


Abstract
Neuroblastoma is one of the most common childhood malignancies and the primary cause of death from pediatric cancer. Derivatives of 17β-estradiol, 2-methoxyestradiol, as well as selective estrogen receptor modulators, such as fulvestrant, are novel potentially active anticancer agents. In particular, 2-methoxyestradiol is effective in treatment of numerous malignancies, including breast and prostate cancer, Ewing sarcoma, and osteosarcoma. Herein, we treated neuroblastoma SH-SY5Y cells with physiologically and pharmacologically relevant concentrations of 2-methoxyestradiol. We used flow cytometry in order to determine cell viability, cell death, level of nitric oxide and mitochondrial membrane potential. We demonstrated that at pharmacologically relevant concentrations, 2-methoxyestradiol results in induction of apoptosis of neuroblastoma SH-SY5Y cells via nitric oxide generation and reduction of mitochondrial membrane potential. Based on the obtained data, we propose that 2-methoxyestradiol may be a natural modulator of cancer cell death and survival through nitro-oxidative stress-dependent mechanisms. Moreover, the results confirm the efficiency of 2-methoxyestradiol in treatment of neuroblastoma.
Estrogens, a type of female sex steroids, are important factors in the brain and may also play an important role in non-reproductive functions (1). Interestingly, they are able to directly impact on other behaviors in brain regions, thus controlling mood and cognition, enhancing memory or inducing neuroprotection (1).
2-Methoxyestradiol (2-ME) is a natural metabolite of 17β-estradiol (E2), a hormone of both women and men. 2-ME is a monomethyl ether of 2-hydroxyestradiol, formed in the reaction catalyzed by catechol-O-methyltransferase. Its physiological level in blood serum ranges from 30 pM up to 30 nM during pregnancy (1), while pharmacologically relevant concentrations are micromolar. Interestingly, 2-ME is a novel potentially effective anticancer agent, active after oral administration. 2-ME in pharmacological doses affects cancer cell growth both in vivo and in vitro (2-5). 2-ME (branded as Panzem) is currently being evaluated in advanced phases of clinical trials (3-5). In spite of its proven anticancer activity, the molecular mechanisms of 2-ME remain unclear. 2-ME directly inhibits angiogenesis and induces apoptosis in tumorous and rapidly proliferating cells. Additionally, it takes part in stress-induced apoptosis due to the generation of reactive oxygen (ROS) and nitrogen (RNS) species (6-8). Common targets for 2-ME are mutated p53, or B-cell lymphoma 2 (BCL2) family proteins. 2-ME induces both extrinsic and intrinsic apoptotic pathways associated with the overexpression of p53 (2).
We previously reported the dual facets of 2-ME, namely that it causes cancer cell death, but on the other hand may also play a key role as a neurotoxin (6, 8). We observed anticancer effects of 2-ME in osteosarcoma cells even under physiological concentrations (8). Though, neuroprotective effects of E2 are well known, there are inconsistent data concerning the action of 2-ME (9, 10). Previously, we demonstrated that 2-ME leads to cancer cell and hippocampal cell death through selective induction of neuronal nitric oxide synthase and nitric oxide (NO) generation.
In the current study, we used the human dopaminergic neuroblastoma SH-SY5Y cell line, subline of SKNSH (9-13). Neuroblastoma is a childhood malignancy and the primary cause of death from pediatric cancer for children between the age of 1 and 5 years, accounting for approximately 13% of all pediatric cancer mortality (14, 15). It is a complex and heterogeneous disease. Neuroblastoma remains distinct from other cancer types because of its biological heterogeneity and range of clinical behavior, from spontaneous regression to highly metastatic disease (14, 15).
Thus, we aimed to determine whether 2-ME exerts any anticancer effects on neuroblastoma SH-SY5Y cells at physiologically and pharmacologically relevant concentrations.
Materials and Methods
Reagents. Tissue culture media, antibiotic cocktail, fetal bovine serum and 2-ME, were purchased from Sigma-Aldrich (Poznan, Poland).
Cell line and culture conditions. The human neuroblastoma SH-SY5Y cell line was obtained from Sigma Aldrich (Poznan, Poland). The cells were cultured at 37°C in a humidified atmosphere saturated with 5% CO2 using Ham's F12:EMEM (EBSS) (1:1) supplemented with 2 mM glutamine, 1% non-essential amino acids, and 15% fetal bovine serum (Sigma Aldrich).
