


Abstract
Background/Aim: Several proteins involved in immune regulation and the relationship among these, the tumor microenvironment, and clinical outcomes of eribulin treatment were evaluated in advanced or metastatic breast cancer patients. Patients and Methods: This retrospective cohort study comprised 52 eribulin-treated locally advanced or metastatic breast cancer patients. Cancer tissue samples were obtained before and after treatment in 10 patients. Immunohistochemistry was performed to determine programmed death (PD)-1, CD8, and forkhead box P3 (FOXP3) expression by stromal tumor-infiltrating lymphocytes, and PD-ligand (L1) and PD-L2 expression by cancer cells. Results: Of the 10 patients, 5 were responders (partial response) and 5 were non-responders (stable disease, 2; progressive disease, 3) to eribulin. PD-1, PD-L2, and FOXP3 expression became negative in 5 patients, PD-L1 expression became negative in 6 patients, and CD8 expression became positive in 3 patients after treatment. The response to eribulin was significantly associated with PD-L1 and FOXP3 negative conversion (p=0.024 and 0.004, respectively). The change in E-cadherin expression (positive or negative) was also correlated with the changes in PD-L1 and FOXP3 (p=0.024 and 0.004, respectively). Kaplan–Meier analysis with log-rank tests revealed that progression-free survival and time-to-treatment failure were significantly longer in patients with PD-L1 and FOXP3 negative conversion (p=0.012 and 0.001; p=0.049 and 0.018, respectively). Conclusion: The efficacy of eribulin may be attributed to its biological effects on the immune system (reduction of PD-L1 and FOXP3 expression) through epithelial–mesenchymal transition suppression, and vascular remodeling and improvement of the tumor microenvironment.
Breast cancer is the second most common cancer worldwide, and the most common cancer in women. In 2012, there were an estimated 1.67 million new cases of breast cancer worldwide, and 522,000 deaths were attributed to breast cancer. Advanced breast cancer, particularly stage IV, is associated with poor outcomes and a recent study reported a median survival rate of 2.3 years and 10-year actuarial breast cancer-specific survival rates of 15.7%, 14.9%, and 11.7%, respectively, for women aged ≤40, 41-50, and 51-70 years at the time of diagnosis (1).
Initial treatment for breast cancer usually comprises surgical resection and radiotherapy, depending on the patient's condition. Adjuvant or neoadjuvant therapy with chemotherapy or molecular targeted therapies may also be valuable depending on the breast cancer's receptor status or other risk markers, such as nodal status, tumor grade, and lymphovascular invasion. In many countries, including Japan (2), treatment decisions are increasingly being led by the presence or absence of biomarkers and patient choice (3-5). Although many agents have been developed to target hormone receptors, these are unsuitable for treating triple-negative breast cancer, underscoring the need to target alternative molecular pathways. To understand why existing chemotherapies frequently show limited long-term efficacy, researchers are increasingly focusing on the tumor microenvironment in order to develop molecular-targeted drugs. It has become clear that cancer cells exploit molecular pathways to evade the immune system and chemotherapies.
Cells within the tumor microenvironment, including tumor-infiltrating lymphocytes and cancer cells, may be used to monitor immune responses and predict treatment responses (6, 7). In particular, it has been demonstrated that T cells and cancer cell phenotypes associated with a poor antitumor response display abnormal expression of programmed cell death protein (PD)-1, programmed death ligand-1 (PD-L1), PD-L2, and forkhead box P3 (FOXP3) (8).
The epithelial–mesenchymal transition (EMT) is a critical cellular transition by which cancer cells switch from an epithelial phenotype to a mesenchymal phenotype that supports their ability to migrate and invade other tissues (9-11). It was reported that inhibition of EMT could reduce this immunosuppression (12).
Eribulin mesylate (eribulin) is an analog of halichondrin B that causes microtubule depolymerization by inhibiting the growth rate of microtubules without affecting microtubule shortening (13). Additionally, several studies have demonstrated that eribulin is clinically-effective in patients with breast cancer (14, 15). In a pooled analysis of patients with metastatic breast cancer who had received at least one prior chemotherapy regimen, eribulin significantly extended overall survival (OS), and increased the progression-free survival (PFS) and clinical benefit rates compared with the control treatments (15). The survival benefit of eribulin was also apparent in patients with human epidermal growth factor receptor 2 (HER2)-negative or triple-negative breast cancer (15). Interestingly, in a preclinical study, eribulin demonstrated unique effects such as reversed EMT and induced mesenchymal–epithelial transition (MET) (16). This process may explain the clinical observation of prolonged OS in eribulin-treated patients.
It is important to analyze the effects of eribulin on microenvironments, including the immune microenvironment, and on the extent of EMT. To date, however, no studies have examined the effects of eribulin on the antitumor immune response in breast cancer. Therefore, the objectives of this study were to evaluate the changes in the tumor microenvironment, focusing on the antitumor immune response, in patients with advanced or metastatic breast cancer treated with eribulin.
