


Abstract
Background/Aim: Sonodynamic cancer therapy is based on the preferential uptake and/or retention of a sonosensitizing drug (sonosensitizer) in tumor tissues and subsequent activation of the drug by ultrasound irradiation. In the present study, we investigated the participation of lipid peroxidation in the mechanism of the sonodynamically-induced antitumor effect with functionalized fullerenes, such as polyhydroxy fullerene (PHF. Materials and Methods: Ultrasonically-induced cell damage and lipid peroxidation with PHF were compared in the same in vitro insonation setup. Sarcoma 180 cells suspended in PBS were exposed to 2 MHz ultrasound in the presence and absence of PHF. Cell viability was determined by the Trypan Blue exclusion test. Lipid peroxidation in cell membranes was estimated by measuring the amount of malondialdehyde as the thiobarbituric acid-reactive-substances. Results: Significant enhancement of the rates of both ultrasonically-induced cell damage and lipid peroxidation was observed in the presence of PHF, both of which were positively correlated with PHF. The enhancement of cell damage and lipid peroxidation with PHF was suppressed by reactive oxygen scavengers such as histidine and tryptophan. Conclusion: The good correlation observed in the presence of PHF suggests that membrane lipid peroxidation is one of the important intermediary events in sonodynamically-induced cellular damage. The inhibitory effects of histidine and tryptophan also provide evidence that singlet oxygen plays an important role in PHF-mediated sonosensitization of membranes and that this moiety may be an important mediator of cell destruction in sonodynamic therapy associated with PHF and ultrasound.
- Ultrasound
- nanomedicine
- fullerenes
- sonodynamic therapy
- membrane lipid peroxidation
- sarcoma 180
- reactive oxygen species
Nanomedicine is the medical application of nanotechnology for the diagnosis and treatment of human diseases. It is defined as the study and use of structures with sizes that vary between 1 and 100 nm. Nanomaterials such as functionalized fullerenes have unique physicochemical properties, such as small size, large surface area mass ratio and high reactivity, which are used to overcome some limitations of traditional therapeutic agents (1-3). One of the most important features of nanomaterials is the potential for improved drug delivery to the target area, thereby maximizing the therapeutic efficacy (4-6). Due to their size, nanomaterials are capable of accumulating in malignant areas such as solid tumors (5).
Ultrasound has an appropriate tissue attenuation coefficient for penetrating intervening tissues to reach non-superficial objects while maintaining the ability to focus energy into small volumes. This is a unique advantage when compared to electromagnetic modalities, such as laser beams, in the application to non-invasive treatment of non-superficial tumors. Although the use of ultrasound for tumor treatment has been relatively well-investigated with respect to the thermal effects associated with ultrasound absorption, only few studies have reported on non-thermal effects, such as the sonochemical effects of ultrasound cavitation (7).
Recently, we found that photochemically-active porphyrins, such as hematoporphyrin and porfimer sodium, can induce significant cell damage when activated by ultrasound. When implanted murine tumors are treated with ultrasound after the administration of such chemicals, tumor growth is significantly inhibited compared to ultrasound-alone. Therefore, photochemically-active porphyrins may be useful for sensitizing tumors to ultrasound. We proposed that this potential modality be called ‘sonodynamic therapy’ (8-21).
In recent years, water-soluble functionalized fullerenes, such as polyhydroxy fullerenes (PHF, Figure 1) have received considerable attention specifically due to their interesting sensitizing properties, which have been exploited in many biological fields. For example, a potential biological application of functionalized fullerenes is related to their photosensitization by either UV or visible light (22, 23). The photochemically-mediated cytotoxicity of water-soluble functionalized fullerenes is induced through photo-oxidation reactions initially involving the excitation of a porphyrin molecule by light. The excited photosensitizer may accept or lose an electron, leading to free radical reactions (Type I) or may transfer energy to oxygen-producing singlet oxygen (Type II) (24).
