


Abstract
Background: Occludin is an integral membrane protein localised at tight junctions (TJ). There is no consensus regarding its paramount role in TJ. In previous work we showed that occludin is aberrantly expressed in both human breast tissues and cancer cell lines. This study demonstrates a link to bone metastasis in human cancer. Materials and Methods: Primary breast tumours (n=124) and matched normal tissues (n=30) were processed for quantitative polymerase chain reaction (QPCR) analysis. A hammerhead ribozyme was constructed to create occludin knockdown cell lines, MDA-MB-231ΔOcc and PC-3ΔOcc. The effect of human bone matrix extract (BME) was investigated using cell growth and electric cell impedance sensing (ECIS) technology to measure changes in attachment/migration. Trans-epithelial resistance (TER) was measured for assessing changes in TJ function. Cells used were MDA-MB-231, PC-3, CORL-23, SKMES-1 and A-549 human cancer cell lines. Results: Tumours from patients with bone metastasis had significantly lower occludin expression compared to those remaining alive/well (60.7±21 vs. 331±98, respectively, p=0.008). This was striking in ductal carcinomas, where patients alive/well had significantly higher occludin expression compared to those with bone metastasis (391±12.5 vs. 67.9±28, respectively, p=0.0014). ECIS demonstrated that MDA-MB-231ΔOcc showed reduced attachment to 5% BME compared to controls (84% vs. 100%) that prevented closure of wounded cell layers. Moreover, these cells had reduced growth on BME. In addition, BME changed the TER of a number of human cell lines and was able to effect changes in the growth of MDA-MB-241 and PC-3 cells, with greater effect on knockdown cells. Conclusion: This is the first study to demonstrate that occludin expression has a clear relationship with bone metastasis in human cancer. The discrepancy between this and the in vitro data indicating a reduction in migration/growth rate of occludin knockdown indicates that loss of occludin leads to complex changes in human cancer cell phenotype.
Occludin was one of the first identified transmembrane tight junction (TJ) proteins (1, 2). It has four transmembrane domains in its N-terminal half, with both the N- and C-termini located in the cytoplasm, and the C-terminal (~150 amino acids) binding to Zonula occludens (ZO-1) (3, 4). The cytoplasmic domain (domain E) also interacts with both ZO-1 and -2. This conformation gives rise to two transmembrane loops projecting into the paracellular space that interact with occludin loops originating from a neighbouring cell, therefore promoting the interaction and sealing of the paracellular space (5). The C-terminal of occludin is sufficient to mediate endocytosis, as it governs the intracellular transport of occludin (6). Occludin is a functional component of the TJ and widely expressed in cells and tissues that have TJs (6).
It has become apparent over the past decade that the TJ is a key component in the process of cancer metastasis. For cancer cells to successfully metastasize, they must pass through a series of barriers in order to proceed to a secondary foci. TJs are a defence barrier against endothelial invasion (both in intravasation and extravasation), as they are known to work as a barrier to the paracellular passage of cells and substances between the epithelial or endothelial cells (7). Preferential organs for distant metastasis of tumours are the lung, liver, brain and bone and prostate, lung, breast, kidney and thyroid cancer account for 80% of skeletal metastases (8).
Although numerous events are believed to contribute to the process of metastasis, it is widely accepted that the loss of cell–cell adhesion in the neoplastic epithelium is necessary for the invasion of the surrounding stromal elements and subsequent metastatic events (9). Our previous work showed that occludin is expressed in variant forms in human breast cancer cell lines and in human breast cancer, and the loss of occludin expression after knockdown, results in breast cancer cell lines with a significantly altered phenotype and reduced TJ functionality. We have also demonstrated that occludin expression is lost in human breast cancer tissues and is associated with poorer patient prognosis (10). Our aim in the present study was to determine if loss of occludin is linked to metastatic disease to bone, and if so, how the bone environment is able to affect the function of occludin knockdown cells.
