


Abstract
Background/Aim: Mulberry (Morus alba L.) leaves (ML) contain many functional components, such as 1-deoxynojirimycin, flavonoids (rutin, quercetin, kaempferol). It is well known that 1-deoxynojirimycin functions to suppress increases in blood glucose level by α-glucosidase inhibitory activity. Thus, the molecular mechanism underlying the protective and therapeutic effects of ML supplementation was investigated on a mouse model of high-calorie diet (Western diet: WD)-induced hepatic steatosis (HS). Materials and Methods: The C57BL/6J mouse was used for the HS model. The mice were divided into three groups: control (normal diet: ND), WD, and WD + 1% ML groups. The WD group was fed a high-calorie (high carbohydrate and high fat) diet for 12 weeks to develop HS. At week 12, all mice were sacrificed, blood was collected for biochemical tests, and the liver was obtained for histological examination and RNA sequencing (RNA-Seq). Results: Liver weight, plasma triglycerides (TG), alanine aminotransferase (ALT), and alanine aminotransferase (AST) levels of both ML groups were significantly lower than those of the WD group. On histological examination of the liver, the area of fatty deposits was found to be suppressed by ML administration. In the gene expression analysis of the liver of WD- versus ML-fed mice by RNA-Seq, 722/45,706 genes exhibited a significant change in expression (corrected p-value<0.05). Gene network analysis of these genes showed that genes related to liver inflammation were inactivated and those related to regeneration of liver were activated in the ML group. Conclusion: ML functions to suppress HS in WD-fed mice and regulates genes related to inflammation and regeneration of liver cells.
In the modern society, opportunities to eat a high-calorie diet rich in fat and carbohydrates are greatly increasing. As a result of such lifestyle, visceral fat can accumulate, leading to metabolic syndrome and non-alcoholic fatty liver disease (NAFLD) (1, 2). Because diabetes and fatty liver disease exhibit poor subjective symptoms, treatment can be delayed, and it is not unusual for these diseases to lead to serious life-threatening situations. For example, when fatty liver disease progresses, it turns into non-alcoholic steatohepatitis (NASH); the liver becomes fibrotic and the risk of liver cancer increases (3). Regarding diseases caused by metabolic syndrome, emphasis is placed on improvement of lifestyle, such as nutritional therapy from the viewpoint of prevention.
Mulberry trees grow naturally in Southeast Asia, including Japan, the Korean Peninsula, and China, and more than 10 extant species have been confirmed thus far (4). From ancient times, leaves, root skin, branches, fruits, and so on have been used as folk medicine, and in China, the dried mulberry root skin has been used as a traditional Chinese medicine for its anti-inflammatory, anti-tussive, and other curative effects (5).
In Japan, mulberry trees have been cultivated as a silkworm plant for silk production, but in recent years the use of its leaves as health food has been attracting attention because of the decline in the silk industry. The reported components of mulberry leaf (ML) include 1-deoxynojirimycin, flavonoids (rutin, quercetin, kaempferol, astragalin) and sterols (ergosterol, campesterol, β-sitosterol). In particular, 1-deoxynojirimycin inhibits α-glycosidase and is reported to inhibit the rise in blood glucose levels (6). Therefore, mulberry leaves are currently being popularized as food/supplement products in many different forms, including powder, tablet, capsule, and liquid, to prevent diabetes.
Other benefits include anti-cholesterol, anti-hypertensive, anti-obesity, and liver protective effects. In addition, the following anti-oxidative effects have also been reported: scavenging of 1,1-diphenyl-2-picrylhydrazyl (DPPH) radicals and inhibition of the oxidation of human low-density lipoprotein (LDL) (7, 8).
A pathological model of NASH caused by metabolic syndrome was developed by exposing mice long-term to a high-fat, high-carbohydrate diet (Western diet, WD). Furthermore, by administering mulberry leaves to this mouse model and examining the gene expression of their liver, the NASH-preventive effect of mulberry leaves was determined.
