


Abstract
Background: The aim of this study was to analyze the spatial distribution and proliferation of adoptively transferred CD8+ T-lymphocytes sensitized against allogeneic tumors. Materials and Methods: Transgenic β-actin-luc mice that express luciferase were sensitized against allogeneic SL2 lymphoma. CD8+ T-lymphocytes from these mice were transferred to lymphocyte-deficient, recombination activating gene-deficient (Rag−/−) mice bearing SL2 tumors and were tracked using bioluminescence imaging. Results: Two out of six Rag−/− mice rejected their tumors. There were no apparent differences in spatial distribution and proliferative intensity of adoptively-transferred CD8+ T-lymphocytes between the two Rag−/− mice that rejected allogeneic SL2 tumors and the four Rag−/− mice that did not. Conclusion: The pattern of distribution in the mouse body and proliferative intensity of CD8+ T-lymphocytes do not seem to be decisive factors influencing allogeneic tumor rejection.
The prevailing view holds that cytotoxic T-lymphocytes kill tumor cells by a lytic mechanism requiring direct contact between T-lymphocytes and target tumor cells (1). If this lytic mechanism indeed operates in vivo, rejection of tumors by cytotoxic T-lymphocytes should always be preceded by expansion of a specific clone of CD8+ T-lymphocytes and their migration to the tumor site. Migration of CD8+ T-lymphocytes to tumors expressing human papillomavirus-encoded oncogenic protein E7 (2), and chicken ovalbumin and SV40 large T-antigen (3) has been observed using bioluminescence imaging. In our previous publication, using bioluminescence imaging, we demonstrated preferential accumulation of lymphocytes at the tumor site during re-challenge with allogeneic SL2 lymphoma cells after primary SL2 tumor has been rejected (4). Thus, bioluminescence imaging is a sensitive tool for detecting intratumoral accumulation of lymphocytes. In addition, measuring the total photon flux allows estimation of lymphocyte proliferation. In the present study, we analyzed the spatial distribution and proliferation intensity of CD8+ T-lymphocytes during challenge with primary allogeneic SL2 tumors.
Materials and Methods
Mice and tumors. Transgenic luciferase-expressing B6, FVB β-actin-luc female mice (Light Producing Transgenic Animals®, Model 10500) were purchased from Taconic (Germantown, NY, USA). These mice are described in more detail in our previous publication (4). Transgenic β-actin-luc mice at the age of 10 months were sensitized against allogeneic SL2 tumor by subcutaneous injection on the left side of the chest of 1×106 tumor cells in phosphate-buffered saline (PBS).
CD8+ T-lymphocytes isolated from spleens of transgenic β-actin-luc mice were transferred to 11-to 14-week-old lymphocyte-deficient, recombination activating gene-deficient (Rag−/−) mice, bearing SL2 tumors. Rag-deficient (Rag1−/− or Rag2−/−) mice were a kind gift from Professor Reinhard Voll, University Erlangen-Nürnberg, Germany, and were bred at a local breeding facility at the State Research Institute Centre for Innovative Medicine, Vilnius, Lithuania. Experimental research on animals was conducted according to guidelines of the Lithuanian Ethics Committee for Laboratory Animal Use (approval no. 02-24).
SL2 lymphoma cells were maintained by weekly intraperitoneal passage in syngeneic DBA/2 mice and subsequently frozen. Prior to use in experiments, SL2 cells were thawed, washed and resuspended in RPMI-1640 medium. Solid tumors in Rag−/− mice were induced by subcutaneous injection into the right side of the chest of 1×106 SL2 cells in PBS.
Transgenic β-actin-luc mice (Model 10500) are of C57BL/6 (B6), FVB genetic background (H2b and H2q haplotype, respectively), whereas SL2 tumor cells are of DBA/2 origin which carry H-2d haplotype. Thus, SL2 tumor cells are allogeneic to T-lymphocytes from transgenic β-actin-luc mice.
Isolation and transfer of CD8+ T-lymphocytes. At 15 days after implantation of SL2 tumors, transgenic β-actin-luc mice were sacrificed by cervical dislocation, spleens were removed and single-cell lymphocyte suspensions were prepared by enzymatic digestion using Spleen Dissociation Kit in combination with mechanical dissociation using GentleMACS Dissociator (both from Miltenyi Biotec GmbH, Bergisch Gladbach, Germany). CD8+ T-lymphocytes from spleen cell suspension were separated using magnetic bead-mediated negative selection using CD8a+ T-Cell Isolation Kit, LS columns, and MidiMACS separator (Miltenyi Biotec GmbH). Isolated CD8+ T-lymphocytes were transferred intravenously in PBS to SL2 tumor-bearing Rag−/− mice (3.6×106 cells per mouse) via retro-orbital injection on day 8 after implantation of tumors.