Yumita et al. have investigated the sonodynamically-induced antitumor effect of PHF (25). The cell damage induced by sonication was significantly enhanced in the presence of PHF. Histidine significantly inhibited this enhancement. This inhibitory effect suggests that the sonodynamically-induced antitumor effect was mediated by sonodynamically-generated reactive oxygen species. This hypothesis is similar to the above-described Type II mechanism of photo-induced cytotoxicity. It is believed that lipid-rich cell membranes are a potential target for oxidative cell damage by active oxygen species (26). Therefore, it would be of interest to know whether PHF enhances ultrasonically-induced oxidative damage to cell membranes.
In this study, the sonodynamically-induced damage and lipid peroxidation of cell membrane with PHF was investigated. These were compared under the same ultrasonic exposure condition.
Materials and Methods
Chemicals. Polyhydroxy small gap hydrated fullerenes (PHF), histidine, tryptophan, mannitol and superoxide dismutase (SOD) were purchased from Sigma-Aldrich (St. Louis, MO, USA). Thiobarbituric acid (TBA) and malondialdehyde (MDA) were purchased from Wako Junyaku Kogyo (Tokyo, Japan). All the other reagents were commercial products of analytical grade.
Tumor cells. The sarcoma 180 cell line was supplied by Meiji Seika Kaisha Co. Ltd., (Tokyo, Japan). The cells were passaged weekly through male ICR mice (5 weeks old) in the form of ascites. Sarcoma 180 cells harvested from the peritoneal cavity of a tumor-bearing animal 7 to 10 days after inoculation were suspended in an air-saturated phosphate buffer saline solution (PBS, pH 7.4) at a concentration of 5×106 cells/ml. The integrity of the cells was confirmed before every treatment and cell suspensions with integrity above 99% were used in both ultrasonically-induced lipid peroxidation and cell damage experiments (16, 27).
Insonation apparatus. An ultrasound transducer of basically the same design as in the previous studies (16, 27) with a resonance frequency of 2 MHz was used for all the insonation experiments. Sine waves were generated by a wave generator (model MG442A; Anritsu, Tokyo, Japan) and amplified by a radio-frequency (RF) amplifier (model 210L; ENI, Rochester, NY, USA). The air-backed transducer uses a lead-titanate piezoelectric ceramic disk of 24 mm diameter, which was purchased from Hitachi Metals Ltd. (Tokyo, Japan) and attached to the aluminum layer of the transducer with a low heat-expansion epoxy adhesive. The experimental set-up is shown in Figure 2. The transducer was submerged in degassed water at room temperature facing upward with its acoustic surface parallel to the water surface. An insonation glass container of 31 mm diameter with a flat bottom layer of 1.5 mm thickness was placed 30 mm from the transducer. A 2.5 ml aliquot of air-saturated sample suspension or solution was placed in the container. The temperature rise in 2.5 ml air-saturated water in the glass container during the insonation was confirmed using a 0.25 mm diameter Chromel-Almel thermocouple. The increase was less than 1°C and 10°C in the insonation for 1 min and 15 min, which were the maximum insonation lengths for the experiments with cells and for the lipid peroxidation, respectively.

The chemical structure of PHF.
Cell damage detection. Ultrasonically-induced cell damage in the presence and absence of PHF in suspensions with and without active oxygen scavengers was detected by Trypan Blue staining of the isolated cells. A 0.2-ml aliquot was taken from the cell suspension immediately after a fixed duration of insonation and mixed with 0.2 ml of 0.5% Trypan Blue solution. The integrity of the cells was determined by counting the number of unstained cells on a hemocytometer glass plate using an optical microscope. Each result is presented as the mean with standard deviation of four experiments. The ratio of unstained cells was fitted using a least squares method to a monoexponential function of ultrasonic exposure time and the cell damage rate was calculated from the coefficient of the function (16).
