


Abstract
Aim: The aim of the present study was to detect the relative expressions of p53, p21Waf1/Cip1, p27Kip1 Bcl-2 and cleaved caspase-3 in cervical lesion samples from Brazilian women by immunohistochemistry. Materials and Methods: A total of 230 cervical biopsies in paraffin-embedded blocks were studied: 43 were invasive squamous cell carcinomas (SCC), 52 carcinomas in situ/cervical intraepithelial neoplasias III (CIN III), 54 cervical intraepithelial neoplasias II (CIN II), 51 cervical intraepithelial neoplasias I (CIN I) and 30 non-neoplastic lesions (NN) with benign cellular changes. Results: Significant differences were observed in the p53 expression between the different groups: NN and CIN I (p=0.010); NN and CIN II (p<0.00001); CIN II and CIN III (p=0.02); CIN II and CIS (p=0.0220); CIN II and CEC (p=0.010). Regarding p21WAF1/Cip1, significant differences were observed between NN and CEC (p=0.001); CIN I and CEC (p=0.001); CIN II and CIN III (p=0,001); CIN II and CIS (p=0.0004) and CIN II and CEC (p<0.0001). For p27Kip1, significant differences were observed between NN and CIN I (p<0.00001); NN and CIN II (p<0.00001); NN and CIS (p=0.038); CIN I and CIN III (p=0.001); CIN I and CIS (p=0.009); CIN I and CEC (p=0.0001); CIN II and CIN III (p=0.0003); CIN II and CIS (p=0.002); CIN II and CEC (p< 0.00001). Bcl-2 and caspase-3 did not show remarkable differences between groups. Conclusion: p53, p21WAF1/CIP1, p27KIP1 appear to be involved in the course of carcinogenesis. Rare expression of Bcl-2 and cleaved caspase-3 suggests that these proteins probably do not participate in cervical apoptosis.
Cervical carcinogenesis is considered a multi-factorial and multi-stage process. An early indicator of this process is an increased proliferation of cervical epithelial cells including basal cell hyperplasia, dysplasia, in situ carcinoma and advanced cervical squamous cell carcinoma. In Brazil, 5,160 women died of carcinoma of the uterine cervix in 2011 (1).
Cervical cancers arise through disruption of p53 and pRb pathways by the human papillomavirus (HPV) oncoproteins E6 and E7. Expression of E6 and E7 proteins induces immortalization of cells through their inhibitory effects on proteins pRb and p53, respectively, altering the cell-cycle control and leading to chromosomal instability (2). The HPV oncogenes also affect the expression of other proteins, such as p21Waf1/Cip1 and p27Kip1 either directly or indirectly (3).
p53 is a cell-cycle checkpoint protein that contributes to the preservation of genetic stability by mediating either a G1 arrest or apoptosis in response to DNA damage (4). p53 causes growth arrest through transcriptional activation of the cyclin-dependent kinase inhibitor p21WAF1/Cip1. p27kip1 is also a protein with ability to suppress cell growth. p21Waf1/Cip1 as well as p27Kip1 are members of the Cip/Kip family of cyclin-dependent kinase (CDK) inhibitors and both are targeted by HPV E7 (5). Both proteins affect the activity of kinase complexes controlling the G1-S transition and are considered putative tumor suppressors (6). Therefore, wild-type p53, p21Waf1/Cip1 and p27Kip1 play an important role in G1-S transition of the cell cycle.