Materials and Methods
Animals. Six-week-old C57BL/6 male mice were purchased from Charles River Japan, Inc. (Kanagawa, Japan). They were housed in individual cages under conventional conditions with a 12-h light–dark cycle, 23°C±1°C temperature and 55%±15% humidity.
After 1 week of acclimation, the mice were divided into three groups (n=8 per group): (i) normal diet (ND), (ii) WD, and (iii) WD in 1% ML (ML). The WD, which was used to induce hepatic steatosis (HS) in the mice, comprised F2WTD (sucrose 34%, salt-free butter 20%, 417 kcal), whereas the ND comprised MF (359 kcal) purchased from the Oriental Yeast Co., Ltd. (Tokyo, Japan). ML powder (Lot 1604) was obtained from Health Age Co., Ltd. (Tokyo, Japan) and was added to the F2WTD at 1%.
This study conformed to the Guiding Principles for the Care and Use of Experimental Animals of Hokkaido Pharmaceutical University (published 1998, revised in 2001 and 2007). The protocol approval number is H27-008.
Plasma biochemical examinations and liver histological analysis. At week 12, all mice were treated under isoflurane anesthesia and blood and liver samples were obtained. Blood samples were collected in an EDTA-containing plasma separator tube (PST) (BD Japan Co., Ltd., Tokyo, Japan) from the heart of the mice. Each plasma sample was obtained from whole blood, separated by centrifugation at 12,000 rpm/min for 10 min and stored at −80°C. The levels of triglyceride (TG), total cholesterol (T-Cho), glucose (GLU), aspartate aminotransferase (AST), and alanine aminotransferase (ALT) in plasma were determined using a DRI-CHEM 4000 device (Fujifilm Medical Co., Ltd., Tokyo, Japan).
Each liver sample was fixed in 10% neutral-buffered formalin, embedded in paraffin, and cut into thin sections (5 μm). These sections were stained with hematoxylin & eosin (HE) solution. HE-stained liver tissues were histologically examined using an Olympus AX70 light microscope, (Olympus Co., Ltd., Tokyo, Japan), equipped with 20× and 60× objective lenses.
Statistical analysis. These biochemical results were expressed as means±standard error of the mean (S.E). Statistical significance was performed using EXCEL 2015 ver.1.15 software. The Dunnett's multiple comparison test was used to compare relative gene expression levels and cell viability. p-Values of *p<0.05 and **p<0.01 were considered as statistically significant differences.
Effects of mulberry leaves (ML) on a murine hepatic steatosis model.
Global analysis of gene expression of the liver using RNA-Seq. Livers were dissected and stored in RNAlater® Solution (Thermo Fisher Scientific Inc., Waltham, MA, USA). Then, total RNA was extracted using a RNeasy Lipid Tissue Mini Kit (Qiagen Co., Ltd., Venlo, Netherlands). The RNA quality and quantity were assessed on a 2100 Bioanalyzer using the RNA 6000 Nano kit (Agilent Technologies Inc., Santa Clara, CA, USA).
One microgram of total RNA from the livers of the ND (n=3), WD (n=3), and ML (n=3) groups were applied to library preparation. Sequencing libraries were generated using TruSeq RNA Library Preparation Kit v2 (Illumina Inc., San Diego, CA, USA). Library construction procedures were as follows. Total RNA was purified to mRNA and then mRNA was fragmented and converted to double-stranded cDNA. Sequencing adapters containing index sequences were ligated to the end of the cDNA. Libraries were sequenced on Illumina HiSeq 2500 (Illumina Inc.,) to generate 100-bp reads.
Sequence reads were trimmed and filtered by quality, and these reads were mapped to GRCm38 (mm10) with the RNA-Seq mapping algorithm in CLC Genomics Workbench v9.5.3 (Qiagen Co., Ltd.). RNA-seq data analyses were performed using the CLC Genomics Workbench with Advanced RNA-Seq Plugin 1.5.
For principal component analysis (PCA), total counts for each individual gene were normalized as log CPM values and projected onto two dimensions. Two principal components are specified by the 2D plot. The first principal component specified the direction with the largest variability in data, whereas the second component specified the direction with the second largest variation.