Determination of lipid peroxidation. Lipid peroxidation of cell membranes was estimated using the thiobarbituric acid (TBA) method (16, 27). It is known that this method is not specific to a particular kind of peroxide and that lipid peroxides such as endoperoxides and hydroperoxides can be detected. Lipid peroxidation was estimated immediately after ultrasonic exposure unless otherwise noted. To each 0.5-ml of insonated cells, 4 ml of 0.083 N H2SO4 and 0.5 ml of 10 % sodium phosphotungstate were added. After centrifugation at 1500 ×g for 10 min, the precipitate was discarded and 4 ml of H2O and 1 ml of 0.67% TBA were added. Samples were incubated for 60 min at 95°C in a boiling water bath and then cooled. Subsequently, n-Butanol (4 ml) was added and mixed vigorously, and the butanol phases were separated by centrifugation. The fluorescence intensity was measured at excitation and emission wavelengths of 515 and 535 nm, respectively. Values were converted to the amount of TBA reactive substance (TBARS) by scaling to an external standard of malondialdehyde (MDA) solutions diluted with PBS. Lipid peroxidation was expressed in terms of the amount of produced TBARS per mg of protein in the cell suspension. The lipid peroxidation rate was obtained by fitting a straight line using a least squares method to the amount of TBARS plotted against insonation time (16, 27).

Diagram of the insonation set-up.
Results
Cell damage. The unstained fractions of isolated sarcoma 180 cells in the air-saturated suspensions, after fixed duration of insonation at an ultrasound intensity of 6 W/cm2, are shown in Figure 3. Results were plotted versus insonation time for PHF concentrations of 0, 20, 40, 60 and 80 μM. Data obtained with 80 μM PHF without ultrasound were also plotted versus time. The unstained fractions on a logarithmic scale decreased linearly with insonation time. A straight line, obtained by fitting a monoexponential curve using a least squares method to the unstained fraction on a linear scale, was drawn for each plot. The cell damage rate corresponding to each line was plotted against PHF concentration in Figure 5. The ultrasonically-induced cell damage rate increased as PHF concentration increased. PHF at a concentration of 80 μM enhanced the rate by more than three-fold. No cell damage was observed with PHF-alone.
Peroxidation. The ultrasonic peroxidation yield in the presence and absence of PHF and under the same acoustic condition, as the cell damage experiment, was plotted against duration time in Figure 4. Peroxidation yield was estimated in terms of the amount of produced TBARS and plotted versus insonation time for PHF concentrations of 0, 20, 40, 60 and 80 μM. The data obtained with 80 μM PHF without ultrasound was also plotted versus time. The peroxidation yield increased linearly with ultrasonic exposure time. A straight line, fitted using a least-squares method, was drawn for each plot. The peroxidation rate corresponding to each line was also plotted against PHF concentration in Figure 5. The peroxidation rate increased as PHF concentration increased. PHF at a concentration of 80 μM enhanced the rate by approximately three-fold. No peroxidation was observed with PHF-alone.