Mutations in the TP53 contribute to the development of up to 50% of all human cancers. p53 overexpression was detected in 50% of human papillomavirus-associated cervical carcinoma with a single mutation in exon 7 (7). Akasofu and Oda (8) did not detected overexpression of p53 in the normal, CIN I and CIN II. High-level of p53 accumulation was identified in basal and supra-basal atypical cells in 27.0% of CIN III and in 43.5% of invasive squamous cell carcinoma. Overexpression of p53 was found to co-exist with infection by HPV in 17 of 37 CIN III and 12 of 23 invasive squamous cell carcinomas. The authors suggested that inactivation of the p53 function by HPV infection or alteration of p53 protein itself precedes the development of tumors with a fully-malignant and invasive phenotype and plays an important role in tumorigenesis in the uterine cervix. Lakshmi et al. observed increased expression of p53 in dysplastic and malignant lesions compared to non dysplastic lesions and suggested that p53 protein accumulation may be an early event in carcinogenesis (9).

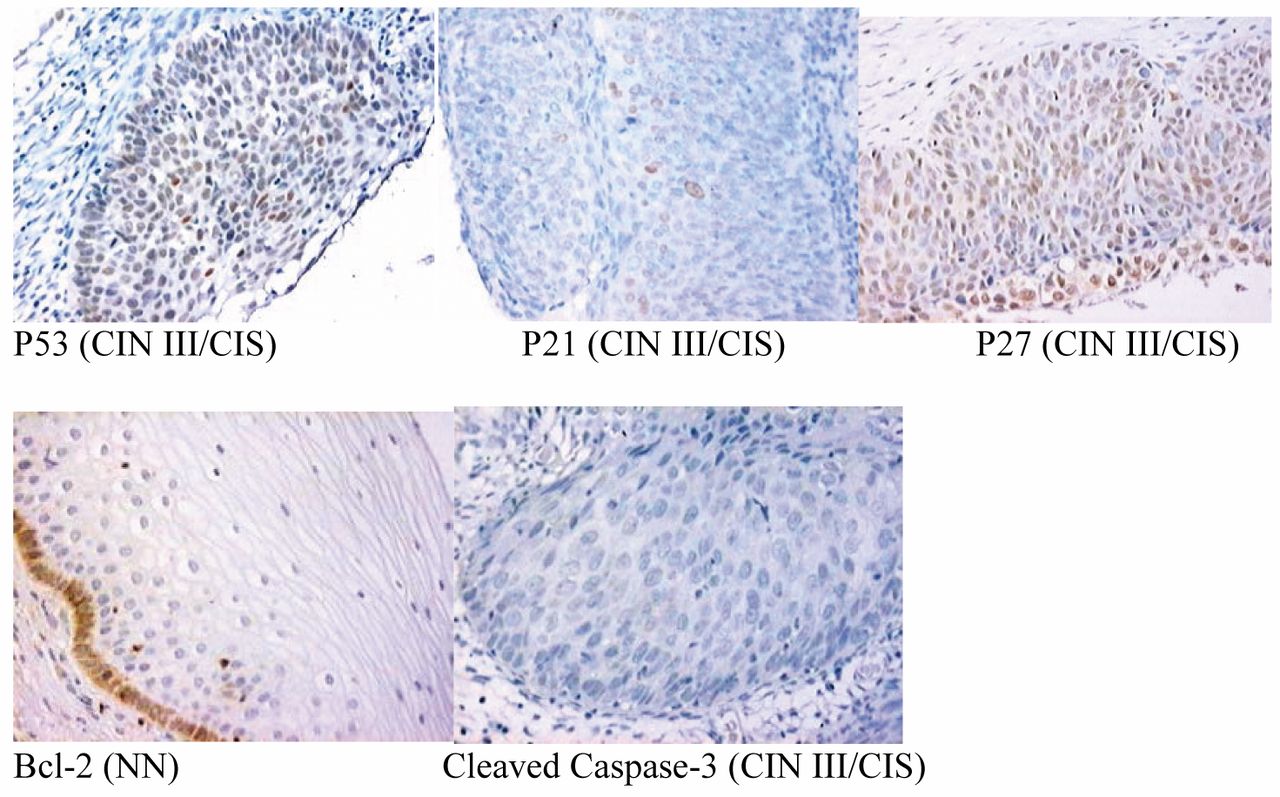
Expression of p53, p21WAF1/Cip1, p27Kip1, Bcl-2 and cleaved caspase 3 in cervical lesions. Nuclear positivity for p53, p21WAF1/Cip1, p27Kip1 Bcl-2 was observed in the cytoplasm of cervical cells and cleaved caspase-3 was negative for staining. (Original magnification, ×100).