An analysis of differentially expressed genes (DEGs) was conducted. DEGs were determined based on a false discovery rate (FDR)-corrected p-value <0.05 using the empirical analysis of digital gene expression test tool in the CLC Genomics Workbench.
Two-dimensional heat maps of expression values were created with each column corresponding to one sample and each row corresponding to one gene. Euclidean distance and average linkage were set as parameters for the hierarchical clustering of samples and genes. To reduce the number of genes, only 250 DEGs (FDR<000.1, absolute fold change>4) were used for clustering.

Effects of ML on histological features of Western diet (WD)-induced Hepatic Steatosis in a murine model. Livers were fixed with formaldehyde and examined after HE staining. Numerous large lipid droplets were observed in WD (middle), but not in the Normal group (left). In contrast to WD, the size of lipid droplets in the ML (right) group was smaller.
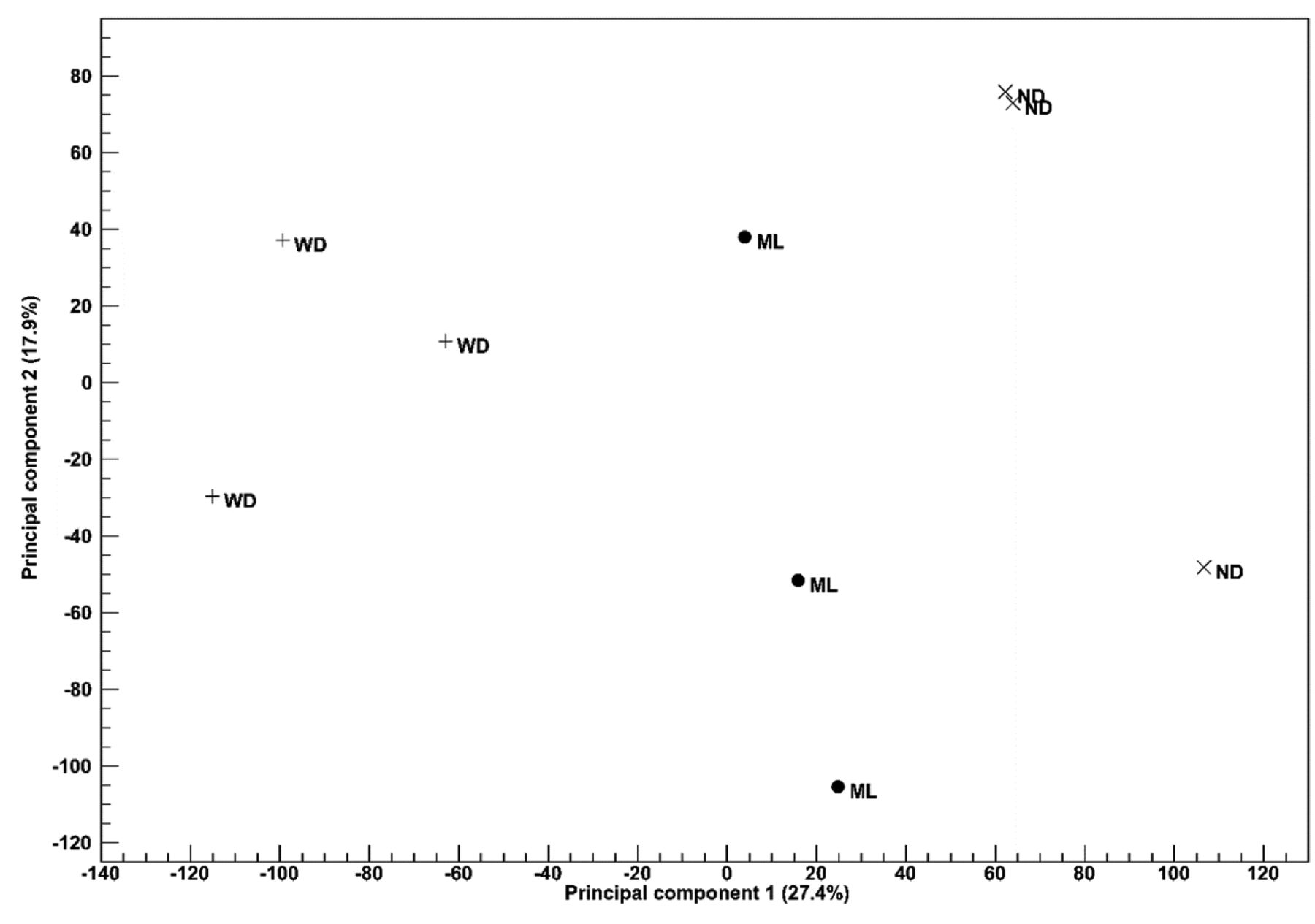
Two-dimensional principal component plot of principal component analysis for RNA-seq. The first principal component is shown for the X axis and the second principal component is shown for the Y axis. The value after the principal component identifier indicates the proportion of variance explained by this particular principal component. Shapes of symbols are assigned based on treatment (WD: Plus; ML: circle; ND: cross).
