


Abstract
Background: SET domain containing protein 2 (SETD2) is a histone methyltransferase that is involved in transcriptional elongation. We previously demonstrated SETD2 to be a potential tumour suppressor gene in breast cancer. The aim of this study was to compare SETD2 expression in breast cancer with that in adjacent non-cancerous breast tissue (ANCT) in paired samples. A hypothesis is proposed that explains the mode of action of SETD2 as a tumour suppressor gene. Materials and Methods: Paired samples of tumour and adjacent non-cancerous tissue (ANCT) from 25 patients were analysed. The levels of transcription of SETD2 were determined using quantitative polymerase chain reaction and normalized against cytokeratin 19. Immunohistochemical staining with appropriate antibodies against SETD2 protein was also performed in selected samples. Results: Levels of SETD2 mRNA were significantly higher in ANCT when compared to those in tumour samples (p=0.01). Immunohistochemistry also demonstrated a higher protein expression in ANCT. Conclusion: This study offers further evidence that SETD2 behaves like a tumour suppressor gene. Our hypothesis links SETD2 mode of action with telomerase regulation through human telomerase reverse transcriptase (hTERT). Several studies have emphasised the importance of histone methylation of hTERT promotor in telomerase regulation. SETD2 function of histone methylation could be the missing link in this chain which could explain the potential tumour suppressor function of SETD2.
Histone-lysine N-methyltransferase (SETD2) is a protein that methylates Lys-36 of histone H3. H3 Lys-36 methylation represents a specific tag for epigenetic transcriptional activation (1).
The SETD2 gene is located on the 3p21.31 region of chromosome 3 which has been shown to be associated with telomerase suppression and permanent growth arrest of breast cancer cells (2, 3). SETD2 plays a role in chromatin structure modulation during transcriptional elongation via its interaction with hyperphosphorylated (POLR2A) polymerase (RNA) II (DNA-directed) polypeptide A. It also may act as a transcription activator that binds to promoters (4). Previous studies showed that histone modifications, which include acetylation, phosphorylation, methylation and ubiquitination, play key roles in gene regulation (5). These modifications create both agonistic and antagonistic signals that correlate with the transcriptional activity of a gene, through engaging protein complexes or through changing the structure of chromatin to allow access for RNA polymerase to initiate transcription (1). We have recently shown that mRNA expression of SETD2 in breast cancer correlates with the progression of the disease and the gene behaves like a tumour suppressor in human breast cancer (3). SETD2 may exert its function through interaction with p53 and by selectively regulating the transcription factor activity of p53 by enhancing the protein stability of p53 (6). The aim of this study was to compare SETD2 expression in breast cancer with that in adjacent non-cancerous breast tissue (ANCT) in paired samples.
Materials and Methods
Samples. Institutional guidelines, including ethical approval and informed consent, were followed. Paired samples of tumour and adjacent non-cancerous tissue (ANCT) from 25 patients were analysed. A consultant pathologist examined haematoxylin and eosin stained frozen sections to verify the presence of tumour cells in the collected samples. All tissues were randomly numbered and the details were only made known after all analyses were completed.
Materials. RNA extraction kits and reverse transcription kits were obtained from Sigma-Aldrich Ltd (Poole, Dorset, England, UK). PCR primers were designed using Beacon Designer (Palo Alto, CA, USA) and synthesized by Sigma-Aldrich. Custom made hot-start Master-mix for quantitative PCR was obtained from Abgene (Surrey, England, UK) (7, 8).
