


Abstract
Aim/Materials and Methods: In order to develop better drugs against non-small cell lung cancer (NSCLC), we screened a variety of compounds and treated the human lung adenocarcinoma cell line A549 with different drug concentrations. We then examined the cell viability using the MTT assay. Results: Data show that a new candidate drug, acriflavine (ACF), suppresses the viability of A549 cells in a concentration- and time-dependent manner. Flow cytometry analysis revealed that ACF significantly caused cell growth arrest in the G2/M phase on A549 cells. Moreover, ACF decreased Bcl-2 expression and increased Bax expression. The content of cleaved poly(ADP-ribose)polymerase-1 (PARP-1) and caspase-3 are significantly increased. These findings suggest that ACF is cytotoxic against A549 cells and suppresses A549 cells growth through the caspase-3 activation pathway. In the in vivo test, nude mice bearing A549 cells xenografts by intravenous injection were randomly assigned into two groups: control and experimental group. Treatment was initiated 10 days after implantation and intraperitoneal injection of 0.9% normal saline or 2 mg/kg of ACF was continued daily for five weeks. ACF treatment significantly decreased tumor size and tumor spots on lung surface of tumor-bearing mice. Conclusion: ACF can inhibit cell growth in A549 cells. Our results may assist on the delineation of the mechanism(s) leading to NSCLC cell growth inhibition and provide a new antitumor strategy against NSCLC.
Lung adenocarcinoma is the major cause of cancer-related mortality worldwide (1, 2). Non-small cell lung cancer (NSCLC) is any type of epithelial lung cancer other than small cell lung cancer (SCLC). NSCLC is the most common form of lung cancer, accounting for approximately 85% of cases. NSCLCs are relatively insensitive to chemotherapy and radiation therapy compared with SCLC (3). Despite clinical success of concurrent therapeutic approaches against lung cancer including surgical resection, chemotherapy and radiotherapy, marked chances of undesirable and adverse side-effects caused by such therapies need to be managed (4). Most often, the standard first-line chemotherapy of NSCLC treatment remains a combination of cisplatin or carboplatin with third-generation agents including vinorelbine, gemcitabine, pemetrexed, paclitaxel or docetaxel (5). Studies have shown that these models of treatment do not have much benefit and are likely to cause more side-effects. Therefore, more effective treatments for NSCLC are required.
Three main strategies are currently used for the development of novel cancer drugs: rational drug design, drug screening and development of analogs. Therefore, this study aims to apply a high-throughput screen of the cytotoxic activity of over 300 molecules from a commercial library on human pulmonary adenocarcinoma cell line, A549. Using this approach, we successfully identified acriflavine (ACF), an antiseptic agent, as a potential drug for NSCLC treatment. ACF is a mixture of 3,6-diaminoacridine (proflavine) and 3, 6-diamino-10-methylacridinium chloride (trypaflavin) (6-9) used for the treatment of microbial infections for humans and fish, particularly as an antiseptic agent possessing a quaternary ammonium structure known to possess antiseptic agent mechanisms of action, such as inhibition of cell-wall synthesis, protein synthesis and DNA synthesis (10, 11). As shown in an earlier report, the 1:1 mixture of ACF and guanosine has been evaluated as a possible antitumor agent in pre-clinical and clinical studies (11). The antitumor effects of ACF have been recently reported in hepatocellular carcinoma (HCC) and colon cancer (12, 13). In our study, we showed that ACF may suppress cellular NSCLC growth more efficiently than its 6 kinds of compounds with similar structures and promote antitumor efficacy in vivo.