Studies of expression of p21Waf1/Cip1 and p27Kip1 in cervical cancers have produced inconsistent results (10). A significant increase in p21Waf1/Cip1 and p53 expression was found in normal cervix by El-Ghobashy et al. (11). Moreover, the expression of p21Waf1/Cip1 was seen markedly increased in high grade lesions by Santopietro et al. (12).
Bcl-2 protein is a mitochondrial membrane protein which protects cells from programmed cell death. Intrinsic apoptosis pathway (mitochondrial mechanism) controlled by Bcl-2 family promotes activation of proteolytic enzymes cascade (13). The principal intracellular effectors of apoptosis are a family of cysteine proteases. Caspase-3 represents a prominent member of this family of proteases which operates in the distal portion of convergent proteolytic cascades, serving as an effector of apoptosis by cleaving critical protein substrates required for apoptotic cell death.
Zanotti et al. (14) reported that Bcl-2 immunoexpression was restricted to the basal cell layer in all CIN stages as in normal cervical epithelium. Active caspase-3 was present in the supra-basal layer and extended to all upper layers in normal epithelium and slightly decreased with increasing dysplasia. In invasive carcinoma, it was restricted to few scattered cells. Walker et al. demonstrated that active caspase-3 was expressed in the normal mucosa and CINs from HIV-negative patients but in rare to few cells scattered in the mucosa (15). The number of immunopositive cells was enhanced in the CINs from HIV-positive patients by comparison with those from HIV-negative ones, especially in high-grade lesions.
The role of apoptosis-associated proteins in cervical carcinogenesis is unclear. In the present study, the relative expressions of p53, p21Waf1/Cip1, p27Kip1 Bcl-2 and cleaved caspase-3 were examined in cervical lesions from Brazilian women.
Materials and Methods
Retrospectively, a total of 230 cervical biopsies in paraffin-embedded blocks from the archive of Department of Pathology, Federal University of São Paulo and Laboratory of Anatomopathological analysis of Avaré (Sao Paulo State, Brazil) were analyzed. The biopsies were collected during ambulatorial and surgery procedures to pathological analyses. Histological preparations of all cases were reviewed by two investigators (GRAF and FOL), and a consensus diagnosis was reached. All samples were histologically-graded by Richart and Bethesda systems as 43 invasive squamous cell carcinomas (SCC), 52 carcinoma in situ/cervical intraepithelial neoplasia III (CIN III), 54 cervical intraepithelial neoplasia II (CIN II), 51 cervical intraepithelial neoplasia I (CIN I) and 30 non-neoplastic lesions (NN) with benign cellular changes. Ethical approval for the study was granted by the local Ethics Committee (The Resolution n° 196 of National Health Council).
Frequency (total number of cases) of p53, p21WAF1/cip1, p27kip1 and Bcl2 immunoexpression in different cervical lesions.