Materials and Methods
Ethics. The research reported here conformed to the provisions of the Declaration of Helsinki (64th WMA General Assembly, Fortaleza, Brazil, October 2013). This study comprised a retrospective chart review. At the time of treatment, patients provided written informed consent for the use of their data in later research studies. The study protocol was approved by the Ethics Committee of Osaka City University (#926).
Patients and treatments. We obtained and analyzed the electronic medical records of patients with metastatic breast cancer who underwent chemotherapy using eribulin from August 2011 to June 2013 at our institute. The cut-off was limited to June 2013 because few patients received eribulin after this date, and to ensure that there was sufficient follow-up. Patients received a chemotherapy regimen, in 21-day cycles, in which eribulin mesylate (1.4 mg/m2) was intravenously administered on days 1 and 8, followed by a drug-free period until day 21 (17). This protocol was repeated until progressive disease was detected or a severe adverse event requiring the discontinuation of the scheduled chemotherapy was noted. Eribulin was administered in an outpatient setting.
Survival endpoints. The antitumor effect was evaluated according to the Response Evaluation Criteria in Solid Tumors version 1.1 (18). The overall response rate (ORR), clinical benefit rate (CBR), disease control rate (DCR), OS, time to treatment failure (TTF), and PFS were calculated as follows. The ORR was calculated as the sum of patients with complete response (CR) or partial response (PR). The CBR was calculated as CR + PR + stable disease (SD) (for >24 weeks)/total number of patients. The DCR was calculated as CR + PR + SD (other)/total number of patients. The OS was evaluated daily and was calculated as the time from the start of treatment to the patient's death. The TTF was evaluated daily and was calculated as the time from the start of treatment to discontinuation of treatment for any reason, including disease aggravation, adverse events, or death. The PFS was evaluated daily and was calculated as the time from the start of treatment to either confirmation of progressive disease or death, whichever occurred first.
Tumor morphology and grading. Tumor morphology was assessed using tissue samples obtained by core needle biopsy at diagnosis or during breast cancer resection. Morphologic assessments included histological tissue type and nuclear grade, as evaluated by conventional hematoxylin and eosin (HE) staining. Breast cancer was classified into subtypes according to the immunohistochemical expression of estrogen receptor (ER), progesterone receptor (PgR), HER2, and Ki67, yielding the following immunophenotypes: luminal A (ER+ and/or PgR+, HER2−, Ki67low), luminal B (ER+ and/or PgR+, HER2−, Ki67high), luminal HER2 (ER+ and/or PgR+, HER2+), HER2-enriched (ER−, PgR−, and HER2+), and triple-negative breast cancer (ER−, PgR−, and HER2−) (19).
Immunohistochemical scoring was performed by two pathologists (M.O. and S.T.) specialized in mammary gland pathology who were blinded to the patient details to ensure the diagnosis was objective and reproducible. The cut-off values for ER and PgR positivity were set at ≥1% in accordance with previous studies (20). HER2 expression was scored using an accepted grading system (0=no reactivity, or membranous reactivity in less than 10% of cells; 1+=faint/barely perceptible membranous reactivity in ≥10% of cells or reactivity in only part of the cell membrane; 2+=weak to moderate complete or basolateral membranous reactivity in ≥10% of tumor cells; or 3+=strong complete or basolateral membranous reactivity in ≥10% of tumor cells). HER2 expression was considered positive if the immunostaining score was 3+ or 2+ together with gene amplification via fluorescent in situ hybridization (FISH). For FISH analyses, each copy of the HER2 gene and its centromere 17 (CEP17) reference were counted. The interpretation followed the American Society of Clinical Oncology/College of American Pathologists guidelines for HER2 immunohistochemical classification of breast cancer, and was considered positive if the HER2/CEP17 ratio was >2.0 (21). A Ki67-labeling index of ≥14% was classified as positive (19).
Immunohistochemistry. Immunohistochemistry was performed using tissue samples obtained by core needle biopsy at diagnosis or during breast cancer resection. Tumor specimens were fixed in 10% formaldehyde solution and embedded in paraffin, and 4-μm-thick sections were mounted onto glass slides. Slides were deparaffinized in xylene and heated for 20 min (105°C, 0.4 kg/m2) in an autoclave in Target Retrieval Solution (Dako, Carpinteria, CA, USA). Specimens were then incubated with 3% hydrogen peroxide in methanol for 15 min to block endogenous peroxidase activity, and then incubated in 10% normal goat or rabbit serum to block non-specific reactions. Primary monoclonal antibodies directed against E-cadherin (clone NCH 38; Dako), PD-1 (clone NAT105; Abcam, Cambridge, UK), PD-L1 (clone 27A2; MBL, Nagoya, Japan), PD-L2 (clone #176611; R&D Systems, Minneapolis, MN, USA), FOXP3 (clone 236A/E7; Abcam), and CD8 (clone C8/144B; Dako) were used. Tissue sections were incubated with each antibody for 70 min at room temperature or overnight at 4°C, and then incubated with horseradish peroxidase-conjugated anti-rabbit or anti-mouse Ig secondary antibodies (HISTOFINE [PO]™ Kit; Nichirei, Tokyo, Japan). The slides were subsequently treated with streptavidin-peroxidase reagent and incubated in phosphate-buffered saline-diaminobenzidine and 1% hydrogen peroxide (v/v), followed by counterstaining with Mayer's hematoxylin. Positive and negative controls for each marker were used according to the manufacturer's data sheets for each antibody.