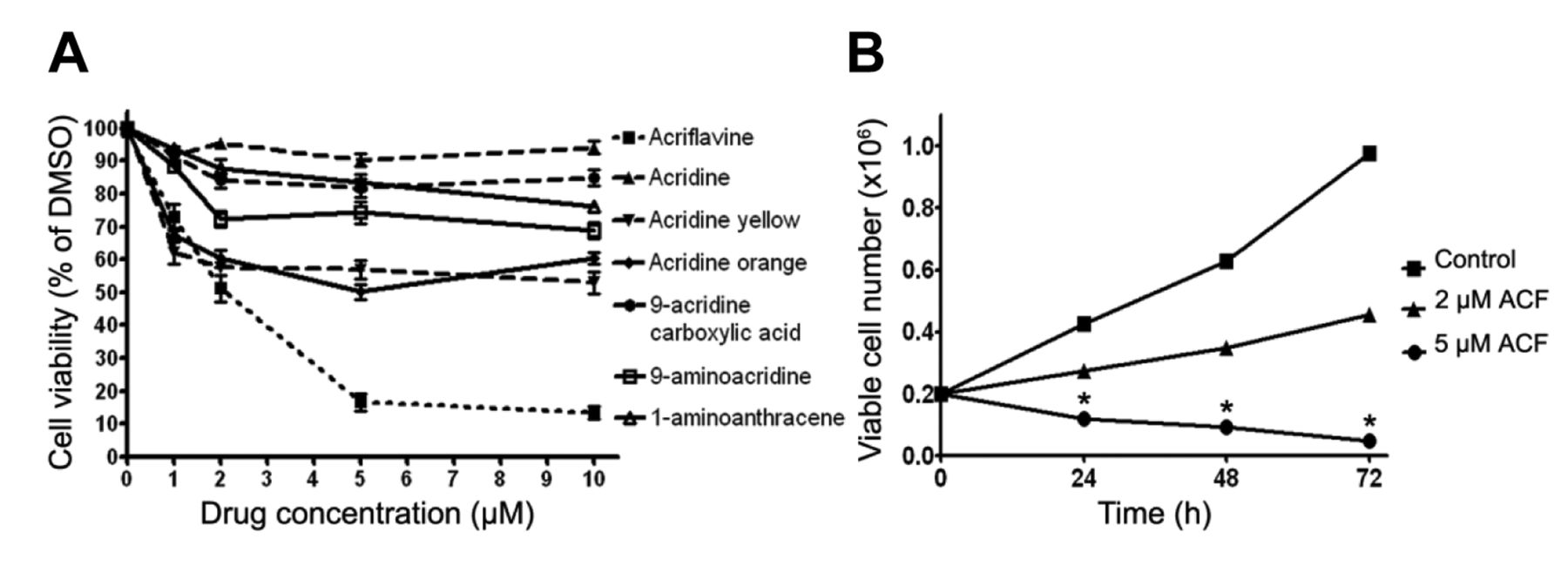
Comparison of the ability of acriflavine (ACF) and its six structurally-similar compounds to arrest cell growth. (A) Screening variety of compounds and treatment of A549 cells with different concentrations of compounds for 48 h. Cell viability was examined using the MTT assay. (B) Effect of ACF on proliferation of A549 cells for 72 h following removal of ACF. *p<0.05.
Materials and Methods
Chemicals, antibodies and cell culture. Compounds were obtained as follows: ACF hydrochloride, acridine, acridine yellow, acridine orange, 9-acridine carboxylic acid, 9-aminoacridine and 1-aminoanthracene were supplied from Sigma-Aldrich (St. Louis, MO, USA). Bcl-2 and poly(ADP-ribose)polymerase-1 (PARP-1) antibodies were purchased from Gene Tex (Irvine, CA, USA). Caspase-3 and Bax antibodies were purchased from Cell Signaling (Danvers, MA, USA). β-actin antibody was purchased from Santa Cruz (Dallas, TX, USA). The lung adenocarcinoma cell line A549 was cultured in Dulbecco's modified Eagle's medium/nutrient mixture F12 (DMEM/F12) medium (Gibco, Carlsbad, CA, USA) supplemented with 10% fetal bovine serum (Thermo, West Palm Beach, FL, USA), 100 U/ml penicillin G, 100 μg/ml streptomycin and 100 μM non-essential amino acids (Gibco) in a humidified atmosphere containing 5% CO2 at 37°C.
