and metastatic processes during tumor progression and alleviation of chemotherapy resistance indicate the chemotherapeutic potential of resveratrol (12). Third, resveratrol consumption is associated with few adverse events, and resveratrol is readily available at a low cost.
The process of tumor cell adhesion and metastasis involves the interaction between endothelial cells of the target organ and blood borne tumor cells, and this is an important event for implementing the subsequent steps of invasion and survival of tumor cells. In the present study, the effects of resveratrol on the adhesion of fibrosarcoma HT1080 cells to endothelial ECV304 cells were investigated. The results presented here clearly demonstrate that resveratrol blocked the PMA-mediated attachment of HT1080 cells to EVC304 cells (Figures 1 and 2). A number of cell adhesion molecules have been implicated in the interaction of cancer cells with vascular endothelial cells including ICAM-1, VCAM and E-selectin. The specific adhesion molecules that mediate tumor cell attachment are dependent on the type of malignancy and on the target tissue endothelium. Here it is demonstrated that tumor cells producing ICAM-1 enhance the adhesion capability of these cancer cells. The following observations suggest that the expression of ICAM-1 in HT1080 cells may contribute to their interaction with ECV304 cells: i) ICAM-1 was expressed constitutively and upregulated following PMA treatment in HT1080 cells (Figure 3A); ii) blockade of ICAM-1 with neutralizing antibodies and siRNA targeting ICAM-1 resulted in significantly decreased adherence of HT1080 cells (Figure 3B).
ICAM-1 is a transmembrane glycoprotein cell surface receptor and a member of the immunoglobulin superfamily. It can be membrane bound or soluble and participates in cell-cell and cell-matrix adhesive interactions (10). ICAM-1 has been demonstrated to influence broad aspects of tumor progression such as cellular proliferation, angiogenesis and metastasis. ICAM-1 principally mediates intercellular contacts via its interaction with CD43 (15). Interestingly, a recent study demonstrated that ICAM-1 is expressed in tumor cells as well as endothelial cells, and that cancer patients with high levels of ICAM-1 expression show poor prognosis when compared to healthy controls (16, 17). Moreover, increased ICAM-1 expression significantly correlates with stage, invasion depth, and lymph node and distant metastasis cancers, indicating that ICAM-1 plays important roles in cancer progression and metastasis. Understanding the precise regulation of ICAM-1 production, under cancerous conditions, is therefore critical for the prevention of tumor progression and metastasis.

Effect of resveratrol on the PMA-induced ICAM expression in HT1080 cells. HT1080 cells pretreated with the indicated concentrations of resveratrol for 1 h were treated with 50 nM PMA for 4 h and then total RNA was extracted. ICAM-1 and GAPDH mRNA expression was determined by RT-PCR (A). HT1080 cells pre-treated with the indicated concentrations of resveratrol for 1 h were treated with 50 nM PMA for 4 h and then stained with PE-conjugated mouse anti-human ICAM-1 antibody for 20 min. The expression of ICAM-1 was analyzed using flow cytometry (B). HT1080 cells pretreated with the indicated concentrations of resveratrol for 1 h were treated with 50 nM PMA for 24 h and then the concentration of ICAM-1 in the culture supernatants was measured by ELISA (C). Data are expressed as the mean±SEM of averages from at least three experiments. #p<0.05 versus none (without PMA); *p<0.05 versus PMA alone.
ICAM-1 gene expression is regulated by numerous cytokines and is primarily regulated at the mRNA level. The architecture of the ICAM-1 promoter is complex and is regulated by the interplay between different transcription factors such as NF-κB and AP-1 (18). Some polyphenolic compounds have been found to suppress cytokine-induced activation of NF-κB and hence the subsequent expression of ICAM-1. In addition, anti-inflammatory compounds, including antioxidants, PPAR activators and statins, have been shown to be effective in reducing ICAM-1 gene expression through the inhibition of NF-κB. Previous studies have shown that resveratrol blocks TNF-α induced activation of NF-κB and consequently inhibits NF-κB-dependent gene expression (19, 20). However, the role of resveratrol in ICAM-1 gene expression, which is mediated by NF-κB activation, remains unclear. In the present study, it has been demonstrated that resveratrol inhibits PMA-induced NF-κB activation in HT1080 cells (Figure 5). The level of NF-κB-DNA binding and NF-κB-dependent transcriptional activities in the cells pre-treated with resveratrol decreased in a dose-dependent manner. The mechanisms by which resveratrol inhibited the activation of NF-κB are cell type- and stimuli-specific.

Effect of resveratrol on the PMA-induced NF-κB activation in HT1080 cells. HT1080 cells pretreated with the indicated concentrations of resveratrol for 1 h were treated with 50 nM PMA for 4 h and then nuclear proteins were extracted. The binding of NF-κB-DNA was assayed by EMSA (A). HT1080 cells transiently transfected with 1 μg NF-κB-reporter were incubated with the indicated concentrations of resveratrol for 1 h and then treated with 50 nM PMA for 4 h. After incubation, the cells were lysed and luciferase activity was measured using a luminometer. Data are expressed as the mean±SEM of averages from at least three experiments (B).
Whereas in U937 cells, resveratrol diminished TNF-α-induced activation of NF-κB by suppressing phosphorylation and nuclear translocation of p65 without affecting IκB degradation (19), in normal human epidermal keratinocytes, the compound inhibited UVB-induced activation of NF-κB by blocking the activation of upstream IKK as well as phosphorylation and degradation of IκB (21). Topical application of resveratrol attenuated TPA-induced NF-κB activation in mouse skin (in vivo) by blocking the activation of IKK, phosphorylation of IκB and p65, nuclear translocation of p65, and the interaction of p65 with a transcriptional co-activator cyclic AMP-response element binding protein (22). Alternatively, resveratrol also exerted epigenetic control on NF-κB activation by inducing SIRT1 activation (23). It was reported that SIRT1, a nicotinamide adenosine dinucleotide-dependent histone deacetylase, interacted physically with the RelA/p65 and negated NF-κB-driven gene transcription by deacetylating RelA/p65 at lysine 310 (24).
This study found that resveratrol could block ICAM-1 expression and cell adhesion between HT1080 and ECV304 cells, which suggest that the down-regulation of ICAM-1 by resveratrol is involved in decreasing cell adhesion. However, further studies will be needed to clarify these mechanisms. Understanding the molecular mechanisms by which resveratrol inhibits NF-κB activation and ICAM-1 expression will serve as a basis for designing more effective anti-metastatic drugs.
Acknowledgements
This work was supported by a Research Grant (0720570) from the National Cancer Center and by a Medical Research Center (R13-2002-013-06001-0) from the Korea Science and Engineering Foundation.
Footnotes
- Received October 27, 2008.
- Revision received November 26, 2008.
- Accepted December 2, 2008.
- Copyright© 2009 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}