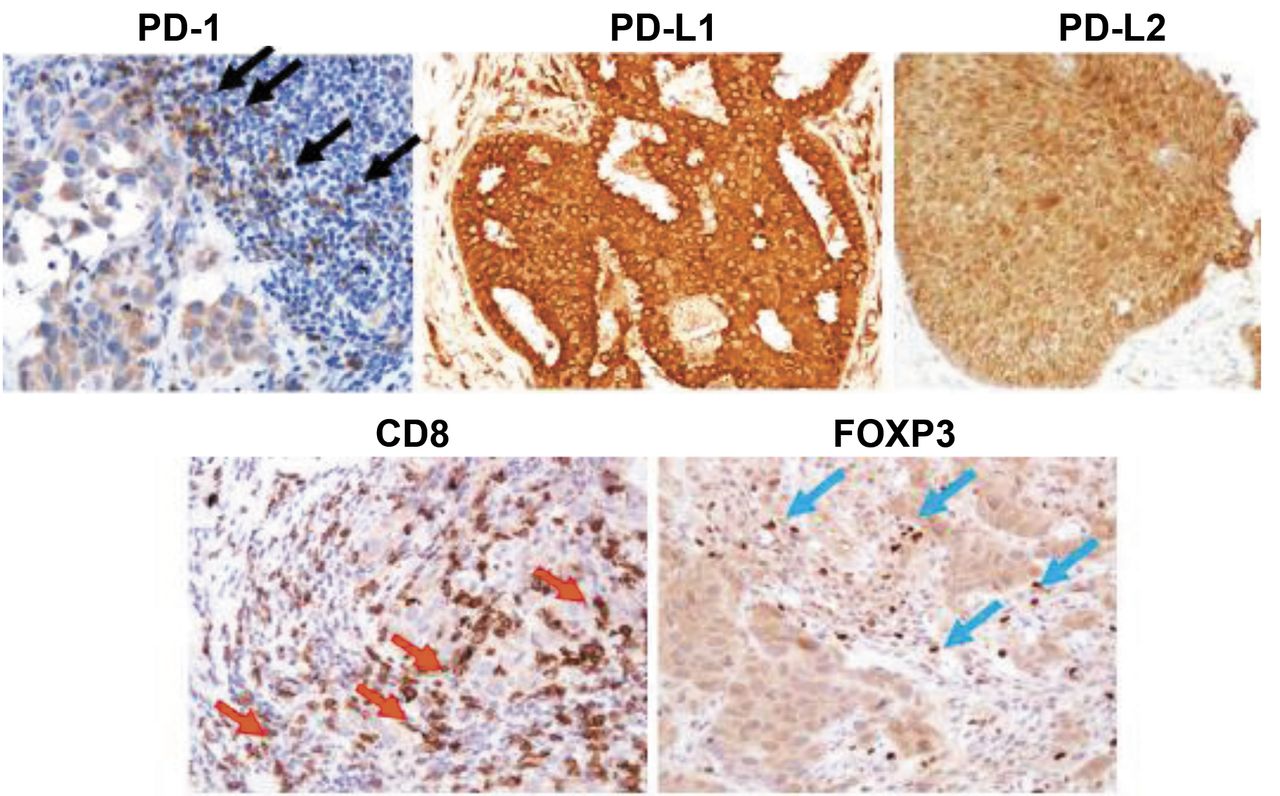
PD-1 is a cell surface receptor that plays an important role in negative regulation in the immune system. Its ligands, PD-L1 and PD-L2 are also cell surface proteins and can bind to PD-1. FOXP3, a marker of regulatory T cells that has a suppressive function for effector T cells, such as CD8+ killer T cells, was evaluated. To evaluate PD-1, FOXP3, and CD8 expression, the number of tumor-infiltrating lymphocytes in the stroma surrounding cancer cells labelled with anti-PD-1, anti-FOXP3, or anti-CD8 antibodies was counted at a magnification of 200× in each of three fields of view (FOVs) selected in darkly stained areas. In accordance with previous studies (22, 23), the median value of the mean of all three FOVs was determined, and that number was set as a cut-off value. To evaluate PD-L1, PD-L2, and E-cadherin, three FOVs in darkly stained areas were selected, and the percentage of cancer cells stained with the corresponding antibody in each FOV was determined at a magnification of 400×. In accordance with a previous study (24), the median value of the mean of all three FOVs was determined; high expression was defined as expression in ≥10% of cells for PD-L1, PD-L2, and E-cadherin. Representative immunohistochemical slides are shown in Figure 1.