Cell treatment. The neuroblastoma SH-SY5Y cells were treated with different concentrations of 2-ME at physiological (10−10-10−8 M) and pharmacological (10−6, 10−5 M) concentrations and for different times, depending on the design of experiments. In order to exclude potential influence of serum-derived hormones, all experiments were performed in medium without fetal bovine serum. The data were obtained from at least three independent experiments (N=3 replicate cultures).
Assessment of cell viability. SH-SY5Y cells were seeded onto six-well plates at a density of 3×105 cells per well. After 24 h of culture in standard medium, the cells were treated with 2-ME for an additional 24 h. The cells were then pelleted and incubated with Muse Count and Viability Reagent according to the manufacturer's protocol (Merck Millipore,Warsaw, Poland). Afterward, the cells were analyzed (5,000 events/sample). The signals were detected using the Muse Cell Analyzer (Merck Millipore) which is based on flow cytometric technology. The results were then analyzed by Muse 1.4 analysis software. Each experiment was performed at least three times.
Assessment of induction of cell death using annexin V and 7-amino-actinomycin D (7-AAD) staining. The Muse™ Annexin V & Dead Cell Assay allows for the quantitative analysis of cell-death induction. SH-SY5Y cells were seeded onto six-well plates at a density of 300,000 cells per well. After 24 h of culture in standard medium, the cells were treated with 2-ME for an additional 24 h. The cells were then pelleted and incubated for 20 min with the Muse™ Annexin V & Dead Cell Assay reagents according to the manufacturer's protocol. Afterward, the cells were analyzed (5,000 events/sample), and the signals were detected using the Muse Cell Analyzer. The results were then analyzed by the Muse 1.4 analysis software. Each experiment was performed at least three times.
Mitochondrial membrane depolarization. The neuroblastoma SH-SY5Y cells were seeded onto 10 cm culture dish plate at a density of 2×106 cells/dish. The cells were treated with 2-ME for 2 h. The cells were then pelleted and incubated for 20 min with The Muse™ MitoPotential Reagent, a cationic, lipophilic dye (Merck Millipore), according to manufacturer's protocol. Afterward, the cells were analyzed (5,000 events/sample), and the signals were detected using the Muse Cell Analyzer (Merck Millipore, Poland). The results were then analyzed by Muse 1.4 analysis software. Each experiment was performed at least three times.
Level of NO. The neuroblastoma SH-SY5Y cells were seeded onto 10 cm culture dish plate at a density of 2×106 cells/dish. The cells were treated with 2-ME for 2 h. The cells were then pelleted and incubated for 30 min with Muse® Nitric Oxide Reagent, a membrane-permeable novel reagent DAX-J2 Orange (Merck Millipore), according to the manufacturer's protocol. Afterward, the cells were analyzed (5,000 events/sample), and the signals were detected using the Muse Cell Analyzer. The results were then analyzed by Muse 1.4 analysis software. Each experiment was performed at least three times.
Statistical analysis. The results represent the mean±SD from at least three independent experiments. Differences between control samples and 2-ME-treated samples were assessed with one-way analysis of variance (ANOVA) with post hoc testing using a Dunnett's multiple comparison test. A p-value of less than 0.01 was considered to correspond with statistical significance. Data were analyzed using GraphPad Prism (version 6; GraphPad Software, Inc., La Jolla, CA, USA).
Results
Impact of 2-ME on neuroblastoma SH-SY5Y cell viability. The first goal of the study was to determine the antiproliferative effects of 2-ME on neuroblastoma SH-SY5Y cells. SH-SY5Y cells were treated with a series of selected, representative concentrations: physiological (10−10-10−8 M) and pharmacological (10−6, 10−5 M) concentrations (Figure 1A) for 24 h. Subsequently, the impact on cell viability was observed. As presented in Figure 1A, 2-ME effectively inhibited neuroblastoma cell viability only at pharmacologically relevant concentrations. We observed a significant reduction of cell viability after 24 h treatment with 10−5 and 10−6 M 2-ME, respectively, as compared to the control (C) (Figure 1A).