Materials and Methods
Reagents and antibodies. Carbonate filter inserts with pore size of 0.4 μm (for 24 well plates) were from Greiner Bio-One Ltd (Stonehouse, Gloucestershire, UK). Gold electrode arrays (8W10E) were purchased from Applied Biophysics Inc (Troy, New Y NY, USA).
Cell lines. The human cancer cell lines MDA-MB-231 (breast), PC-3 (prostate), A-549 (lung), SKMES-1 (lung) and CORL-23 (lung) were from the American Type Cell Culture (LGC, Teddington, Middlesex, UK). The cells were routinely maintained in Dulbecco's modified Eagle's medium (DMEM; Sigma-Aldrich Ltd, Poole, Dorset, UK) supplemented with 10% foetal calf serum (FCS), penicillin and streptomycin (Sigma-Aldrich Ltd).
Construction of the occludin ribozyme for knockdown experiments. Ribozyme transgene to occludin was constructed as previously reported (10). The following DNA oligonucleotides were used to generate ribozymes: OCCrib1F CTGCAGGCTGGCTGAGAGAG CATTGGTCCTGATGAGTCCGTGAGGA and OCCrib1RACTA GTGGAGAGATGCATGTTTTCGTCCTCACGGACT. Ribozyme gene cassettes were cloned into pEF/V5-His TOPO® TA parent vector (Invitrogen Ltd., Paisley, UK). Vectors were transfected into chemically competent Escherichia coli, and bacterial colonies were grown on agar plates. Colonies were examined using polymerase chain reaction (PCR) technique to confirm absence of occludin expression. Plasmid purification and extraction were performed using a plasmid extraction kit (Qiafilter; Qiagen, Crawley, Surrey, UK). The concentration of each plasmid was determined using a spectrophotometer (WPA UV 1101; Biotech Photometer, Cambridge, UK). MB-MDA MB-231 human breast cancer cells or PC-3 cells (105 cells in 100μl) were mixed with 10 μl of occludin or control plasmid for 2 min and then electroporated at 170 V with an electroporator (Easyjet; Flowgen, SLS, Hessel, Yorkshire, UK). Cells were immediately transferred to complete medium pre-warmed to 37°C and plated into 6-well plates. After 24 h, cells were transferred to 25 ml cell culture flasks and selected using blasticidin at 5 μg/ml (Melford Ipswich, Suffolk, UK). The medium was changed every 3-4 days. After 4 weeks, cells were changed to maintenance medium (with 1 μg/ml blasticidin). Occludin-knockdown cells were designated MDA MB-231ΔOcc and PC-3ΔOcc, and control plasmid cells MDA MB-231Plas and PC-3Plas. Stably transfected cells were then used for subsequent assays (after 6 weeks of culture and confirmation of expression). In vitro biological assays analysing occludin knockdown cells were used to determine what, if any, the effect of occludin knockdown had on the human cancer cell lines MDA-MB-231 and PC-3.
Trans-epithelial resistance (TER). TER was measured with an EVOM voltohmmeter (World Precision Instruments, Aston, Herts, UK), equipped with a pair of STX-2 chopstick electrodes (WPI, Sarasota, FL, USA), as we previously reported (11, 12). Briefly, cells were seeded into the 0.4 μm pore size insert (Greiner Bio-One Ltd) and allowed to reach full confluence, after which fresh medium was replaced for further experiments. Inserts without cells, inserts with cells in medium, and inserts with cells with BME were tested for a period of 2 h. Electrodes were placed at the upper and lower chambers and resistance measured with the voltohmmeter.
Cell-matrix adhesion assay. The cell-matrix attachment assay was carried out as previously reported (11). Briefly, Matrigel (10 μg/well) was added to a 96-well plates, which were incubated for 24 h to allow binding of matrix protein to the surface of the well. The plates were then washed and blocked with 5% bovine serum albumin, (Sigma-Aldrich Ltd). Cells were added at 104/well for 30 min, followed by aspiration and washing. The number of attached cells was determined by direct counting under microscope (seven counts per experimental setting).