Functional analyses of DEGs were performed using the Ingenuity Pathway Analysis (IPA) software from Ingenuity Systems (Qiagen Co., Ltd.). Activation z-scores of gene networks were predicted using IPA. The activation z-scores represent activation status (activated or inactivated state) of bio-function and diseases related to gene networks.
Results
Change in body weight and plasma biochemical examinations. Significant increases in body weight and biochemical indicators were observed in the WD group compared with the ND group on the final day of the experiment. In contrast, ML suppressed the increase in TG, AST, and ALT. In contrast, no significant changes in body weight, T-Cho, and GLU were found in the ML group compared with that of the ND group (Table I).
Activated disease or functional annotation (|z-score| ≥2) for differentially expressed genes (DEGs) in Ingenuity Pathway Analysis.
Histological analysis of the liver. A lipid droplet was observed as white sections in HE staining. In the WD group, numerous depositions of lipid droplets were visible; in contrast, the size of lipid droplets was smaller in the ND and ML groups than in the WD group. The state of hepatic tissue in the ML group resembled that of the ND group (Figure 1).
Analysis of gene expression of the liver by RNA-Seq. For quality control of samples and their groups, PCA was conducted. Each group could be separated by the first principal component (27.4 %, horizontal axis) (Figure 2).
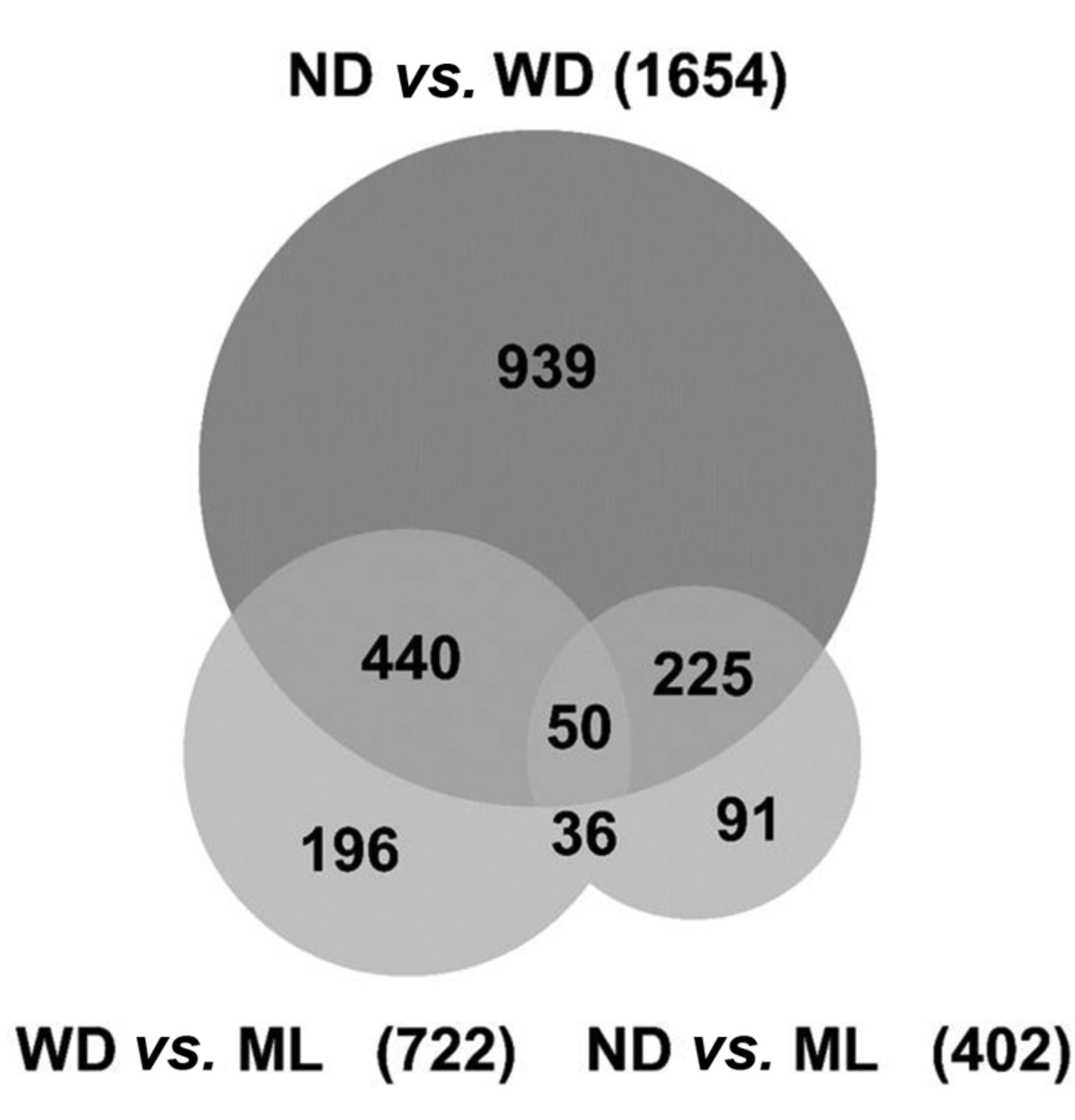