It was previously reported that eribulin regulated EMT (16), and we reported that E-cadherin expression became positive in tissues from locally advanced or metastatic breast cancer patients with objective clinical response to eribulin treatment (25). Therefore, the relationship between EMT and immune regulation by eribulin was analyzed.
Statistical analyses. Statistical analyses were performed using SPSS® version 19.0 statistical software (IBM, Armonk, NY, USA). Categorical data are represented as numbers and percentages, and continuous data are represented as medians and range. The association between treatment efficacy and clinicopathological variables was analyzed using Fisher's exact test. Survival rates were evaluated using Kaplan–Meier plots and log-rank tests. In all analyses, p<0.05 was considered statistically significant. The cut-off values for the biomarkers included in this study were chosen before conducting the statistical analysis.
Results
Patients. Overall, 52 patients were identified from their medical records, and the median follow-up time was 431 days (range=50-650 days). The mean age of patients was 63.5 years (Table I). One-quarter of the patients had locally advanced cancer and 75% had visceral metastases. Overall, 51.9%, 38.5%, 9.6%, and 50.0% of patients were ER+, PgR+, HER2+, or Ki67+, respectively; 42.3% of patients had triple-negative breast cancer. In 11 patients, tissue samples were obtained at diagnosis and during surgical resection after being treated with eribulin (surgical cohort). Their characteristics were consistent with those of the overall cohort.
Survival and cancer outcomes in the overall cohort. Table II shows the survival and cancer outcomes of the total cohort and of the 11 patients in the surgical cohort. In the overall cohort, the median OS, PFS, and TTF were 431 days (range=50-650 days), 325 days (range=40-650 days), and 155 days (range=49-650 days), respectively. CR was achieved in 1 patient (1.9%), PR in 17 patients (32.7%), and SD in 9 patients (17.3%) in the overall cohort. Five patients had SD for >24 weeks and 4 patients had SD for ≤24 weeks. The ORR, CBR, and DCR were 34.6% (18/52), 44.2% (23/52), and 51.9% (27/52), respectively, in the overall cohort. These rates tended to be lower in triple-negative breast cancer subtypes than in other cancer subtypes.
Immunohistochemistry. Tissue samples were obtained at diagnosis and after surgical resection and treatment with eribulin in 11 patients, but one patient had pathological CR and tissue samples were unavailable for immunohistochemistry. Therefore, immunohistochemical data were available for 10 patients: 5 with PR and 5 non-responders. In general, eribulin was administered within 1 month of diagnosis, and the resections were performed within 1 month of determining the effect of eribulin. Using the immunohistochemical data for these 10 patients, the expression rate of PD-1, PD-L1, PD-L2, CD8, and FOXP3 in each tissue specimen before and after eribulin treatment was plotted (Figure 2). The expression levels of PD-1, PD-L1, and FOXP3 decreased in all 5 responders to eribulin, increased CD8 expression was observed in 4/5 responders, and expression of PD-L2 was decreased in 4/5 responders. The positive or negative conversions in the expression levels of these markers tended to occur in opposite directions in non-responders. The response to eribulin was significantly associated with PD-L1 and FOXP3 negative conversion (p=0.024 and 0.004, respectively; Table III).
Regarding the analysis of the relationship between EMT and immune regulation by eribulin, the change in E-cadherin expression (positive or negative) was also correlated with the changes in PD-L1 and FOXP3 (p=0.024 and 0.004, respectively; Table IV).
Patient characteristics.
Kaplan–Meier analysis (Figure 3) with log-rank tests revealed that PFS and TTF were significantly longer in patients with PD-L1 negative conversion (p=0.012 and 0.001, respectively) compared with other patients. In the case of FOXP3 negative conversion, PFS and TTF were also significantly longer in patients with PD-L1 negative conversion (p=0.049 and 0.018).
Discussion
In this retrospective study, we assessed the treatment and survival outcomes after eribulin treatment in a cohort of 52 Japanese patients with metastatic breast cancer. The effects of eribulin on the tumor microenvironment and expression of immune-related biomarkers was also examined in 10 patients in whom immunochemistry was performed at diagnosis and after treatment with eribulin. The median OS, PFS, and TTF were 431, 325, and 155 in the overall cohort, and the ORR, CBR, and DCR were 34.6%, 44.2%, and 51.9%, respectively. In a pooled analysis of patients who had received at least one prior chemotherapy, the median OS and PFS for eribulin were 15.0 months and 3.9 months, respectively, while the CBR was 30% (15). Therefore, our results in this cohort of Japanese patients were generally similar to those in two European phase 3 trials (15).