Immunohistochemistry. Conventional 5-μm thick sections were obtained and mounted on slides pre-treated with 3-minopropyl-triethoxysilane (Sigma). Sections were then de-paraffinized, hydrated and processed as follows. For antigen retrieval slides were placed in 0,01 M citrate-buffer pH 6,0 and heated in a steamer for 30 min. Endogenous peroxidase activity was blocked by using 10% hydrogen peroxide for 20 min. The sections were incubated with the monoclonal mouse anti-human p53 protein (clone DO7-Dako, Copenhagen, Denmark) (1:100), monoclonal mouse anti-human p21wafi/Cip1 protein (Clone SX118-Dako, Copenhagen, Denmark) (1:50), monoclonal mouse anti-human p27kip1 protein (Clone SX53G8-Dako) (1:100), monoclonal anti-human Bcl2 protein (Dako) (1:80) and monoclonal anti-Cleaved Caspase-3 (Cell Signaling, Boston, Massachusetts, USA) (1:300) overnight in a humid chamber. After washing with PBS, sections were allowed to react with a secondary biotinylating antibody and streptavidin-biotin-peroxidase (LSAB, Dako) for 30 minutes each one. Finally, the reaction was revealed using 3, 3’-Diaminobenzidine tetrahydrocloride (Sigma) (0,07 diaminobenzidine in PBS buffer 0,05 M, pH 7,6, 0,01% hydrogen peroxide) counterstained with Harris's hematoxylin and coverslipped with Entellan (Sigma). Negative and positive controls were ran simultaneously. Positive control was represented by breast carcinoma tissue. Negative controls were prepared by eliminating the primary antibody.
Data analysis. The p53, p21WAF1/Cip1, p27Kip1 and cleaved caspase 3 proteins were located in the nucleus of the tumor cells, and the staining ability was analyzed based on the estimation of the percentage of positive tumor cell. p53, p21Waf1/Cip1 and cleaved caspase-3 were considered positive when ≥10% of nuclei of tumor cells presented strong-positive staining and p27Kip1 when >40% of nuclei of tumor cells presented strong positive stain (16). The positive pattern of Bcl-2 was the cytoplasmic type. This pattern was analyzed according the estimation of the percentage of staining (17).
Statistical analysis. In order to determine the possible statistical differences, we compare the results of each protein using the χ2 test with correction of Yates. All the pairs were compared through of multiple comparisons of Dunn. The results were considered significant for a value of p<0.05.
Results
Nuclear staining was considered positive for p53, p21Waf1/Cip1, p27Kip1 and cleaved caspase-3. Cytoplasm staining was considered positive to Bcl-2. Representative staining for all markers are shown in Figure 1. No nuclear or cytoplasmic staining was observed in the negative controls, as well as no staining of deep in the samples and in the controls.
The intensity and distribution of p53 staining in tumor cells were variable. We verify the expression of p53 in 32 (13.91%) samples (Table I). In NN and CIN I the expression of p53 occurred more frequently in the basal and parabasal cells layers. Rare imunoexpression was observed in CIN I and CIN II. Significant differences were observed in the p53 expression between the different groups: NN and CIN I (p=0.010); NN and CIN II (p<0.00001); CIN II and CIN III (p=0.02); CIN II and CIS (p=0.0220); CIN II and CEC (p=0.010) (Table II).
Nuclear expression of p21WAF1/cip1 was observed in 60 (26.09%) samples mainly in CIN III/CIS and CEC (Table I). Significant differences were observed between NN and CEC (p=0.001); CIN I and CEC (p=0.001); CIN II and CIN III (p=0.001); CIN II and CIS (p=0.0004) and CIN II and CEC (p< 0.0001) (Table II).
The expression of p27Kip1 was observed in 79 (34,35%) samples (Table I). p27Kip1 was found positive in the cells of stroma (fibroblasts), the glands and the vases. Cells of basal and parabasal layers showed increase of the intensity in agreement with cellular maturation in NN. Significant differences were observed between NN and CIN I (p< 0.00001); NN and CIN II (p< 0.00001); NN and CIS (p=0.038); CIN I and CIN III (p=0.001); CIN I and CIS (p=0.009); CIN I and CEC (p=0.0001); CIN II and CIN III (p=0.0003); CIN II and CIS (p=0.002); CIN II and CEC (p<0.00001) (Table II).
Cytoplasmic expression of Bcl2 was distributed in the cells of basal and parabasal layers of NN and CIN I. Positive staining of polimorfonuclear cells was considered as internal positive control. Five samples presented staining for Bcl2 (one case (3.33%) of NN, two cases (3.92%) of CIN I and two cases (4.65%) of CEC).