DEGs (FDR<0.05) were compared by a Venn diagram (Figure 3). The difference in number of DEGs between the WD group and other groups was larger in the ND group than in the ML group. The difference in DEGs between the WD group and ND was 1654 (962 were larger in WD, 692 were larger in ND). The difference in DEGs between the WD group and ML group was 722 (336 were larger in WD, 386 were larger in ML). The difference in DEGs between the ND group and ML group was 402 (155 were larger in ND, 247 were larger in ML).
Using the 250 DEGs chosen for expression analysis, clustering of individual mice (horizontal axis) and information between genes (vertical axis) were used to create a heat map (Figure 4). Clusters of samples were clearly divided into the 3 groups. Furthermore, ND and ML groups made a clade.

Venn diagram of differentially expressed genes. The number in each area indicates the number of DEGs. This Venn diagram summarizes the overlap of DEGs from ND vs. WD (the upper circle) and WD vs. ML (the left circle), and ND vs. ML (the right circle). WD: Western diet; ND: normal diet; ML: mulberry leaf.

Heat map of expression values of differentially expressed genes. Each column corresponds to one sample, and each row corresponds to a gene. The genes are hierarchically clustered by the similarity of their expression profiles over the set of samples and the samples are hierarchically clustered by the similarity of expression patterns over their features. Only differentially expressed genes (250) were used for creating the heat map.
Activation state of diseases and function were predicted according to the functional analysis of the DEGs (FDR<0.001) between the WD and ML groups using IPA (Table II). The results showed that organismal death and inflammatory responses were decreased, whereas cellular growth and proliferation, and adipogenesis of cells increased. Metabolic disorder (p-value=1.47E-03, z-score=−1.85) was decreased (not shown in the table).