Changes in the tumor microenvironment were also examined in 10 patients in whom tissue samples were available before and after treatment with eribulin. Although we observed some variability in the microenvironmental responses, the results suggest that responders to eribulin were more likely to show decreases in PD-L1 and FOXP3 expression compared with non-responders. The decrease of the inhibitory molecules in the responders and the increase in CD8, which may be caused by an influx on CD8+ T cells, suggest that eribulin promotes an antitumor immune response, and that these changes were associated with longer PFS and TTF than in non-responders.
It has been proposed that eribulin renders residual tumors less aggressive and less likely to metastasize by triggering a shift from mesenchymal to epithelial phenotypes by reversing EMT to MET (16). In addition, we have previously reported that all eribulin responders demonstrated positive conversion to E-cadherin expression, suggesting that eribulin inhibited EMT (25). A variety of factors in the tumor microenvironment can promote EMT. In particular, inflammation, hypoxia, and stem cells in the tumor microenvironment are inextricably linked to the EMT through complex pathways that are only now becoming fully understood (10). One key mediator may be hypoxia inducible factor-1α (HIF-1α), which is induced by hypoxia and growth factors. HIF-1α was reported to mediate EMT and cancer cell metastasis by upregulating Snail and TWIST1 expression, and hence promoting the morphological change of epithelial cells to a mesenchymal phenotype (26). Interestingly, eribulin was reported to improve tumor perfusion in human breast cancer tumor models, an effect driven by vascular remodeling associated with increased numbers of smaller functional microvessels. The increase in tumor perfusion alleviates hypoxic stress (and hence HIF-1α) associated with the abnormal tumor microenvironment (27).
It was also reported that eribulin induced reoxygenation by vascular remodeling in advanced breast cancer patients (28). In our previously study, CA9 expression, an indicator of hypoxia, was significantly decreased in eribulin responders suggesting that eribulin improved the hypoxic condition (25). The researchers mentioned that TGF-β, which is typically associated with hypoxic conditions, was decreased in the eribulin-treated group. TGF-β plays crucial roles in EMT and in immune regulation, including the induction of regulatory T cells and the suppression of effector T cells. It was found that hypoxia induced EMT (29, 30) and increased the expression of PD-L1 via the HIF-1 transcription factor on myeloid-derived suppressor cells (31). Recently, researchers demonstrated that PD-L1 is up-regulated in EMT-activated human breast cancer cells by a mechanism involving ZEB-1 and miR-200 (32).
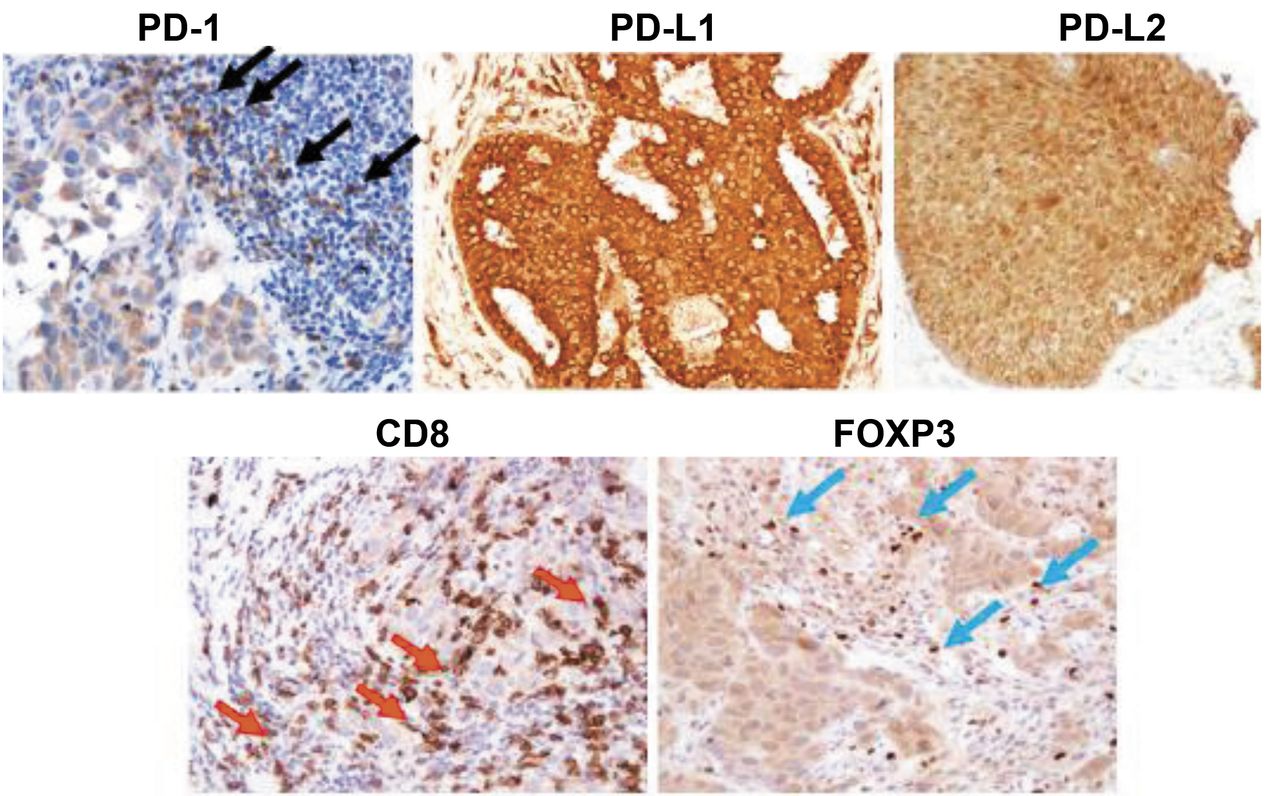
Representative immunohistochemical staining. Black arrows point to PD-1+ lymphocytes, red arrows point to CD8+ lymphocytes, and blue arrows point to FOXP3+ lymphocytes. The PD-L1- and PD-L2-positive cells show cytoplasmic staining. PD-1: Programmed death 1; PD-L: programmed death ligand; FOXP3: forkhead box P3.