Cell viability and proliferation assay. Cell cytotoxicity and viability were determined by measuring the activity of mitochondrial malate dehydrogenase with the tetrazolium dye 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT; Sigma-Aldrich Co.,) assay or Trypan blue exclusion assay (Sigma-Aldrich). A549 cells were seeded at a density of 5×103 with 2×104 cells in each well of 96- and 24-well plates and treated with a range of ACF concentrations (0 μM to 10 μM). After one day of seeding, ACF was added to the culture medium and at 0, 24, 48 and 72 h post-treatment, the MTT reagent was added to each well at a concentration of 1 mg/ml and incubated for 3 h at 37°C. The media were then aspirated and cells were solubilized in 100 and 500 μl dimethyl sulfoxide (DMSO; Sigma-Aldrich Co.,). Cells were incubated for 30 min at room temperature with gentle shaking. Absorbance was measured at 550 nm using a computerized microplate analyzer (BioTek, Winooski, VT, USA) by ELISA. In addition, cells were treated for 72 h with ACF (2 and 5 μM) or DMSO (control) and then incubated in ACF-free medium for the indicated time periods and the number of viable A549 cells was measured using the Trypan blue exclusion assay.
Flow cytometric analysis. A549 cells were treated with 0, 1 and 2 μM of ACF for 48 h. After collection, the cell were washed twice with phosphate-buffered saline (PBS; Gibco), and then fixed in 70% ethanol overnight at -20°C. Fixed cells were stained with a solution containing 10 μg/ml propidium iodide (PI, Sigma), 100 μg/ml RNase A (Sigma) and 0.1% Triton X-100 (Sigma) at 37°C in the dark. After 30 min, cell cycle distribution was performed using FACScan (BD Biosciences, San Jose, CA, USA) flow cytometry.
Western blot analysis. Cells were washed with PBS and lysed with cold radioimmunoprecipitation assay buffer (RIPA buffer; Thermo) supplemented with protease and phosphatase inhibitors. The protein concentration was determined using the Bradford assay. Proteins were separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE). After electrophoresis, proteins were transferred to polyvinylidene difluoride (PVDF) membranes by electroblotting and then blocked in TBST (50 mM Tris-HCl, pH 7.4, 150 mM NaCl, and 0.2% Tween-20, Sigma) containing 5% nonfat milk. Membranes were incubated with the primary antibody (1:1000) at 4°C overnight and then washed with TBST thrice. The blots were then incubated with an appropriate horseradish peroxidase (HRP)-conjugated secondary antibody at room temperature for 1 h. After washing, membranes were incubated with ECL Western blotting detection reagent and exposed to X-ray films.
Animal model. The effects of ACF on nude mice were evaluated using a tumor xenograft model. Four-week-old male nude mice (BALB/cAnN.Cg-Foxnlnu/CrlNarl) were obtained from the National Laboratory Animal Center (Taipei, Taiwan). Nude mice were intravenously injected through tail veins with A549 cells (3×106) and treatment was performed 10 days post-injection. Tumor growth was captured using the IVIS Lumina image system (Xenogen Biosciences, Cranbury, NJ, USA). Once the tumors were established, the mice were treated with ACF daily through intraperitoneal injection for six consecutive weeks with 0.9% normal saline as control group (n=4) or 2 mg/kg ACF (60 μl ACF) for the experimental group (n=4). Four animals are considered the minimum number of animals required provided that the harvested data is statistically significant (14). Experiments were performed in duplicate. Tumor size was measured once a week with a caliper using the following formula: tumor volume (mm3)=0.5236 length ×width2. Control and treated mice were sacrificed after five weeks and the removed tumors were fixed in formalin. Tumor tissues isolated from each group were examined by hetatoxylin and eosin staining (Sigma-Aldrich).
Statistical analysis. Values are reported as the mean±standard deviation (SD) and data were analyzed using one way analysis of variance (ANOVA) followed by post hoc tests for multiple comparisons. The Student's t-test was used in two-group comparisons. Values of p<0.05 were considered to be statistically significant.