Growth assay. Standard growth assays using crystal violet were carried out over 5 days to determine whether the knockdown of occludin changed the growth rate of MDA-MB-231 and PC-3 cancer cells.
Tissue collection and preparation. Breast tissue samples (124 tumour and 33 matched normal tissues) (11) were collected and immediately frozen in liquid nitrogen before processing; a portion of each sample was used for quantitative-PCR (Q-PCR) analysis, a portion for immunohistochemical analysis, and a portion for routine histological examination. RNA was isolated from tissue samples using standard RNA-zol procedures. For reverse transcriptase polymerase chain reaction (RT-PCR), cDNA was synthesized in a 20 μl reaction mixture using 1 μg RNA, as described in the protocol (AB Gene Reverse Transcription System, Epsom, Surrey, UK).
The anonymised breast tissue samples were obtained within the guidelines of the appropriate Ethics Committee (Bfo Taf Health Authority 01/4303 and 01/4046). Informed patient consent was not applicable in this instance (as stated in the Human Tissue Act 2004, UK).
Quantitative-PCR. Specific primer pairs for occludin were designed by the Authors using a Beacon Designer software (Biosoft International, Palo Alto, CA, USA) and manufactured by Invitrogen (Invitrogen Life Technologies, Paisley, Scotland, UK), each amplifying a region spanning at least one intron, generating an approximately 100 base pair product from both the control plasmid and cDNA. PCR primers were: OCZF GAATTCAAACCGAAT CATTG and OCZR ACTGAACCTGACCGTACATGAAGAATTTC ATCTTCTGG. The Q-PCR system used the Amplofluor™ Uniprimer™ system (Intergen Company, Oxford, UK) and Thermo-Start® (ABgene, Epsom, Surrey, UK). Using the Icycler IQ system (Bio-Rad, Hemel Hempstead, Hertfordshire, UK), which incorporates a gradient thermocycler and a 96-channel optical unit, the plasmid standards and breast cancer cDNA were simultaneously assayed in duplicate reactions using a standard hot-start Q-PCR master mix. Quantitative-PCR conditions were as follows: Enzyme activation 95°C for 12 min, one cycle, followed by 60 cycles of denaturing: 95°C for 15 s; annealing: 55°C for 40 s; Extension: 72°C for 25 s. Using purified plasmids as internal standards, the level of cDNA (no. of copies/50 ng RNA) in the breast cancer samples were calculated. Quantitative-PCR for β-actin was also performed on the same samples in order to correct for any residual differences in the initial level of RNA in the specimens (in addition to spectrophotometry). Results were then normalized using cytokeratin-19 levels in the same tissues. The products of Q-PCR were verified on agarose gels (not shown).
Bone matrix extract (BME) preparation. Fresh bone tissues were collected immediately after surgery, under a local ethical approval, and immediately stored at −80°C until use. The tissues were first processed using a surgical macerator to obtain a fine bone mixture under sterile conditions. The mixture was resuspended in a sterile non-reducing buffered saline solution (BSS) buffer and placed in a Bioruptor™ unit (Thermo Fisher Scientific, Inchinnan, Paisley, UK). This was subject to a 5-min interrupted pulse for 20 s with 20-s pause. The soluble proteins were recovered after removal of debris at 3000 × g in a refrigerated centrifuge. The concentration of protein was quantified using a Bio-Rad protein quantitation kit and reconstituted to give a preparation with 2 mg/ml protein. The proteins were stored at −80°C until use. The nature of the protein was verified by using western blotting by probing with antibodies recognising small and large proteins, namely anti-actin and anti-cadherin and anti-cMET, respectively. The effects of the BME on the growth of cells were tested over a concentration range of 20-1000 μg/ml.