Immunohistochemical analysis of immune biomarkers in metastatic breast cancer samples before and after treatment with eribulin (evaluable patients, n=10). % refers to the percentage of positive cells. The blue squares/lines show the results for responders and the red triangles/lines show the results for non-responders. PD-1: Programmed death 1; PD-L: programmed death ligand; FOXP3: forkhead box P3.
Survival outcomes.
Correlation between the response to eribulin and immune biomarker expression.
Correlation between E-cadherin expression and changes in immune biomarkers.
Our results suggest that eribulin may also inhibit an immune evasion mechanism involving immune checkpoints and improve the cancer immune microenvironment in breast cancer (Figure 4). Taken together, it is possible that eribulin regulates EMT and immune responses through vascular remodeling. Kudo-Saito et al. reported that in melanoma cells with typical EMT features, upregulation of Snail-induced regulatory T cells and impaired dendritic cells in vitro and in vivo, partly through thrombospondin-1 production (12). Meanwhile, Snail knockdown reduced the effects of Snail in terms of tumor metastasis and immunosuppression, and was followed by tumor inhibition along with local infiltration of antitumor effector cells in tumor tissues and systemic induction of tumor-specific CD8+ T-cell responses in vivo (12). In this manner, EMT also regulated immune functions in tumor tissues. We have previously reported that the number of tumor-infiltrating lymphocytes is a predictive marker for eribulin-responders (33). Our updated results indicate that eribulin accelerated the anti-tumor immune response and also showed efficacy in patients with metastatic breast cancer.
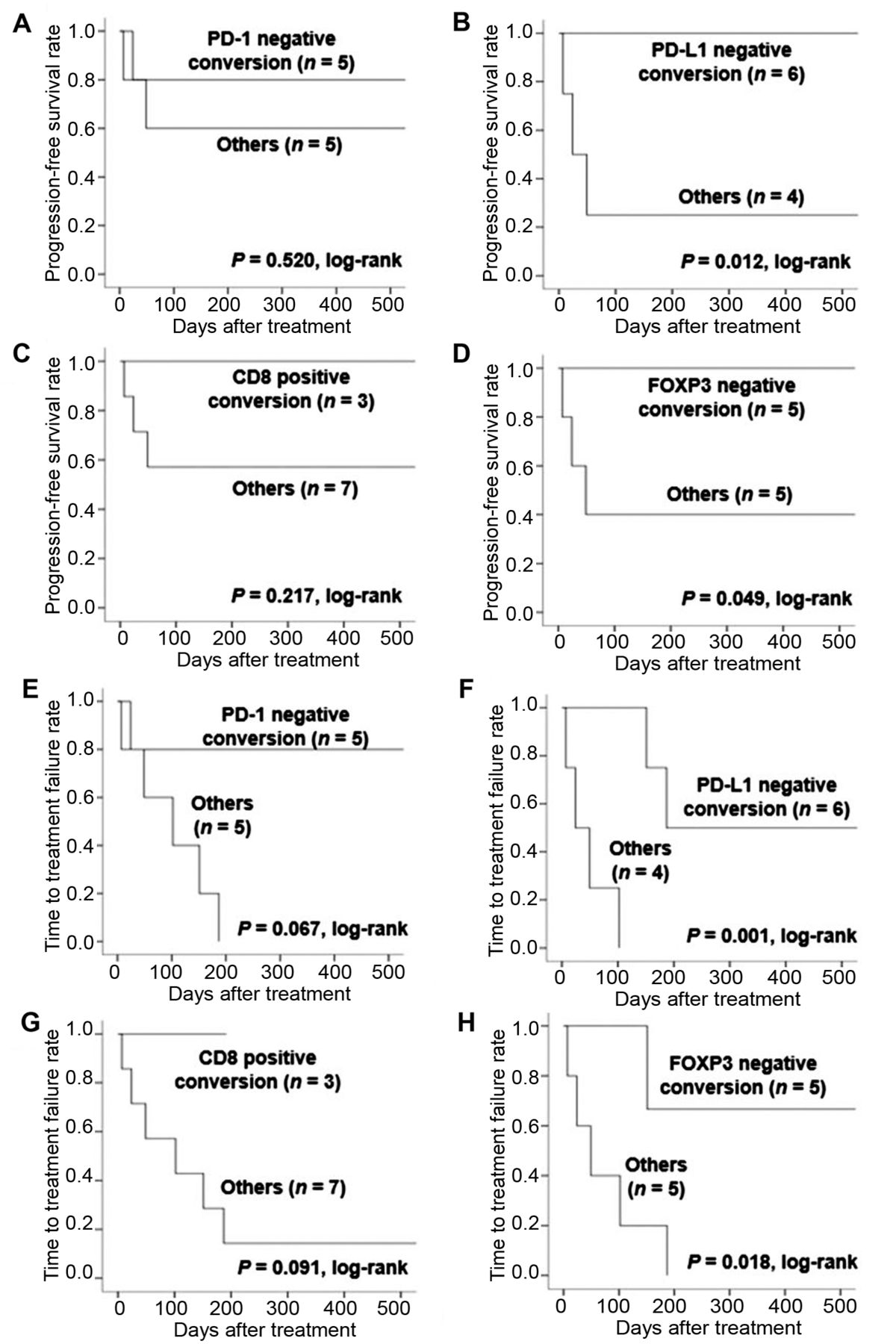
Progression-free survival (A-D) and time to treatment failure (E-H) in the surgical cohort according to changes in PD-1, PD-L1, CD8, and FOXP3 (n=10). PD-1: Programmed death 1; PD-L: programmed death ligand; FOXP3: forkhead box P3.