Results
ACF suppresses the growth of A549 cells in vitro. We performed random screening of hundreds of compounds from a commercial library and found ACF with a novel antitumor activity in the A549 cell line. We subsequently compared for the cellular viability of A549 after treatment with ACF and its 6 compounds (acridine, acridine yellow, acridine orange, 9-acridine carboxylic acid, 9-aminoacridine, 1-aminoanthracene) whose structure are somewhat similar to ACF. A549 cells were treated with different concentrations of the compounds at a concentration range from 0 μM to 10 μM for 48 h and ACF relative to other analogues, which at low concentrations (1, 2, 5 and 10 μM) exhibited an apparent inhibition effect on cell growth in a significant dose-dependent manner. The data are shown in Figure 1A. ACF-induced reduction of cell growth rate is evident at a starting dose of 5 μM in A549 cells at the 48-hour time-point.
Recovery of proliferation assay following removal of ACF. To examine whether ACF is cytotoxic and not cytostatic in A549 cells, we evaluated the ability of ACF-treated cells to recover their viability and proliferation activity. For this reason, A549 cells were incubated with 0, 2 and 5 μM ACF for 3 days and then cultured in ACF-free medium for different periods of time and cell viability was measured using the Trypan blue exclusion assay (Figure 1B). These data indicate that ACF is cytotoxic at the dose of 5 μM since cells were unable to resume proliferation after ACF removal.

Effect of ACF on the cell cycle distribution of A549 cells by flow cytometry. A549 cells were incubated without or with 1, 2 and 5 μM of ACF for 48 h. (A) control, (B) 1 μM ACF exposure, (C) 2 μM ACF exposure and (D) 5 μM ACF exposure for 48 h. (E) A549 cells were predominantly accumulated in the G2/M-phase. *p<0.05 and ** p<0.01 the level are increased/decreased as compared to the same fraction in the dose 0 control group.

Total A549 cell lysates analyzed by western blot with antibodies against PARP-1, Bcl-2, Bax and caspase-3. A549 cells were incubated without or with 1, 2, 5, and 10 μM of ACF for 48 h. Effects of ACF on protein expression in (A) dose-dependent and (B) time-dependent manners.
Alteration of cell cycle by ACF in A549 cells. To evaluate whether ACF-induced cell growth retardation is mediated by alterations in cell-cycle progression, we performed flow cytometry analysis to determine whether the change in the cell cycle profile was induced by ACF. A549 cells were treated with different concentrations of ACF for 48 h and then collected and stained with PI through FACS analysis. Cell-cycle analysis showed that ACF caused a significant G2/M arrest at the expense of G1 phase cell population accumulated at 1 μM of treatment and this effect was sustained with increased treatment concentration (2 μM) on the A549 cell line (Figure 2). We found that ACF treatment of A549 cells could effectively induce cell-cycle arrest in G2/M phase.
ACF induces apoptosis in A549 cells. Bcl-2 family and caspase-3 activation is considered to be a key hallmark of apoptosis (15). We investigated the effects of ACF on the expression of the Bcl-2 family and activation of caspase-3 in A549 cells that were exposed to increasing concentrations of ACF for 48 h. We measured the protein levels of apoptosis, including the pro-apoptotic protein Bax and anti-apopotic protein Bcl-2. In our study, ACF increased the expression of the cleaved-caspase-3, cleaved PARP-1 and Bax, but decreased the expression of Bcl-2 in A549 cells. As shown in Figure 3, ACF treatment activated caspase-3, followed by subsequent PARP-1 cleavage, decreased levels of Bcl-2 and increased levels of Bax. These findings suggest that ACF could induce apoptosis through the mitochondria-mediated apoptotic pathway, as well as to promote A549 cells apoptosis.
ACF inhibits tumor growth. To evaluate the therapeutic effects of ACF, nude mice bearing A549 cells xenografts were intraperitoneally injected with 2 mg/kg ACF daily for six weeks. As shown in Figure 4A, compared with the tumor size in the normal saline group, ACF-treated mice had smaller tumor sizes and fewer tumor spots on the lung surface after treatment (Figure 4C). ACF significantly suppressed tumor growth at the dose of 2 mg/kg for a 6-week period compared to the control group. At this very same time point of 42 days, the mice were sacrificed and tumor size was evaluated. The mean size of tumors (week 6) of the normal saline group was 1,230±120 mm3 and 796.3±21 mm3 of the ACF group. This significant decrease in tumor growth in mice treated with ACF compared to control is shown in Figure 4B. Likewise, we also found the fibrosis levels to be remarkably induced in lung slices (data not shown). Tumor invasion in lung lobes was significantly lower in the ACF-treatment group than those in the control group (Figure 4C). These findings demonstrate that tumor size is reduced by using the ACF treatment.