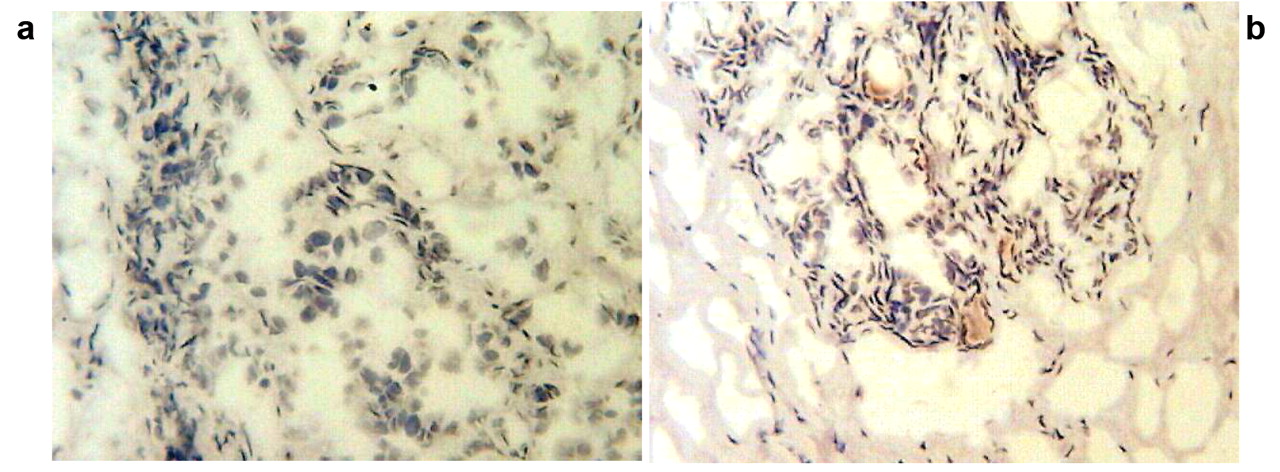
Immunohistochemistry on tumour and normal (ANCT) tissue samples. a: Tumour sample stain-free; b. ANCT sample showing staining.
Tissue processing, RNA extraction and cDNA synthesis. Frozen sections of tissue were cut at a thickness of 5-10 mm and kept for routine histological analysis. An additional 15-20 sections were mixed and homogenized in ice-cold RNA extraction solution using a hand-held homogenizer. The concentration of RNA was determined using UV spectrophotometry. Reverse transcription was carried out using a reverse transcription kit with an anchored oligo (dT) primer supplied by Abgene, using 1 mg of total RNA in a 96-well plate. The quality of cDNA was verified using B-actin primers.
Quantitative analysis. The level of SETD2 transcripts from the above prepared DNA was determined using real-time quantitative PCR based on Amplifluor technology, modified from a method reported previously (9). PCR primers were designed using Beacon Designer software, but to the reverse primer an additional sequence, known as the Z sequence (5′-ACTGAACCTGACCGTACA-3′) which is complementary to the universal Z probe (Intergen, Inc., Oxford, UK) was added. The product expands one intron. The reaction was carried out using the following: Hotstart Q-master mix (Abgene), 10 pmol of specific forward primer, 1 pmol reverse primer with the Z sequence, 10 pmol of FAM (fluorogenic reporter dye, carboxyfluorescein)-tagged probe (Intergen, Inc.), and cDNA from 50 ng of RNA. The reaction was carried out using the IcyclerIQ (Bio-Rad Ltd, Hemel Heamstead, England, UK), which is equipped with an optical unit that allows real-time detection of 96 reactions, under the following conditions: 94°C for 12 min and 50 cycles of 94°C for 15, 55°C for 40 s, and 72°C for 20. The levels of the transcript were generated from a standard that was simultaneously amplified with the samples. Levels of mRNA expression were then normalized against expression of cytokeratine fragment 19 (CK19), already measured in these specimens, to correct for varying amounts of epithelial tissue between samples. CK19 transcripts were quantified as previously reported (9). With every PCR run, a negative control without a template and a known cDNA reference sample as a positive control were included.
Immunohistochemistry. The Vector Laboratories Universal IHC kit was used to carry out immunohistochemistry using a rabbit antibody supplied by Abnorva (catalogue number: pab2373). The staining dilution was 1:100 (primary) and 1:50 (secondary).