Comparisons of different cervical lesions relative to p53, p21Waf1/Cip1, p27Kip1 and Bcl-2 expressions (χ2 test with correction of Yates).
The presence of cleaved caspase-3 protein was rare and weak in the all groups. Polimorfonuclear cells located in the stroma, as well as in other cells in process of apoptosis presented cytoplasmic staining. Cleaved caspase-3 protein was considered negative for all groups.
Tables II and III showed the results of the comparisons of different cervical lesions relative to p53, p21Waf1/Cip1, p27Kip1 and Bcl-2 expressions. The comparisons of p53, p21Waf1/Cip1, p27Kip1 and Bcl-2 expressions relative to different cervical lesions (χ2 test with correction of Yates), respectively.
Discussion
The process of carcinogenesis at the cellular level is related to disorders of the control of cell proliferation, differentiation and controlled cell death (apoptosis). Therefore, the present study aimed to investigate the role of a panel of cell cycle and apoptosis associated-proteins in cervical lesions (NN until CEC) of Brazilian women. More recently Hellberg and Tot published “Tumor Marker Score for Prognostication of Early-stage Squamous Cell Cervical Cancer” and among the markers are cited p53 and p27 (18).
The relationship between p53 protein abnormalities and cancer has been well-documented. The detection of p53 protein in cervical cancers by immunohistochemistry has been attributed to the use of antibodies which detect de-stabilization of wild-type protein resulting from ongoing stress or disruption to proteins involved in p53 degradation in addition to mutations (19). Wild-type p53 protein has a very short half-life and thus is present in only minute amounts (20), generally below the detection level of immunocytochemical methods (21). Furthermore, mutant p53 proteins often are much more stable than wild-type p53 protein; therefore, the mutant p53 protein accumulates to a high level (20). The immunostaining of p53 can occur to as result of its stabilization by interaction with the E6 and E7 protein of the HPV or when complex with protein as the Hsp27 (22).
Comparisons of p53, p21Waf1/Cip1, p27Kip1 and Bcl-2 expressions relative to different cervical lesions.
In this study, we used the monoclonal antibody anti-human p53 (clone DO7) that labels wild-type and mutant-type p53 protein and is used for the identification of p53 accumulation in human neoplasias (23). High expression of mutated p53 was observed in NN in relation to CIN I and CIN II, mainly in the basal and suprabasal cell layer, just as described for Holm et al. (24). Perhaps this immunostaining can be explained by the activation of the wild type p53 to arrest the cell-cycle by cell injuries. The majority of CIN I and CIN II were negative for p53 followed of CIN III/CIS and CEC. This pattern also was described for Holm et al. that found few positive neoplasic cells, results related to the high frequency of HPV infection (24).
Expression of p53 protein was detected in 42% of cervical cytologies without squamous intra-epithelial lesions and 33.3% of low-grade squamous intra-epithelial lesions (CIN I). On the other hand, the authors did not find signifi¬cant differences in p21 expression between the two lesions (25). In a previous study, Graspa et al. analyzed the expression of p53 in normal cervical tissue and low grade squamous intraepithelial lesions (CIN I), however, they did not detect p53 expression in either cervical cytologies without squamous intraepithelial lesions or low grade squamous intraepithelial lesions (26).
Amaro-Filho et al. in a comparative analysis of clinical and molecular factors and stage of cervical cancer in a Brazilian cohort found positive expression of p53 in 49.4% of cervical cancer (27). In contrary, our population of cervical cancer (CIN III/CIS/CEC) was positive in only 17%. The expression of p53 was increased in CIN III/CIS and CEC in relation to CIN II. Park et al. also observed an increased expression of p53 in more advanced stages of cervical cancer (28). Moreover, all samples contained the HPV16 integration, demonstrating that HPV infection and alterations of the TP53 gene have the role in cervical carcinogenesis. Bahnassy et. al (29) analyzing the possible role of cell-cycle regulators in multistep process of HPV-associated cervical carcinoma concluded that p53 pathway abnormalities was late event. On the other hand, Chetty observed that the expression of p53 protein was independent of the HPV 16/18 status of the tumor (23).