ECIS measurements for attachment and migration. The ECIS 1600R instrument, (Applied Biophysics) system was used to measure transepithelial impedance in the transfected and control cells to ascertain differences in cell attachment and migration with and without BME. For ECIS measurements, 105 cells were seeded into each well of an electrode array (8W10E) with 0.4 ml of medium per well. For ECIS-based adhesion experiments, the gold ECIS electrodes were coated with/without BME (5%) for 1 h at 37°C. Cells were seeded and ECIS was subsequently monitored for up to 3 days. Migration/wound-healing or barrier function assays were carried out with ECIS with confluent cell layers cultured in ECIS electrode arrays and submitted to an elevated voltage pulse of 40-kHz frequency, 4.5-V amplitude for two 30 s bursts followed by continuous resistance measurements for 4 h.
Statistical analysis. Statistical analysis was performed by MINITAB version 13.32 (Minitab Inc. State College, PA, USA) using a two-sample Student t-test and the non-parametric Mann–Whitney or Kruskal-Wallis test, where appropriate. In addition, Microsoft Excel Professional (Microsoft, Reading, Berkshire, UK) was used to calculate correlation coefficients and SPSS (12.01) (IBM, Portsmouth, Hampshire, UK) for survival curves.
Results
Expression of occludin in human metastatic breast cancer. Continning our previous work examining the role of occludin in breast cancer (10), we herein further studied the expression of occludin and how it could be related to metastatic disease. Quantitative-PCR analysis revealed that occludin was decreased in patients with bone metastasis from breast cancer (p=0.08) and indeed, was significantly decreased in patients with all types of metastatic disease (p=0.024) (Figure 1A). In addition to this, when looking more closely at the type of breast cancer and occludin expression, we found that loss of occludin was significantly related to metastatic disease in patients with ductal tumours (p=0.02) and also to bone metastatic disease (p=0.0014) (Figure 1B). Survival analysis indicated that the loss of occludin was associated with poor survival in patients with bone metastasis (Figure 1C), although this did not reach significance.
Effect of BME and occludin knockdown in human breast cancer cell migration and attachment. Continuing on from the Q-PCR data linking the loss of occludin and metastasis to bone in human breast cancer, we investigated the effect of BME on the behaviour of a metastatic breast cancer cell line, MDA-MB-231. We also used occludin knockdown cells to assess any effect this might have on BME treatment [as described in Martin et al. (10)]. Knockdown of occludin caused a reduction in the migration of MDA-MB-231 cells (p<0.05). After addition of a low concentration of BME (5%), it was seen that MDA-MB-231 control cells showed an increased migratory rate, but this did not reach significance. The addition of BME increased the migratory rate of occludin knockdown cells only slightly (Figure 2A). There was, however, a significant reduction of the attachment of MDA-MB-231 control cells after treatment with BME (p<0.05) that was not seen in the occludin knockdown cells (Figure 2B).
Effect of BME and occludin knockdown in human cancer cell growth. We next investigated the effect of BME on the growth of two metastatic cancer cell lines, MDA-MB-231 (as above) and PC-3 prostate cancer cells with and without occludin knockdown. In MDA-MB-231 cells, there was a concentration-dependant effect on growth with BME. Control cells showed an increase in growth until a concentration of 30% BME was used. Conversely, all concentrations of BME caused reduced growth in occludin-knockdown cells (Figure 2C). The same effect was observed in PC-3 cells, albeit at a lower concentration of 10% BME (Figure 2D).
BME is able to effect changes in TJ function. As occludin is a key protein for correct TJ assembly and function, we examined the effect of BME on TER, a measure of TJ function. We used the MDA-MB-231 and PC-3 cells previously described, and in addition, three human lung cancer cell lines (A-549, SKMES-1 and CORL-23). Figure 3A shows the effect of 5% BME on TER of these cell lines. Interestingly, there was a clear difference regarding the response to BME. The MDA-MB-231 breast cancer cells only demonstrated an increase in resistance after 12 h of co-culture with BME, which was followed by a sharp decrease by 24 h. PC-3 cells showed a rapid increase in resistance after only 2 h of co-culture, which again decreased by 24 h. The lung cancer cell line A-549 showed a steady increase in TER over the 24-h time period, as did the CORL-23 cells. Conversely, the SKMES-2 lung cancer cell line showed a significant decrease in resistance with 2-h co-culture (p<0.04), with a recovery by 24 h. Figure 3B illustrates these data at the 24-h mark, with basal versus BME co-culture. We then returned to ECIS technology to assess barrier function after co-culture with BME (Figure 3C). Overall, BME was unable to modulate the resistance of occludin-knockdown MDA-MB-231 cells, whereas it was able to reduce the resistance and hence barrier function of control cells (p<0.05).