Impact of 2-ME on neuroblastoma SH-SY5Y cell death. Next, we determined the impact of 24 h treatment with physiologically and pharmacologically relevant concentrations on induction of cell death (Figure 1B and C). As demonstrated, a significant increase in apoptotic cells as compared to the control was observed only after 24 h treatment with 10−6 and 10−5 M, pharmacologically relevant, concentrations of 2-ME, respectively (Figure 1B). Interestingly, we did not observed induction of necrosis in SH-SY5Y cells by 2-ME (Figure 1C).
Impact of 2-ME on the generation of NO in neuroblastoma SH-SY5Y cells. Previously, we demonstrated that 2-ME increases NO level in osteosarcoma cells, resulting in cell death or cell-cycle arrest (8). Herein, neuroblastoma SH-SY5Y cells were treated for 2 h with 2-ME at physiologically and pharmacologically relevant concentrations, the level of NO was determined. In our recent studies, we confirmed that 2 h incubation with 2-ME is sufficient to observe generation of RNS (8). Here, we found an increased level of intracellular NO after 2-h treatment of SH-SY5Y cells with all used concentrations of 2-ME (Figure 2A). We noticed a significant increase in NO-positive after treatment at all concentrations of 2-ME, respectively (Figure 2A).
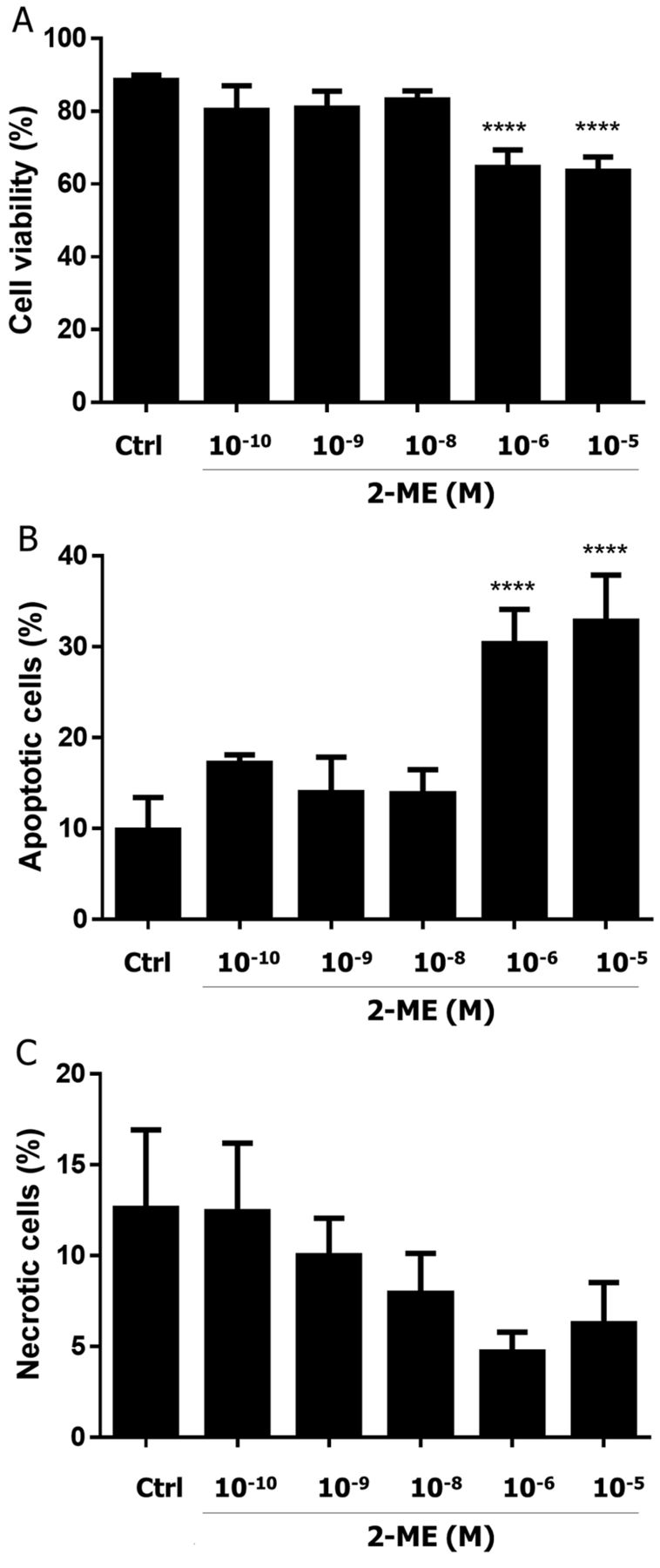
Impact of 2-methoxyestradiol (2-ME) on cell viability and cell death induction of neuroblastoma SH-SY5Y cells. SH-SY5Y cells were treated with a series of selected, representative concentrations: physiological (10−10-10−8 M) and pharmacological (10−6 and 10−5 M) for 24 h. Inhibition of cell growth (A), induction of apoptosis (B) and necrosis (C) by 2-ME were determined using the Muse Cell Analyzer. Values are the mean±SE of three independent experiments (N=3 replicate cultures). The absence of an error bar denotes a line thickness greater than the error. ****p<0.00001 versus control cells (Ctrl). Each experiment was performed at least three times.