Bioluminescence imaging. Bioluminescence measurements in SL2 tumor-bearing Rag−/− mice were performed using IVIS® Spectrum equipment (Caliper/Perkin Elmer, Hopkinton, MA, USA) at 2, 4, 6, 8, 20, 14 and 18 days after adoptive transfer of CD8+ T-lymphocytes. Two-dimensional in vivo bioluminescence imaging using auto-exposure mode was performed to evaluate the pattern of distribution of adoptively transferred CD8+ T-lymphocytes in the mouse body. Three-dimensional imaging of the whole mouse body was carried out to estimate the total flux of photons per second, which is proportional to the number of living lymphocytes in vivo and thereby can be used to estimate lymphocyte proliferation and death. Data were acquired and analyzed using Living Image® software, version 4.3.1 (Caliper/Perkin Elmer). Details of bioluminescence imaging are described in our previous publication (4).
Results
Two out of six SL2 tumor-bearing Rag−/− mice rejected tumors after adoptive transfer of CD8+ T-lymphocytes from transgenic β-actin-luc mice sensitized against allogeneic SL2 cells. The other four SL2 tumor-bearing Rag−/− mice died within 2 weeks after lymphocyte transfer (within 22 days after implantation of tumors).
Overlay of luminescent and light photographic images from tumor-bearing Rag−/− mice at 2, 6, 10 and 18 days after adoptive transfer of CD8+ T-lymphocytes is shown in Figure 1. These images demonstrate that T-lymphocytes were distributed in various parts of the mouse body. No preferential accumulation of lymphocytes at the tumor site was observed.
Total flux of photons from individual SL2 tumor-bearing Rag−/− mice at different time points after the adoptive transfer of CD8+ T-lymphocytes is shown in Figure 2. The average total photon flux increased from 1.11×107 photons per second on day 2 to 1.84×109 photons per second on day 10. This means that the number of lymphocytes in the mouse body increased on average 166-foId during this 8-day period. However, the total photon flux curves of the two mice that rejected tumors did not differ until day 10 from those of the four mice that did not reject tumors. The total photon flux increased about 37-fold from day 14 to 18 in the two mice that rejected tumors, showing proliferation of lymphocytes after tumor rejection.
Discussion
The tumor rejection rate in this study seemed to be lower compared to our previous results showing rejection of allogeneic SL2 tumors in six out of nine and in six out of six Rag−/− mice after adoptive transfer of sensitized spleen cells or T-lymphocytes, respectively (4). One explanation for the difference in tumor rejection rates between our previous and current results is that CD4+ lymphocytes are necessary for the antitumor effect (5, 6). Another possible explanation might be due to the fact that in this study middle-aged mice (10 months old) were used, whereas in the previous study, young mice (7-9 weeks old) were used as lymphocyte donors. It has been shown that the antitumor function of CD8+ T-cells critically declines with age (7).
Whatever the cause of this relatively low rejection rate, the model described here allows comparison of the distribution of CD8+ T-lymphocytes between mice that reject and mice that do not reject allogeneic tumors.
In this study, allogeneic tumors were rejected in two mice with no preferential intratumoral accumulation of CD8+ T-lymphocytes and with lymphocyte proliferation similar to that seen in the four mice that did not reject tumors. This observation is somewhat in contrast to the reports of Kim et al. (2), Charo et al. (3) and to our previous publication (4). The differences may be explained by the fact Kim et al. (2) and Charo et al. (3) used foreign antigens to increase tumor immunogenicity. In our previous publication (4), we described accumulation of T-lymphocytes in secondary allogeneic tumors after primary tumors had been rejected. It can be hypothesized that in all these cases, more specific T-lymphocytes existed with higher capacities to accumulate within tumors.
Our current observation suggests that intratumoral accumulation and expansion of CD8+ T-lymphocytes may not be decisive for rejection of allogeneic tumors. Direct contact between a sufficient number of specific CD8+ T-lymphocytes and tumor cells would be necessary if tumor cells were killed by a cytotoxic mechanism. Thus, doubts can be raised whether direct cytotoxicity is the most important mechanism in the control of allogeneic tumor by CD8+ T-lymphocytes. Our observation adds to the evidence from publications by other authors, suggesting existence of alternative mechanisms of tumor growth control by CD8+ T-lymphocytes. Cole and Ostrand-Rosenberg described rejection of allogeneic tumor by CD8+ T-lymphocytes that were not lytic for the tumor (8). Karan et al. demonstrated that decreased cytotoxic T-cell activity in vitro was associated with augmented antitumor immunity in a mouse model (9). Elucidation of alternative mechanisms of tumor growth control by CD8+ T-lymphocytes might have important implications for cancer immunotherapy.

Overlay of luminescent and light photographic images from three female (left column) and three male (right column) SL2 tumor-bearing recombination-activating gene-deficient (Rag−/−) mice at 2, 6, 10 and 18 days after adoptive transfer of 3.6×106 CD8+ T-lymphocytes isolated from the spleen of transgenic β-actin-luc mice sensitized against SL2 cells. Tumors were induced by injecting 1×106 SL2 cells into the right side of the chest 8 days prior to adoptive transfer of lymphocytes.