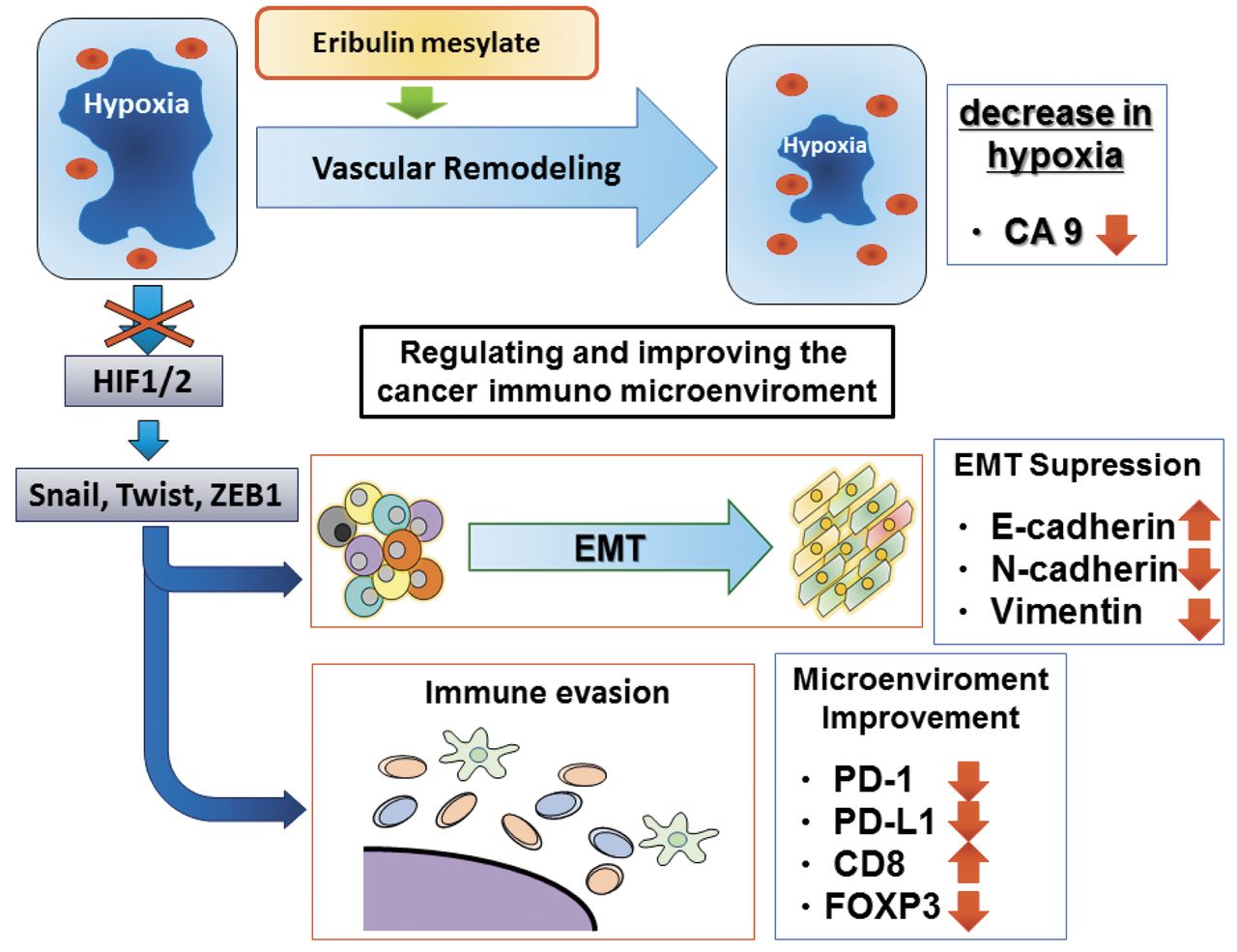
Proposed molecular mechanism of eribulin in metastatic breast cancer. EMT: Epithelial-mesenchymal transition; HIF: hypoxia-inducible factor; PD-1: programmed death 1; PD-L: programmed death ligand; FOXP3: forkhead box P3.
Interestingly, some non-responders showed favorable responses to eribulin in that the changes in protein expression were consistent with those in responders. The reason why these patients did not experience a tumor response is unclear and warrants clarification. It is feasible that the beneficial effects of eribulin on the tumor microenvironment were insufficient to counteract other cellular mechanisms involved in cancer progression. If so, some patients may benefit from being administered eribulin in combination with drugs targeting independent or complementary pathways. Alternatively, the disease may have been too advanced in some non-responders to observe a clear benefit from eribulin.
It is important to acknowledge that immunohistochemistry was performed using tissue samples before and after treatment with eribulin in just 10 of 52 patients. Therefore, the results might not fully reflect the changes in expression in the wider population of patients with advanced or metastatic breast cancer treated with eribulin.
The results of this study and those of prior studies, lead to the conclusion that by inhibiting the EMT, eribulin may enhance the antitumor immune response by improving the cancer immune microenvironment. In patients with locally advanced or metastatic breast cancer, the efficacy of eribulin might be attributed to the biological effects of eribulin on the immune system on the basis of EMT and/or vascular remodeling.
Acknowledgements