Impact of 2-methoxyestradiol (2-ME) on cell viability and cell death induction of neuroblastoma SH-SY5Y cells. SH-SY5Y cells were treated with a series of selected, representative concentrations: physiological (10−10-10−8 M) and pharmacological (10−6 and 10−5 M) for 24 h. Nitric oxide generation (A) and mitochondrial membrane potential (B) were determined using the Muse Cell Analyzer. Values are the mean±SE of three independent experiments (N=3 replicate cultures). The absence of an error bar denotes a line thickness greater than the error. *p<0.01, **p<0.001, ****p<0.00001 versus control cells (Ctrl). Each experiment was performed at least three times.
Impact of 2-ME on mitochondrial depolarization in neuroblastoma SH-SY5Y cells. Our next goal was to determine if treatment with 2-ME influences the mitochondrial membrane potential. As previously, the cells were treated with physiologically and pharmacologically relevant concentrations of 2-ME for 2 h; subsequently, the changes in mitochondrial potential were determined. As demonstrated in Figure 2B, we noted a significant decrease in mitochondrial membrane potential after 2 h treatment with 10−7, 10−6 and 10−5 M and compared to the control cells, while incubation with 10−8-10−10 M 2-ME did not significantly affect mitochondrial membrane depolarization (Figure 2B).
Discussion
2-ME is an active anticancer agent being evaluated in ongoing clinical trials. 2-ME has been established as affecting cancer cell growth and survival both in vivo and in vitro, as well as leading to inhibition of metastatic processes (breast cancer, pancreatic cancer, Ewing sarcoma, osteosarcoma) (3-5). Serum levels of 2-ME range from 3×10−11 M in men to over 3×10−8 M in pregnant women (2-6). Pharmacological concentrations of 2-ME (10−6 and 10−5 M) inhibited cancer in various experimental models (2-6). We previously demonstrated that 2-ME physiologically inhibits cancer cell growth by inducing DNA damage, and suggested that it can be considered not only as derivative of E2, but as a hormone per se (8).
To our knowledge, there are only few data considering the potential usage of 2-ME in treatment of neuroblastoma (7, 16, 17). Anticancer activity of 2-ME towards neuroblastoma has been shown in several in vitro models (7, 16, 17). Moreover, 2-ME crosses the blood–brain barrier, and is effective in treatment of brain tumors (18). We focused on the involvement of the generation of NO and mitochondrial membrane depolarization in anticancer mechanisms of 2-ME at both physiologically and pharmacologically relevant concentrations. We recently demonstrated apoptosis was induced in highly metastatic osteosarcoma cells at physiological 2-ME concentrations (10−10-10−8 M), while high pharmacologically relevant 2-ME concentrations (10−6 and 10−5 M) induced both apoptosis and necrosis (8). The anticancer effect of 2-ME was strictly associated with generation of NO from neuronal nitric oxide synthase. We suggested that 2-ME may be considered as a natural anticancer agent towards osteosarcoma (8). Herein, we demonstrated that 2-ME can effectively reduce the malignancy of neuroblastoma, another childhood cancer. Neuroblastoma is the most common extracranial solid cancer in childhood and the most common type of cancer in infancy. Prognosis and treatment of neuroblastoma depend on clinical and biological risk factors (14, 15, 24). Surprisingly, 2-ME reduced neuroblastoma cell viability and induced apoptosis only at pharmacological relevant concentrations. Interestingly, 2-ME did not affect necrotic death of neuroblastoma SH-SY5Y cells, while it induced apoptosis only at pharmacologically relevant concentrations. Nonetheless, little information is available concerning the induction of necrosis in cancer cells by 2-ME (8, 19). Lack of toxic effects under physiologically relevant concentrations of 2-ME, as compared to previously used osteosarcoma model (8), may be explained by higher resistance of neuroblastoma to chemotherapy, or activation of another signaling pathways (14, 15, 24).