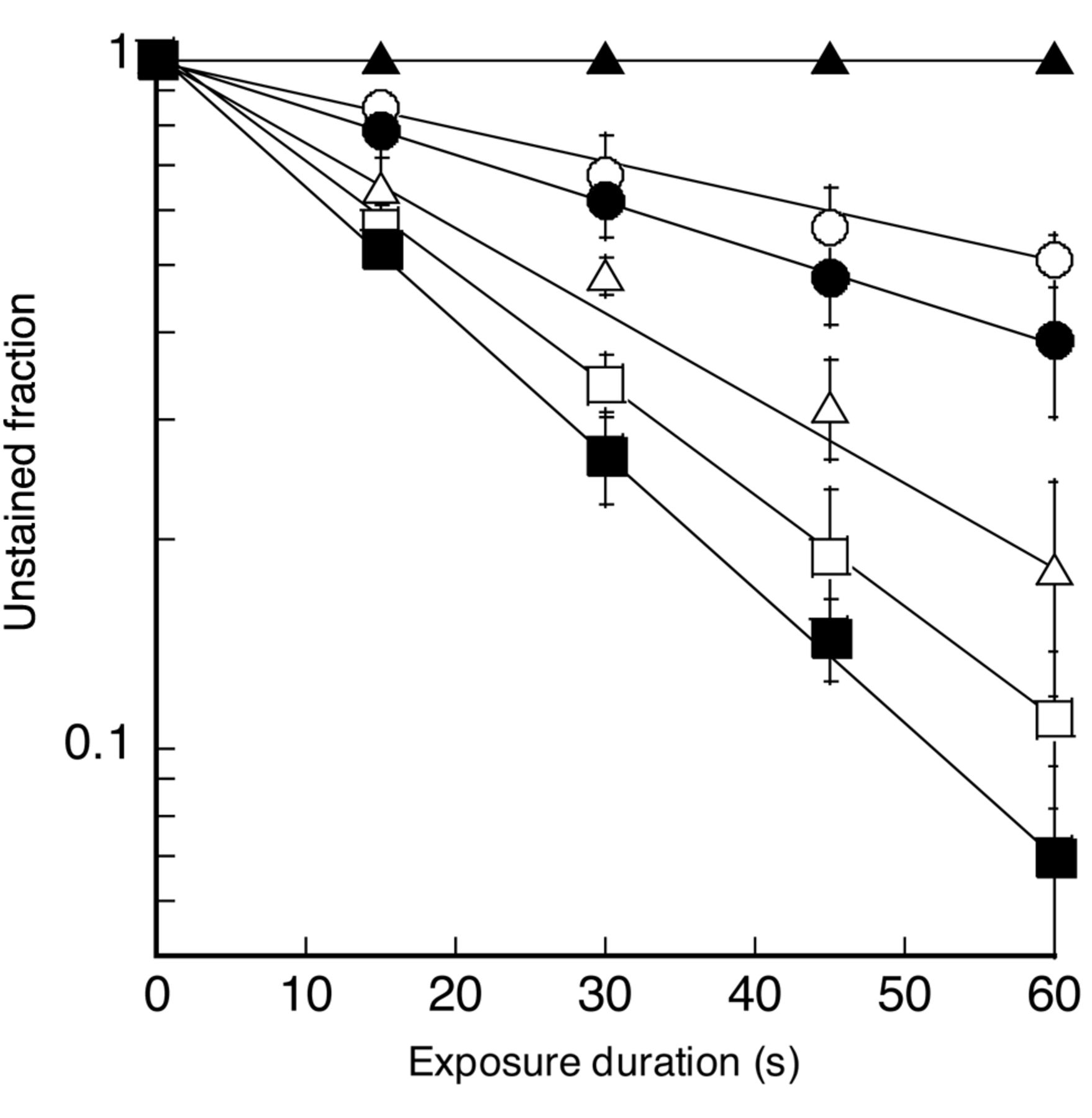
Effect of PHF and/or ultrasound on isolated sarcoma 180 cell damage. ▴, 80 μM PHF alone; ○, ultrasound alone; ●, 20 μM PHF + ultrasound; ▵, 40 μM PHF + ultrasound; □, 60 μM PHF + ultrasound; ▪, 80 μM PHF + ultrasound. Each point and vertical bar represent the mean±S.D. of four insonation experiments.

Effect of PHF and/or ultrasound on membrane lipid peroxidation. ▴, 80 μM PHF alone; ○, ultrasound alone; ●, 20 μM PHF + ultrasound; ▵, 40 μM PHF + ultrasound; □, 60 μM PHF + ultrasound; ▪, 80 μM PHF + ultrasound. Each point and vertical bar represent the mean±S.D. of four insonation experiments.

The relationship between the cell damage rate and the peroxidation rate, obtained at the same PHF concentration under the same acoustic condition, is shown in Figure 6. Both rates correlate well showing a correlation coefficient of 0.978.