In the present study, we did not search the presence of HPV infection. However, epidemiological evidence, clinical investigation, serology blood tests and molecular biology studies, indicate that the HPV infection corresponds to the main etiological factor of cervical carcinogenesis (30). Moreover, about 90% of the cervical cancers presents high oncogenic risk HPV, mainly types 16 and 18 (31).
The loss of TP53 function caused by mutations is the main factor that characterizes the human cancers (32). Several independent studies have shown that in a small percentage of cervical tumors the TP53 is mutated. However, this event appears to be unrelated to the presence or absence of HPV infection and the nature of the tumor (33). However, the relation between HPV infection and the state of TP53 is unclear. Moreover, the frequency of mutations in this gene is rare in patients without HPV infection (34). Expression of p53 like a pro-apoptotic protein is noted in cervical cancer irrespective of expression of HPV oncogenes (35) suggesting that additional mechanisms are operating in the de-regulation of this pathway.
Cells with wild-type p53 that express p21WAF1/Cip1 either undergo apoptosis or repair their DNA. Functional inactivation of TP53 by mutation can occur in various human carcinomas blocking the cancer cells to express the p21WAF1/Cip1. As a result, the cancer cells cannot proceed to apoptosis and the tumor continues to grow (36). In tumor cells, p21WAF1/cip1 expression results in inhibition of proliferation (37), being waited inverse correlation between p21WAF1/cip1 expression and tumor development.
Studies demonstrate that the linking between E7 protein and p21WAF1/cip1 blocks the ability of p21WAF1/cip1 arrest the cell cycle between S and M phases (38). This suggests the involvement of other proteins, as the E6. E6 protein binds the tumor suppressor p53 and promotes its degradation through an ubiquitin-dependent pathway taken to the reduction of p21WAF1/cip1 gene regulation (39).
p21WAF1/cip1 was observed in NN and CIN I. p21WAF1/cip1 expression was significantly higher in CEC comparing to NN according to Horikoshi et al. (40). Increased p21 was detected in CINIII and invasive squamous cell carcinoma (ISCC) by Bahnassy (7). Santopietro et al. found high level of p21WAF1/cip1 in CIN III and CEC (12). In our study, there were correlation between CIN III/CIS and CEC as well as significant differences were observed correlating p21WAF1/cip1 with NN and CEC, CIN I and CEC, CIN II and CIN III/CIS as CIN II and CEC.
Lie et al. demonstrated that higher expression of p21WAF1/cip1 in the pre-cancerous injuries and the CIS is independent of the transcription way of p53 (41). Studies show that the p21WAF1/cip1 is regulated by two ways, independent and dependent of p53 expression (42). Unlike to its tumor suppressor function p21WAF1/cip1 has the ability to protect the cells of apoptosis acting as oncogene. Therefore, the agreement of the action of p21WAF1/cip1 in the cervical cancer is important for the implementation of alternative treatments that promote sensitivity to apt the tumor cells becoming them to suffer apoptosis induced for chemotherapy drugs.
The p27KIP1 cyclin-dependent kinase inhibitor displays apparently contradicting roles by acting as a classic tumor suppressor in one instance, and a pro-tumorigenic oncogene in others (43). Mutations or deletion of p27kip1 gene is rare in human cancers, and reduction or absence of p27kip1 expression is associated with highly aggressive phenotype of cervical carcinoma. (44). E7 can antagonize the ability of p27KIP1 to block cyclin E-associated kinase in vitro (45).
Huang et al. investigated the relationship between different human papillomavirus (HPV) genotypes and the expression of p53, p21 and p27 in cervical carcinomas (46). The results showed that the expression of p53 and p27 was not related to HPV genotype but in the HPV-18 positive cervical carcinomas, expression of p21 was significantly decreased or completely absent.