Manca and Chisu suggested that efficacy of 2-ME in treatment of neuroblastoma may be strictly related to an impairment of microtubule system physiology resulting in morpho-functional changes, block of mitosis and cell death (16). Herein, we suggest that the anticancer activity of 2-ME is correlated with generation of NO and reduction of mitochondrial membrane potential. Previously, it was shown that antioxidants attenuated antiproliferative and apoptotic effects of 2-ME on SK-N-SH and SH-SY5Y cells (7). We also showed that due to induction of neuronal nitric oxide synthase, 2-ME results in cell death of cancer and hippocampal cells (8, 20). An increased level of NO was demonstrated to induce a decrease in the mitochondrial membrane potential of peripheral blood lymphocytes (21). Moreover, it was shown that NO, due to its interaction with components of the electron transport chain, may function as a physiological regulator of cell respiration, and also increase the generation of ROS by mitochondria, thus regulating the mechanisms of cell survival or death (22). Herein, we demonstrated that only at pharmacological concentrations did 2-ME increase NO generation, and reduce mitochondrial membrane potential, resulting in neuroblastoma cell death. Interestingly, physiologically relevant concentrations of 2-ME, although inducing nitro-oxidative stress and even mitochondrial membrane depolarization, did not lead to cell death. Indeed, mitochondrial dysfunction has been shown to participate in the induction of apoptosis. The opening of the mitochondrial permeability transition pore was demonstrated to induce depolarization of the transmembrane potential, in release of apoptogenic factors and the loss of oxidative phosphorylation (25). Moreover, in some apoptotic systems, loss of mitochondrial transmembrane potential may be an early event in the apoptotic process (25). However, mitochondrial depolarization may not always result in induction of cell death due to the various mitochondrial defense systems, including uncoupling proteins (UCP) (26). In particular, uncoupling protein-2 has been shown to protect cancer cells from the cytotoxic actions of chemotherapeutic drugs (26).
Conclusion
These data suggest that regulation of neuroblastoma cell death by 2-ME is dependent on NO generation. 2-ME may be a natural modulator of cancer cell death and survival via nitro-oxidative stress-dependent mechanisms. Interestingly, the role of NO as a modulator of cell death was recently described (23). The potential role of free radicals in regulation of cancer cells and neuronal survival needs to be further evaluated. The obtained data suggest strong anticancer activity of 2-ME and potential therapeutical usage in patients with neuroblastoma.
Acknowledgements
GraphPad Prism 6 for statistical analysis, LAS 500 (GE Healthcare) for Western blot analysis were bought thanks to grant no. 2012/07/B/NZ1/00010 from the National Science Center resources. M.W. acknowledges support from Medical University of Gdansk Funding no. ST46.
The Authors thank Merck Millipore and especially Dr. Barbara Piotrowska for providing Muse Cell Analyzer and helpful advice. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the article.
Footnotes
Author Contributions
Conceived and designed the experiments: Magdalena Gorska; Performed the experiments: Magdalena Gorska; Database collections: Alicja Kuban-Jankowska; Ryszard Milczarek; Analyzed the data: Magdalena Gorska; Manuscript corrections Magdalena Gorska, Michal Wozniak; Wrote the article: Magdalena Gorska.
Conflicts of Interest
The Authors declare no conflict of interest.
Funding
The studies and article publication were by funded grant no. 2012/07/B/NZ1/00010 from the National Science Center resources.
- Received January 5, 2016.
- Revision received February 16, 2016.
- Accepted February 19, 2016.
- Copyright© 2016 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved





{kind=link}
{kind=link}