Effect of active oxygen scavengers. The effect of active oxygen scavengers on both ultrasonically-induced in vitro cell damage and peroxidation in the presence and absence of 80 μM PHF was tested and the results are shown in Figures 7 and 8. The effects of 10 mM histidine, 10 mM tryptophan, 100 μg/ml SOD and 100 mM mannitol on the unstained cell fractions after 60 s of insonation are compared in Figure 7 and their effects on the peroxidation yield during 60 s of insonation are displayed in Figure 8. Both the ultrasonically-induced cell damage and peroxidation yield, which were enhanced by PHF, were significantly reduced in the presence of 10 mM histidine or 10 mM tryptophan but not in the presence of either 100 μg/ml SOD or 100 mM mannitol. None of these scavengers showed any significant effect when ultrasound was used without PHF.

Relationship between cell damage rate and peroxidation rate. The transducer was driven at a voltage corresponding to the free-field ultrasonic intensity of 6 W/cm2. Each point and vertical bar represent the mean±S.D. of four insonation experiments. A straight line was fitted using a least squares method to plotted points for PHF concentration. Symbols are the same as in Figures 3 and 4.
Discussion
The destruction of tumor tissue during sonodynamic therapy has been shown to be caused by ROS generated by the sonosensitization of porphyrins. Among the ROS generated, singlet oxygen has been identified as one of the major species responsible for tumor cytotoxicity (12, 16, 21, 25). Membrane damage is considered a crucial event contributing to sensitizer cytotoxicity (26). Recent spin trapping studies have also suggested that ROS target membrane lipids (27). One of the major processes initiated is lipid peroxidation. There is considerable evidence suggesting that cell membranes are important sites of ultrasound sensitization and the peroxidation of cell membranes is linked to a decrease in cell viability (27, 28). In view of the critical role of cell membranes in sonodynamically-induced cell damage, we examined sonodynamically-induced lipid peroxidation with PHF in cell membranes in comparison to cell damage.
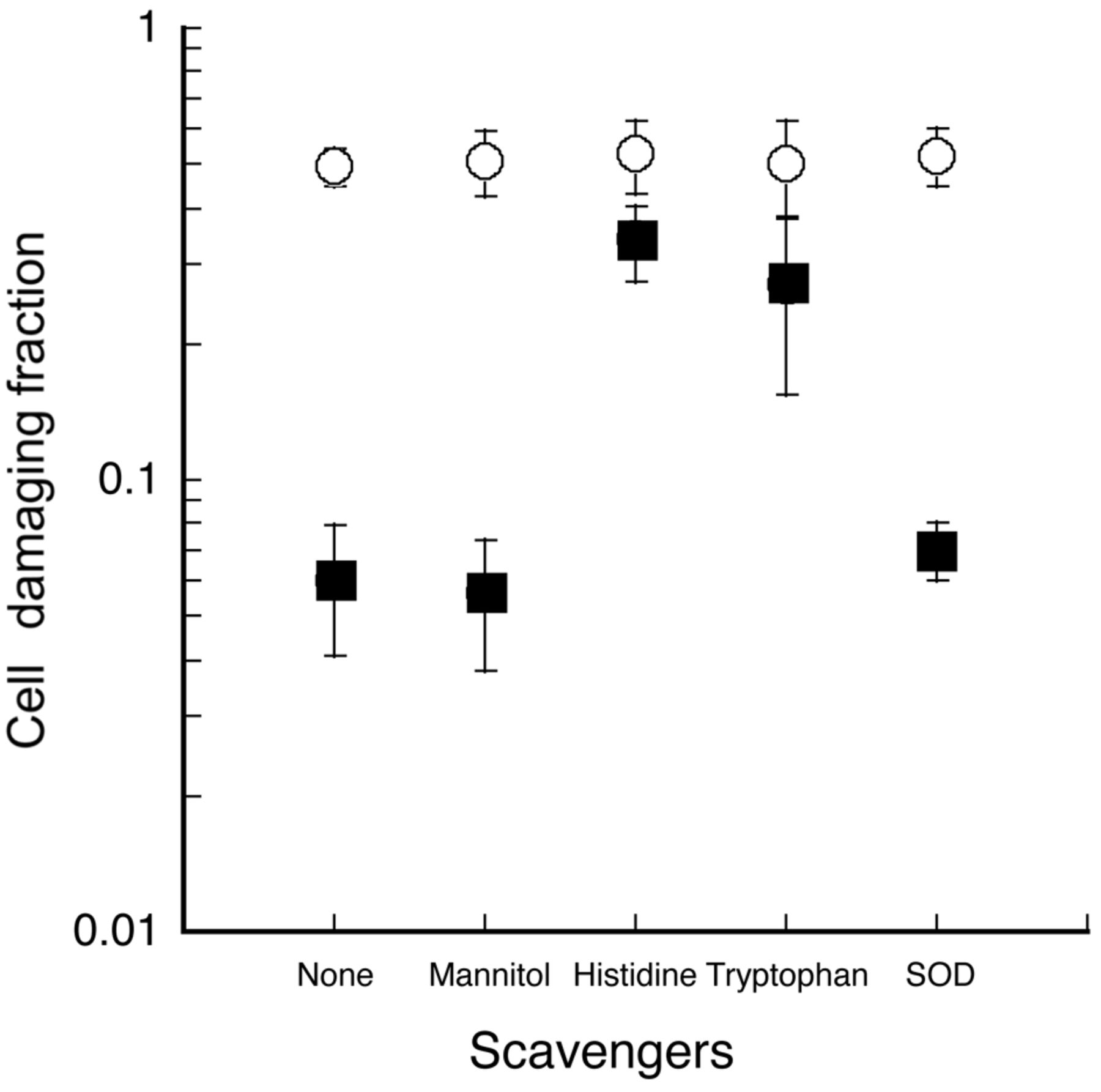