Activation state of tox functions of DEGs between WD and ML were predicted in IPA. Inflammation of liver was decreased (p-value=0.01, z-score=−2.35) and regeneration of liver was increased (p-value=0.01, z-score=2.00). Figure 5 shows the expression networks of genes involved in each function.
Discussion
In the experiments of this study, mice were provided a high-calorie diet, and they were found to have the diseases that originated from metabolic syndrome, including increased body weight, hyperlipidemia, and liver impairment.
Biochemical tests revealed that ML suppressed the increase in biochemical indicators of lipid metabolism in the serum and liver (Table I). In addition, the suppressive effect of ML on lipid droplet accumulation in the liver was confirmed histologically (Figure 1). In particular, this can be considered a model of hepatic steatosis in humans, i.e. a model for NAFLD/NASH. Thus, ML was shown to have suppressive effects on the onset of HS caused by a high-calorie diet.
To elucidate the mechanism of ML on HS, RNA-Seq was conducted and compared DEGs in the WD and ML groups. In PCA (Figure 2) and heat map analysis (Figure 4), samples in the ML group were shown to have an intermediate state between that of WD and ND. These results indicated that ML recovered gene expression. The difference in DEGs was larger for WD vs. ND than WD vs. ML (Figure 3).
Decreases of organismal death and inflammation and increases of cellular growth and adipogenesis of cells were predicted in functional analysis of DEGs between WD and ML. These predictions were different from previously reported low molecular Lychee polyphenol administration (9).
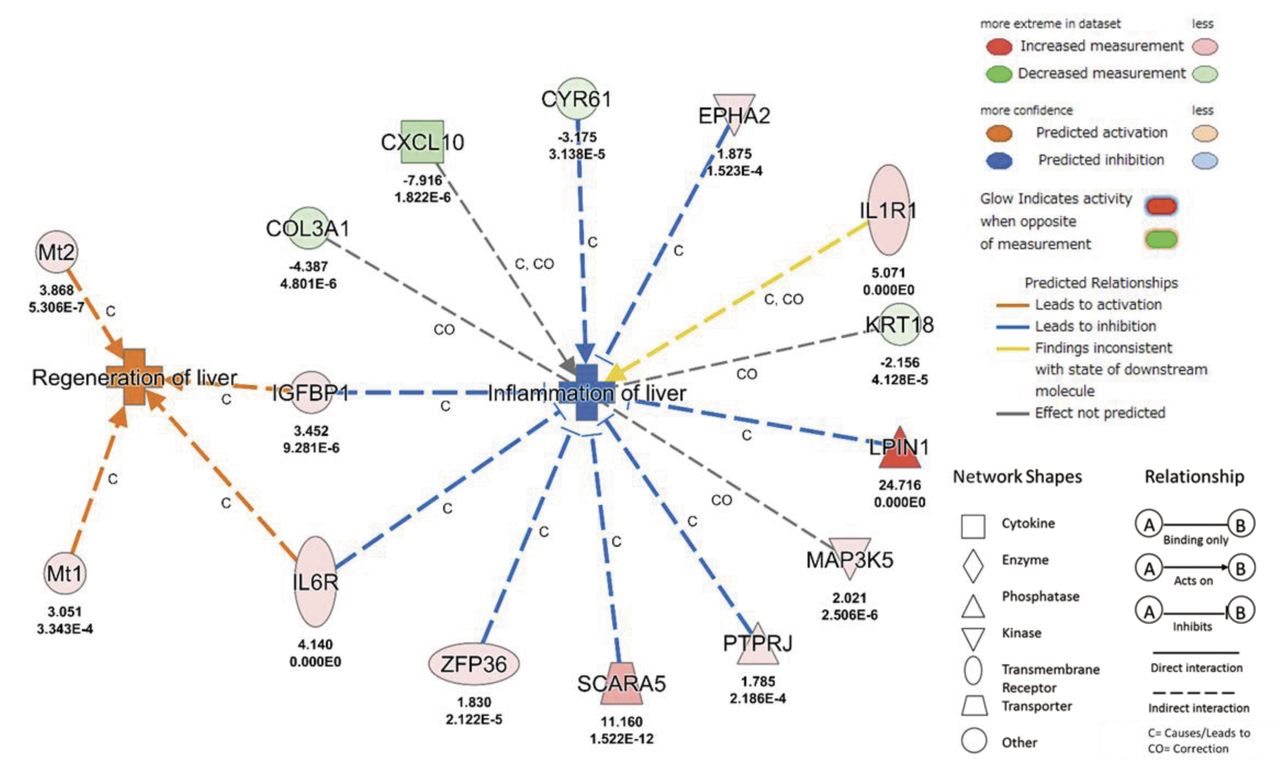
Gene networks of inactivated diseases or functions in WD versus ML groups. Attributed to ML feed, B) genes for the inflammation of the liver were inactivated. WD: Western diet; ML: mulberry leaf. The number below molecules are Log2 fold change ratio based on WD (upper) and FDR (lower). COL3A1: Collagen type III alpha 1 chain; CXCL10: C-X-C motif chemokine ligand 10; CYR61: cysteine rich angiogenic inducer 61; EPHA2: EPH receptor A2; IGFBP1: insulin like growth factor binding protein 1; IL1R1: interleukin 1 receptor type 1; IL6R: interleukin 6 receptor; KRT18: keratin 18; LPIN1: lipin 1; MAP3K5: mitogen-activated protein kinase kinase kinase 5; Mt1: metallothionein 1; Mt2: metallothionein 2; PTPRJ: protein tyrosine phosphatase, receptor type J; SCARA5: scavenger receptor class A member 5; ZFP36: ZFP36 ring finger protein.