Statistical analysis. The Mann–Whitney U-test was used for statistical analysis. In each case, the true copy number was used for statistical analysis and hence samples were not classified as positive or negative. Statistical analysis was carried out using Minitab (14.1) using a custom written macro (Stat 2005.mtw).
Results
The levels of SETD2 mRNA transcription in ANCT were significantly higher (more than 100-fold) than in tumour samples (median: 1772 vs. 16, p=0.01).
Immunohistochemistry was used to verify the protein content of randomly selected samples and this showed higher protein expression in ANCT compared with tumours. Figure 1 highlights the difference between normal tissue containing some staining and the malignant samples, which are virtually stain-free.
Discussion
We have recently published the first study in the literature to examine the direct relationship between SETD2 mRNA expression and breast cancer (3). We demonstrated that this gene potentially had a tumour suppressor function and its mRNA expression may play a role as a prognostic indicator in human breast cancer. The SETD2 gene is located within the 3p21.31 chromosomal region which was previously shown to repress telomerase using microcellular transfer techniques (2).
This study sheds further light on the expression of SETD2 and shows that levels are significantly lower in tumour cells, even compared to normal breast tissue from the same individual patient.
It has been established that telomerase activation is a critical step for human carcinogenesis through the maintenance of telomeres, but the activation mechanism during carcinogenesis remains unclear. Transcriptional regulation of the human telomerase reverse transcriptase (hTERT) gene is the major mechanism for cancer-specific activation of telomerase, and a number of factors have been identified to directly or indirectly regulate the hTERT promoter (10). Certain transcription factors such as tumour suppressor gene products (P53) are able to control hTERT transcription when overexpressed. Recent studies have shown that an endogenous mechanism restored by a telomerase repressor gene on chromosome 3 acts on a specific DNA element within the hTERT gene promoter to repress the hTERT transcription (11). We recently examined 9 genes on the short arm of chromosome 3 (3p21.31 region) and found SETD2 to be the only one with tumour suppressor features (3). It remains unclear how SETD2 exerts this effect.
Recent studies have shown that suppression of hTERT gene promoter is mediated by histone modification (7, 11, 12).
Alteration of the chromatin structure via DNA methylation or modulation of nucleosome histones has been suggested to be important for regulation of the hTERT promoter. DNA demethylation or histone methylation around the transcription start site of the hTERT promoter triggers the recruitment of histone acetyltransferase activity, allowing hTERT transcription (10).
Finally, our previous research demonstrating that the chromosome 3 repressor directly alters the chromatin conformation of hTERT in breast cancer tumour cells to a more open state, permissive for transcription, lends further support to our hypothesis (13).
We hypothesize that SETD2 may interact with hTERT through histone methylation of the promoter region to regulate telomerase activity. In non-cancerous cells, telomerase is repressed by normal histone methylation mediated by normal SETD2 expression. Deletion or mutation of SETD2 leads to reduced histone methylation of hTERT promoter thus leading to increased hTERT transcription and telomerase up-regulation. Further research is required to investigate this hypothesis.
- Received May 30, 2010.
- Revision received June 23, 2010.
- Accepted July 1, 2010.
- Copyright© 2010 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
Jump to section
Related Articles
Cited By...
- Methylation of lysine 36 on histone H3 is required to control transposon activities in somatic cells
- Therapeutic targeting of SETD2-deficient cancer cells with the small-molecule compound RITA
- Intact nucleosomal context enables chromodomain reader MRG15 to distinguish H3K36me3 from -me2
- Utilizing a resource of enrichment profiles in plasma for the systematic assessment of antibody selectivity
- SETD2 Is Recurrently Mutated in Whole-Exome Sequenced Canine Osteosarcoma
- SETting the Stage for Cancer Development: SETD2 and the Consequences of Lost Methylation
- A Systematic Approach to Defining the microRNA Landscape in Metastasis
- Mutations in SETD2 cause a novel overgrowth condition
- Trimethylation of Histone H3 Lysine 36 by Human Methyltransferase PRDM9 Protein