The loss of expression of occludin is related to metastatic disease in patients with breast cancer. Expression of occludin according to site of metastasis in the whole cohort (A) and in patients with ductal carcinoma (B). (C) Survival of patients with and without bone metastasis.
Discussion
Herein for the first time, we demonstrated that the loss of occludin is linked to an increase in breast cancer metastasis, specifically to the bone. Furthermore, we have shown that knockdown of occludin in metastatic breast and prostate cancer cells leads to a differential response to BME, suggesting that occludin can be modulated by the bone environment. We have also shown that BME can have a significant effect on the barrier function or resistance of cancer cell lines.
Occludin was the first identified integral protein for the TJ, and its long COOH-terminal domain is considered to have functions in receiving and transmitting cell survival signals, moreover, the loss of TJ molecules, such as occludin, has been correlated with tumor progression in carcinogenesis (13). We have previously shown that that occludin is differentially expressed in breast tumour tissues and in human breast cancer cell lines (10). The loss or truncation of the N-terminus indicates reduced assembly of the TJ structure and the reduced maintenance of barrier function. The loss of C-terminal expression suggests the reduced intracellular trafficking of occludin to the basolateral membrane and binding to ZO-1, resulting in reduced TJ anchoring, assembly and cell–cell adhesion (10). The knockdown of endogenous occludin in human breast cancer cells can lead to the profoundly reduced junctional integrity, loss of tightness and increased invasive potential of these cells (10).
In the present study, we showed that BME can cause a significant reduction of attachment of MDA-MB-231 cells that was not seen in the occludin-knockdown cells. Interestingly, the BME was able to cause a reduction in barrier function of breast cancer cells, but not the knockdown cells. This suggests that there are factors in the BME that are able to effect changes of the TJ of these cells via occludin. The differential response of the cancer cell lines to BME is difficult to explain and requires additional scrutiny. Breast cancer-derived bone metastases are osteoclastic, and prostate cancer-derived bone metastases are predominantly osteoblastic, whereas lung cancer-derived bone metastases are predominantly osteolytic, which may be a reason why there was such a differential response. Disseminated cells originating from breast, prostate and lung tumors can frequently be detected in the bone marrow with an extended latency at pre-metastatic sites, which may last for several years, being noted for breast and prostate cancer (8).