Progression of such hepatic steatosis is closely associated with inflammation (10), oxidative stress, and lipid metabolism disorders, and can lead to the damage of liver cells, fibrosis, and malignant transformation due to factors such as over-production of reactive oxygen species (ROS), a weakened ROS elimination system, and changes in the hepatic innate immune cell (Kupffer cell, NK cell) networks (11, 12).
In toxicological functions of liver with DEGs between the WD and ML groups, gene expression change of CYR61 (13), EPHA2 (14), IGFBP1 (15), IL6R (16), LPIN1 (17), PTPRJ (18), SCARA5 (19), ZFP36 (20) lead to inhibition of inflammation of liver except ILR1 (21) and gene expression change of IGFBP1 (22), IL6R (23), Mt1 (24), Mt2 (24), lead to activation of regeneration of liver (Figure 5).
These data suggest that ML functions to suppress HS in WD-fed mice and regulates genes related to inflammation and regeneration of liver cells. To date, there have been many reports regarding the functional potential of ML, including its anti-oxidative action towards lipids and its sugar-absorption suppressing action (25, 26). However, the experiments in this study suggest the possibility that not only does ML reduce the accumulation of lipids in the liver, it also suppresses genes related to inflammation and organismal death of the liver, acting to suppress progression of hepatic steatosis, i.e. progression to liver fibrosis and liver cancer (Table II). Detailed investigation into the mechanism of these functions will require screening of components other than 1-deoxynojirimycin and flavonoids, which have been reported previously.
In conclusion, the RNA-Seq method was utilized to perform comprehensive measurements of genes in the liver, thereby examining the functionality of ML. Moreover, the results of this investigation hold promise with respect to increasing the use of ML for the prevention of hepatic steatosis that is becoming a societal problem (27).
Acknowledgements
This work was supported by the Hokkaido Pharmaceutical University School of Pharmacy collaborative research expenses from Life Mate Co., Ltd. This study was supported, in part, by the MEXT-Supported Program for the Strategic Research Foundation at Private Universities 2013-2017 (S1311017) and Agriculture, Forestry and Fishery promotion research project (Type A-H25, 25016A) from the Ministry of Agriculture, Forestry and Fisheries of the Japanese Government. The Authors also thank Rumi Ohtake and Eri Kubota for technical support of RNA-Seq.
- Received April 11, 2018.
- Revision received June 5, 2018.
- Accepted June 6, 2018.
- Copyright© 2018, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}