Effect of active oxygen scavengers on ultrasonically-induced cell damage with and without PHF. ●, ultrasound alone; ▪, 80 μM PHF + ultrasound. Degrees of cell damage in the presence and absence of 100 mM mannitol, 10 mM histidine, 10 mM tryptophan and 100 μg/ml SOD are shown.
Remarkable enhancement of ultrasonically-induced in vitro cell damage was demonstrated with PHF. The cell damage rate with 80 μM PHF was more than three-fold higher than ultrasound alone under the same exposure conditionS. This suggests that PHF has potential as a sensitizer for ultrasound treatment. Here, the ratio of unstained cells was fitted using a least squares method to a monoexponential function of ultrasonic exposure time. Many mathematical models have been proposed to further elucidate details of the behavior of cell killing. In the present study, however, fitting to a monoexponential curve was employed in order to make an overall comparison with ultrasonically-induced oxidation without introducing any other parameters.
Distinctive enhancement of ultrasonically-induced lipid peroxidation in the same acoustical arrangement, as in the in vitro cell damage experiment, was also demonstrated with PHF. The peroxidation rate with 80 μM PHF was more than three-fold higher than with ultrasound alone.
In the presence of PHF, the cell damage and peroxidation rates display a strong linear correlation, as shown in Figure 6. The observed correlation between these two parameters is consistent with the hypothesis that membrane lipid peroxidation is the primary mechanism of the ultrasonically-induced cell damage in the presence of PHF. The strong correlation observed in the presence of PHF suggests that membrane lipid peroxidation is also an important mechanism for tumor cell damage induced by ultrasound. Kessel et al., based on their observation that sonodynamically-induced cell damage depends on the presence of porphyrins not in the intracellular but in the extracellular space, suggested the involvement of the cell membrane in sonodynamic cytotoxicity. The affinity of PHF for tumor cells should also be considered with respect to sonodynamically-induced cell damage (29).