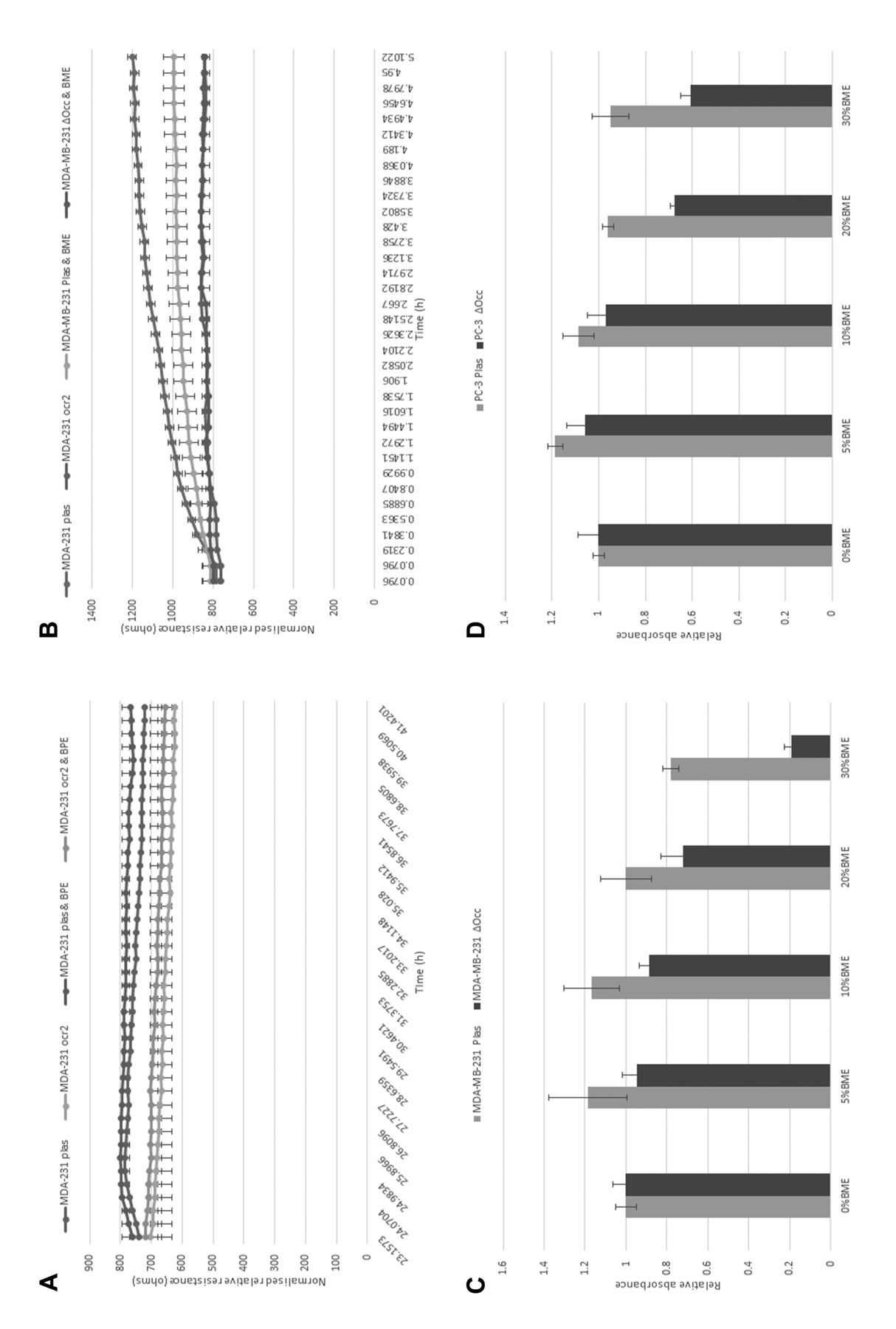
The effect of bone matrix extract (BME) on the migration, attachment and growth of metastatic cancer cells. (A) Shows the migration of MDA-MB-231 human breast cancer cells with or without BME. (B) Shows the effect of BME on attachment for these cells. The effect of BME on the growth of MDA-MB-231 and PC-3 cells is shown in (C) and (D).