The Authors would like to thank Sayaka Tanaka from the Department of Diagnostic Pathology, Osaka City University Graduate School of Medicine, for providing assistance with immunohistochemical scoring. The Authors also thank Yayoi Matsukiyo and Tomomi Okawa (Department of Surgical Oncology, Osaka City University Graduate School of Medicine) for their helpful advice regarding data management. This study was funded by grants from the Japan Society for the Promotion of Science (KAKENHI, Nos. 25461992, 26461957, and 17K10559) to Shinichiro Kashiwagi. The Authors thank Nicholas D. Smith (Edanz Medical Writing) for medical writing support, which was funded by Eisai Co., Ltd.
Footnotes
This article is freely accessible online.
Conflicts of Interest
The Authors have no conflicts of interest to disclose.
- Received March 5, 2018.
- Revision received March 23, 2018.
- Accepted March 26, 2018.
- Copyright© 2018, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Doublet and Triplet Combinations of Eribulin, Fulvestrant, and Palbociclib in Preclinical Breast Cancer Models
- Eribulin Treatment Promotes Re-expression of Estrogen Receptor in Endocrine Therapy-resistant Hormone Receptor-positive Breast Cancer Cells
- The Modified Glasgow Prognostic Score and Prognostic Nutritional Index as Prognostic Markers in Patients With Metastatic Breast Cancer Treated With Eribulin
- Utility of Follow-up With Absolute Lymphocyte Count in Patients Undergoing Eribulin Treatment for Early Detection of Progressive Advanced or Metastatic Breast Cancer
- Molecular Biomarkers of Response to Eribulin in Patients with Leiomyosarcoma
- Dynamic Changes in Absolute Lymphocyte Counts During Eribulin Therapy Are Associated With Survival Benefit
- Ovarian carcinosarcoma genomics and pre-clinical models highlight the N-MYC pathway as a key driver and susceptibility to EMT-targeting therapy
- C-Reactive Protein and Absolute Lymphocyte Count Can Predict Overall Survival of Patients Treated With Eribulin
- Validation of Systemic and Local Tumour Immune Response to Eribulin Chemotherapy in the Treatment of Breast Cancer
- The Effects of Eribulin on Breast Cancer Microenvironment Identified Using Eribulin-resistant Breast Cancer Cell Lines