Effect of active oxygen scavengers on ultrasonic peroxidation with and without PHF. ○, ultrasound alone; ▪, 80 μM PHF + ultrasound. Degrees of peroxidation generation in the presence and absence of 100 mM mannitol, 10 mM histidine, 10 mM tryptophan and 100 μg/ml SOD are shown.
The inhibitory effect of active oxygen scavengers was also examined. Histidine is known to act as a scavenger of singlet oxygen and possibly hydroxyl radicals, while a mannitol concentration of 100 mM is greater than the concentration reported effective for scavenging both photodynamically- and sonodynamically-induced hydroxyl radicals (30). Thus, the significant reduction of both PHF-enhanced cell damage and peroxidation with histidine under the same acoustic condition coupled with the absence of a significant change with either 100 mM mannitol or 100 μg/ml SOD, suggest that singlet oxygen is playing a more important role than hydroxyl radicals and superoxide as a candidate for the active oxygen species accounting for the PHF-enhanced cell damage. The same mechanism has also been hypothesized for photochemically-enhanced cell damage (25, 31).
The ineffectiveness of mannitol, in contrast to histidine and tryptophan, in inhibiting PHF-enhanced active oxygen generation and cell damage could also be explained, at least partly, by the fact that the hydrophilicity of mannitol is much greater than that of histidine. Several mechanisms involving the generation of ROS have been proposed to explain the sonodynamic action of ultrasound sensitizers (32). These activated high energy metabolites are formed during the intermolecular interaction between the excited sonosensitizing compounds and oxygen molecules in the ground state, thereby resulting in intermolecular energy transfer and the formation of singlet oxygen as shown:
 where Sns is sensitizer.
where Sns is sensitizer.
The sensitization mechanism shown above leading to the generation of singlet oxygen is termed type II reaction (32). In the present study and against PHF-mediated enhancement of lipid peroxidation induced by ultrasound, scavengers of singlet oxygen were shown to afford significant protection, while scavengers of superoxide anion and hydroxyl radical did not. In ESR experiments, Kondo and Riesz observed that the sonolysis yield of nitroxide in the absence of either porphyrins or clorins could not be completely inhibited by a hydrophilic active oxygen scavenger at concentrations where the radiolysis yield was completely inhibited (33). The authors explained that the remaining yield from sonolysis occurred due to nitroxides formed in the interfacial and gaseous reaction regions of the cavitated bubbles, where the hydrophilic scavenger molecules are less concentrated.
Rosenthal and Riesz suggested another possible mechanism of sonosensitizer-derived free radicals as mediators for sonodynamically-induced cell damage (34). This statement could also explain the experimental results observed in this study, assuming that histidine is also effective in removing PHF-derived free radicals. Further studies are needed to determine the active oxygen species involved in the observed sonosensitization of cell damage (35).
Conclusion
In conclusion, the enhancement of the rates of both ultrasonically-induced cell damage and lipid peroxidation was demonstrated in the presence of PHF. Both rates were positively correlated with PHF. The strong correlation observed in the presence of PHF suggests that membrane lipid peroxidation is an important mechanism for tumor cell damage induced by ultrasound. The enhancement of both rates by PHF was suppressed by 10 mM histidine or 10 mM tryptophan. These results provide evidence that singlet oxygen plays an important role in PHF-mediated sonosensitization of membranes and that this moiety may be an important mediator of cell destruction in sonodynamic therapy associated with PHF and ultrasound.
Acknowledgements
This work was supported, in part, by a Grant-in-Aid for Scientific Research (C) from Japan Society for the Promotion of Science to Nagahiko Yumita (No. 25350543), Yumiko Iwase (No. 23500569) and Toshio Fukai (No. 23500570).
Footnotes
-
Conflicts of Interest
The Authors have no conflicts of interest to declare.
- Received July 1, 2014.
- Revision received July 30, 2014.
- Accepted August 4, 2014.
- Copyright© 2014 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}