In the present analysis, we found nuclear expression of p27kip1 in the basal and supra-basal cells, where it has cellular response epithelium and stroma of NN (47), and strong nuclear staining of the limphocytes which we consider internal control (48). We did not observe cytoplasmic staining of p27kip1 unlike other studies that associated this staining in different cancers with tumor differentiation in advanced periods of training of the illness (49).
In the present study, differences were observed between NN and CIN I, NN and CIN II, NN and CIS and the groups with the most advanced injuries (CIN I and CIN III/CIS, CIN I and CEC, CIN II and CIN III/CIS, NIC II and CEC). This increase is associated the favorable prognostic factor. The reduction of p27kip1 in CIN I and CIN II would be explained by increased cellular proliferation in the early stage of tumor progression. Reports on the changes in the expression patterns of p21 and p27 in cervical carcinoma progression are contradictory (10).
The Bcl2 protein inhibits the process of apoptosis and draws out the cellular survival. Studies have shown that progression of neoplasia in the uterine cervix is accompanied by an increase of Bcl-2 as well as cellular proliferation (50). Manusirivithaya et al. in 2006 described Bcl-2 expression in initial cervical neoplasic injuries (51). In contrast, Bcl-2 was negative in the majority of our samples (NN to the CEC). In samples of groups NN and CIN I we observed weak expression of Bcl-2 in the basal cells layers, as described in literature (52) and very weak staining in the neoplasic cells. On the other hand, we observed Bcl-2 expression in the lymphocytes, that we considered a internal positive control, validating the reaction beyond of the external positive control. The loss of Bcl-2 was reported previously to be a poor prognostic marker for cervical carcinoma (53).
Caspase-3 is the most important member of the caspase family and is widely expressed in normal human tissues and various tumor tissues. The majority of caspase family members are promoters or effectors of apoptosis, playing important roles in the process of apoptosis. Activated caspase-3 is responsible for the cleavage of poly (ADP-ribose) polymerase (PARP), actin and sterol regulatory element binding protein (SREBP), which are associated with apoptosis (54). After high risk-HPV infection, viral proteins employ strategies to modulate apoptosis. The E2 viral protein induces apoptosis in both normal and HPV-transformed cells through activation of caspase-8. E6 inhibits apoptosis through the proteolytic inactivation of pro-apoptotic proteins such as procaspase-8 (55).
In the present study, we analyzed the expression of cleaved caspase-3. The loss of expression of cleaved caspase-3 was observed since the initial injuries to the cervical carcinoma but some polymorphonuclear cells were positive for this protein. It was considered an internal positive control validating the reaction in set with the external positive controls.
Lu et al. studied expression of survivin, caspase-3 and p53 in cervical cancer assessed by tissue microarrays and suggested that these proteins may play an important role in the occurrence and development of cervix carcinoma (56). Particularly, low expression of caspase-3 is closely correlated with cervical cancer. Cheung et al. observed an increase in expression of Caspase 3 in CIN-H compared to CIN-L and suggested that this increase to be related to the increased proliferative activity in dysplastic cells (57). On the other hand, the reduction of Bax, Caspase 3, and Caspase 6 expression found in carcinoma to indicate that the apoptotic mechanism has become defective in the process of malignant transformation.
Compared to previous reports our study included small number of samples; therefore, it is necessary to validate our results in a larger population. Taken together, these results indicate that p53, p21WAF1/CIP1, p27KIP1 appear to be involved in the course of carcinogenesis. Rare expression of Bcl-2 and cleaved caspase-3 suggests that these proteins probably do not participate in cervical apoptosis.
Footnotes
-
Conflicts of Interest
None declared.
- Received February 22, 2014.
- Revision received April 22, 2014.
- Accepted April 23, 2014.
- Copyright© 2014 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.