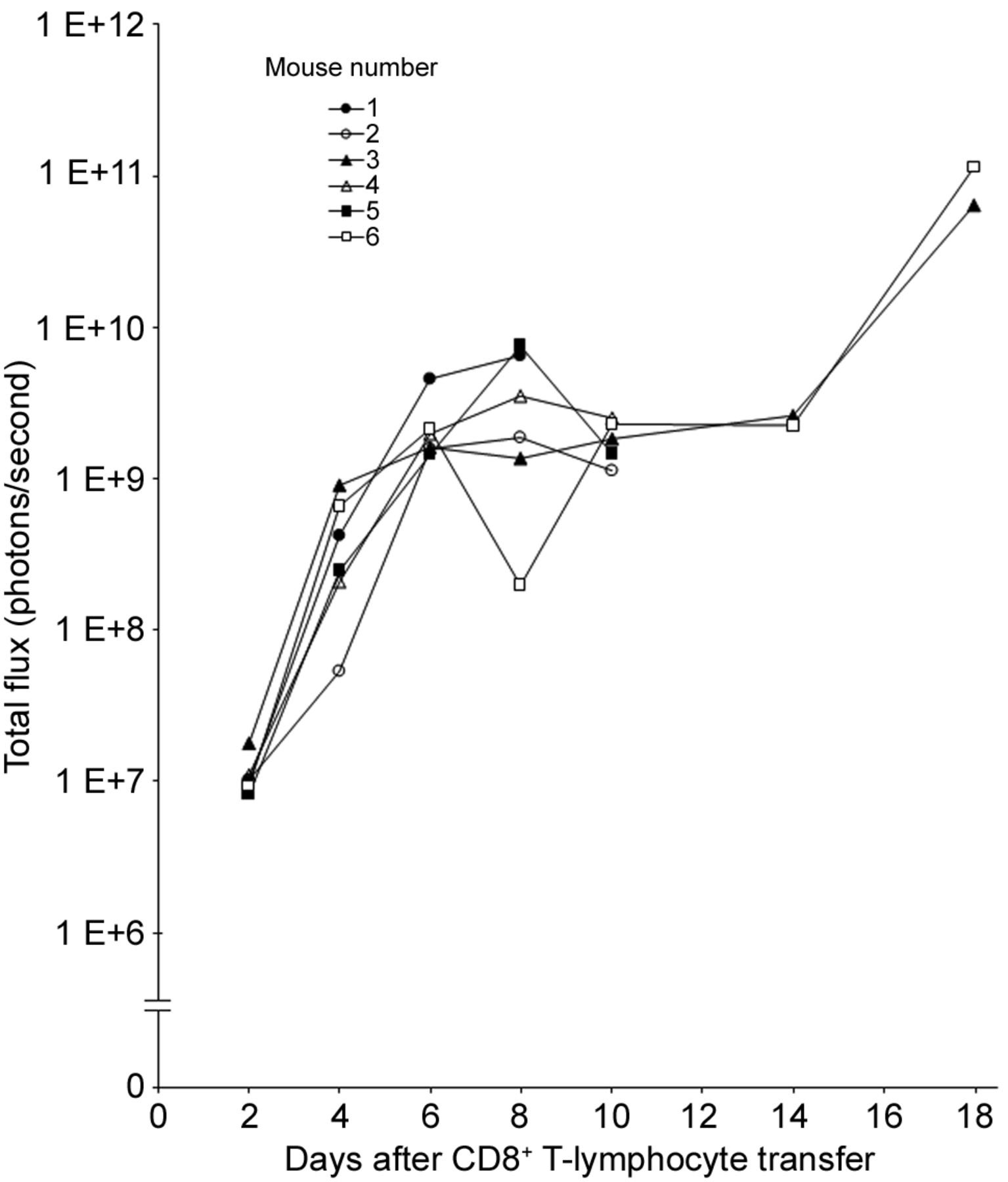
Total flux of photons from individual SL2 tumor-bearing recombination activating gene-deficient (Rag−/−) mice at different time points after adoptive transfer of 3.6×106 CD8+ T-lymphocytes isolated from the spleen of transgenic β-actin-luc mice sensitized against SL2 cells. Tumors were induced by injecting 1×106 SL2 cells into the right side of the chest 8 days prior to adoptive transfer of lymphocytes. Mouse numbers correspond to those in Figure 1.
Acknowledgements
This research was funded by a Grant (no. MIP-046/2013) from the Research Council of Lithuania.
- Received June 19, 2018.
- Revision received June 28, 2018.
- Accepted June 29, 2018.
- Copyright© 2018, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved
In this issue





{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.