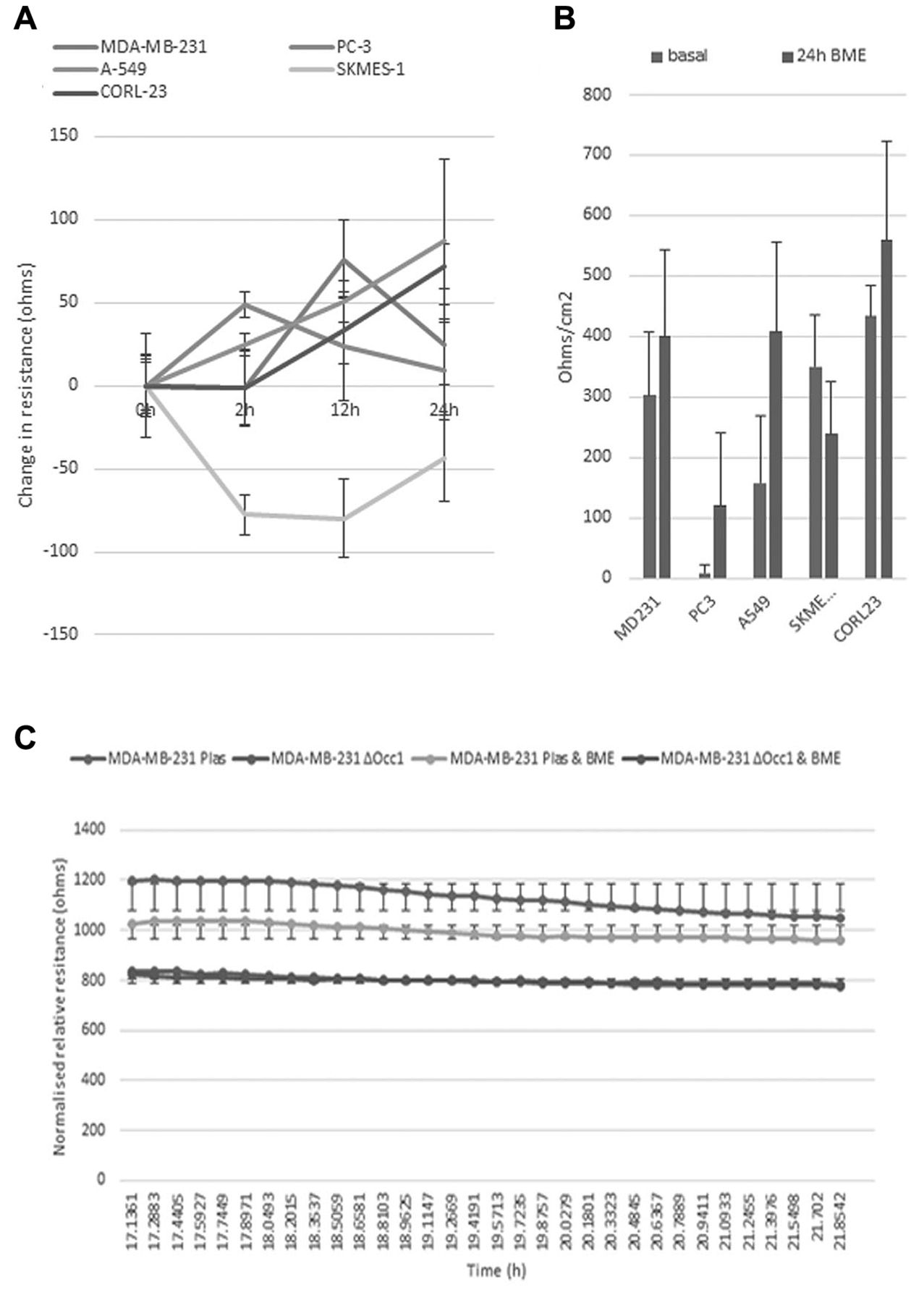
The effect of bone matrix extract (BME) on TJ function assessed using TER and ECIS. (A) Shows that 5% BME is able to effect changes in resistance of a number of human cancer cell lines over 25 hours. The difference between BME-treated and basal level cell to cell resistance is shown in (B). The change in barrier function of MDA-MB-231 cells is shown in (C).
In conclusion, the loss of expression of occludin may be involved in the success or failure of cancer cells to metastasize to the bone. The differential response to BME, suggests that occludin can be modulated by the bone environment and that cancer cells lacking occludin will have an increased propensity for bone metastasis.
Acknowledgements
The Authors wish to the thank Cancer Research Wales for supporting this work.
- Received January 4, 2016.
- Revision received February 5, 2016.
- Accepted February 18, 2016.
- Copyright© 2016 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
In this issue





{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.