Discussion
In the present study, we selected an analogue of acridine and evaluated its effect on cell viability in a human tumor cell line. ACF was a more apoptotic inducer and more cytotoxic than its structural analogues in A549 cells.
ACF-induced cell apoptosis via the caspase signaling pathway. Apoptosis, a form of programmed cell death, is tightly regulated by the pro-apoptotic and anti-apoptotic members of the Bcl-2 family in the intrinsic apoptotic pathway (15). Therefore, the Bcl-2 family serves an important function in the intrinsic apoptotic pathway. Pro-apoptotic Bax is known to promote intrinsic apoptosis by forming oligomers in the mitochondrial outer membrane, whereas anti-apoptotic Bcl-2 blocks mitochondrial apoptosis by hindering the release and oligomerization of Bax (16, 17). In the present study, we found that ACF decreased the expression of Bcl-2 in A549 cells (Figure 3). Thus, we suggest that ACF induces apoptotic cell death through the caspase signaling pathway in NSCLC. The activity pattern observed for ACF is promising from a clinical perspective; however, further understanding the mechanisms of ACF-induced cell apoptosis in NSCLC is beneficial for optimal treatment.

ACF results in significant inhibition of tumor growth in A549 tumor xenografts established in nude mice. (A) Tumor size reduction in ACF-treated mice. (B) Quantification of tumor size. Data represent the mean±SD of experiments performed in triplicate. *p<0.05 compared to control tumors. (C) Representative low-magnification images of hematoxylin and eosin staining of representative lung lobes after treatment.
Molecular mechanism of the effect of ACF in A549 cells. A key point is what makes A549 cells extraordinarily sensitive to ACF. A common property of acridine derivatives, such as ACF, is their capability to intercalate nucleic acids and inhibit topoisomerase II (18). Furthermore, ACF binds to and disrupts the cell surface membrane, thus resulting in inhibition of protein kinase C, which is considered to account for some anti-tumor effects (19). ACF reportedly acts against colorectal cancer (20). ACF was recently found to inhibit the dimerization of hypoxia-inducible factor-1α (HIF-1α) and HIF-1β, which suppress vascular endothelial growth factor (VEGF) and glucose transporter 1 (GLUT 1) gene expressions and are associated with anti-angiogenesis effects and tumor growth inhibition (9). In our study, ACF also decreased the expression of GLUT 1 mRNA in A549 cells (data not shown). Further studies are required to understand the role of ACF in the angiogenesis of NSCLC cells, as well as the mechanism of its renewal.
Inhibition of tumorigenesis by ACF. Numerous stimulatory factors contribute to tumor angiogenesis, such as HIF-1α and VEGF which hold key role in tumors. In pre-clinical studies, the inhibition of HIF-1 activity has marked effects on tumor growth (21). In a previous study, guanosine was found to enhance the anti-tumor effects of ACF against a variety of cancer cells without serious adverse effects (7, 20). In our animal model experiments, we observed the suppressive role of ACF in the progression of tumor growth; ACF treatment was found to reduce tumor sizes in nude mice (Figure 4). The results provide evidence that ACF treatment significantly alters viability of lung adenocarcinoma A549 cells through induction of apoptotic signaling and may shed light on the anticancer effects of ACF, thus providing a pre-clinical basis for potential anticancer application.
Acknowledgements
This work was supported by grants from the Ministry of Science and Technology, Republic of China (NSC 99-2632-B-039-001-MY3, NSC 102-2320-B-039-049, NSC101-2911-I-002-303 and NSC102-2911-I-002-303), Taiwan, Republic of China.
- Received July 2, 2014.
- Revision received August 5, 2014.
- Accepted August 7, 2014.
- Copyright© 2014 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved





{kind=link}
{kind=link}
{kind=link}
{kind=link}