


Abstract
Aim: This study aimed at investigating the anti-invasive activities of α-mangostin on human melanoma SK-MEL-28 and squamous cell carcinoma A-431 cell lines. Materials and Methods: Cytotoxicity was tested by the crystal violet assay; anti-invasive activity was detected by the wound healing, cell-matrix adhesion, and boyden chamber assays; and gene regulatory effects by qRT-PCR. Treatments were at non-toxic concentrations (0-1.25 μg/ml for A-431 cells and 0-2.5 μg/ml for SK-MEL-28 cells). Results: α-Mangostin inhibited motility, adhesion, migration and invasion. Invasive ability was reduced to 4% and 20% following α-mangostin treatment compared with untreated A-431 and SK-MEL-28 cells, respectively. Inhibition of gene expression of MMP-2, MMP-9, NF-κB, and Akt1 was involved in the anti-invasive activities on A-431 cells. Inhibition of MMP-2, NF-κB and IκBα was involved for SK-MEL-28 cells. Conclusion: α-Mangostin suppressed the metastatic processes of SK-MEL-28 and A-431 cell lines by differentially regulating metastasis-related genes, showing potential as an anti-metastatic agent.
Metastasis is the major cause of cancer treatment failure, being responsible for 90% of human cancer-related deaths (1). Skin cancer is a disease of concern because of its increasing incidence (2, 3). Melanoma and squamous cell carcinoma are the two major types of skin cancers (2, 4, 5). Melanoma (approximately 5% of total skin cancers) has a high tendency to metastasize. In patients with advanced metastatic melanoma, the 10-year survival rate is less than 10% (6-8). Squamous cell carcinoma (15-20% of total skin cancers) is also invasive and metastatic, even though the metastasis rate is much lower than that of melanoma (9, 10). Therefore, blocking cancer cell invasion and metastasis are critical targets for the therapeutic treatment of skin cancers.
Many plant-derived natural products have been reported to possess the capacity to inhibit melanoma metastasis. These natural products include 4-nerolidylcatechol (11), γ-tocotrienol (12), curcumin (13, 14), and aqueous extract of the root of Platycodon grandiflorum (15). However, limited information is available on the anti-metastatic activity of natural products on squamous cell carcinoma.
The natural compound investigated in this study was α-mangostin. α-Mangostin, a xanthone compound isolated from the pericarp of mangosteen, has been reported to exert cytotoxic effects on a variety of cancer cells (e.g. human breast cancer, colorectal cancer, hepatoma, leukemia, and small-cell lung cancer) (16-20). Moreover, we have reported that α-mangostin showed an apoptotic effect on human melanoma SK-MEL-28 cell line via activation of the caspase enzymes and disruption of the mitochondrial membrane (21). In addition to cytotoxicity, α-mangostin exhibited anti-metastatic activity on human prostate cancer PC-3, lung adenocarcinoma A-549, breast adenocarcinoma MCF-7 cells through inhibition of MMP-2, MMP-9 and NF-κB (22-24). α-Mangostin was recently reported to reduce tumor growth and lymph node metastasis in an immunocompetent xenograft model of metastatic mammary cancer (25). Panaxanthone (approximately 75% to 85% α-mangostin and 5% to 15% γ-mangostin) showed significant suppression of tumor growth and metastasis in a mouse model of mammary cancer, when administered in the diet (26). The anti-metastatic activities of α-mangostin have been found to be cancer-cell type-dependent because different sensitivities and mechanisms have been observed after treatment with α-mangostin on different cell lines. For example, treatment with α-mangostin inhibited the c-Jun NH2-terminal kinase (JNK) 1/2 pathway but not the extracellular signal-regulated kinase (ERK) 1/2 pathway in PC3 cells (22). In contrast, treatment with α-mangostin inhibited the ERK1/2 pathway but not the JNK1/2 pathway in A549 (24) and MCF-7 cells (23). Due to these differences, questions regarding the roles of α-mangostin in the metastasis of human skin cancer cell lines and its potential underlying mechanisms could not be answered based on currently available literature.
Primer sequences used for target genes amplification.
The present study was therefore designed to investigate the anti-metastatic effects of α-mangostin on two distinctive types of skin cancer cell lines (human squamous cell carcinoma A-431 and melanoma SK-MEL-28), by examining the multiple mechanisms of in vitro motility, adhesion, migration, invasion, and the ability to regulate the metastasis-related gene expression.
Materials and Methods
Materials. α-Mangostin was purchased from Phenomenex Australia Pty Ltd. (NSW, Australia). Bovine serum albumin (BSA), crystal violet, 4’, 6-diamidino-2-phenylindole dihydrochloride (DAPI), and trypsin-EDTA were purchased from Sigma-Aldrich (St. Louis, MO, USA). Fibronectin and type I collagen were from BD Biosciences (Bedford, MA, USA).
Cell lines and cell culture. Human metastatic melanoma SK-MEL-28 and human squamous cell carcinoma A-431 cell lines were purchased from the American Type Culture Collection. A-431 cells overexpress EGFR and 12-lipoxygenase, indicating the potential of invasion and metastasis (27, 28). Cells were cultured in DMEM (Sigma, USA) supplemented with 10% heat-inactivated fetal bovine serum (FBS; Invitrogen Corporation, Australia) in the presence of 100 U/ml penicillin and 0.1 mg/ml streptomycin (Thermo Scientific, Australia) and maintained in an incubator with 5% CO2, at 37°C. Cells under the passage number of 20 were used in experiments and cells were free of mycoplasma contamination, as detected by PCR (data not shown). Treatment preparation. α-Mangostin was dissolved in absolute ethanol and diluted with medium to the desired treatment concentration. Vehicle volume was consistently kept as 1% (v/v) for each treatment.
Cell viability assay. The cytotoxicity of α-mangostin was determined by the Crystal Violet assay, as described previously (21). Briefly, cells were seeded at 104 cells/well (6 wells per treatment condition) in 96-well plates and incubated for 24 h. Cells were then treated with α-mangostin at various concentrations (0, 0.3215, 0.625, 1.25 and 2.5 μg/ml) for 48 h of exposure duration. After treatment, cell culture medium was removed and cells were stained by 0.5% (w/v) crystal violet and destained with 33% (v/v) acetic acid. The absorbance was read at wavelength of 570 nm, on a μQuant automatic spectrophotometer (Bio-TEK instruments, NSW, Australia) with the KC Junior software (Bio-Tek, VT, USA). The cytotoxic effect of α-mangostin was presented as the percentage of cell viability compared to the untreated control cells.
Wound healing assay. A wound healing assay was performed to determine the cell motility, as previously described (29), with minor modifications. Briefly, SK-MEL-28 and A-431 cells (5×105 cells/well) were seeded in a 6-well tissue culture plate and grown to 80-90% confluence. After the medium was removed, a gap with constant width was created in the center of cells by scratching the monolayer with a sterile yellow micropipette tip. After that, cells were rinsed with phosphate-buffered saline (PBS) twice to get rid of cellular debris, and then cells were exposed to various concentrations of α-mangostin (0, 0.625 and 1.25 μg/ml for A-431; 0, 1.25 and 2.5 for SK-MEL-28). The wound closure was monitored and photographed at 0, 24, and 48 h with the Olympus 1×71 phase contrast inverted fluorescence microscope and the analySIS image capture software (magnification ×100). For each well, at least ten different areas of the scratch were photographed.
Boyden chamber invasion and migration assay. An in vitro invasion assay was conducted using a 24-well transwell unit (8 μm of pore size) with polyvinylpyrrolidone-free polycarbonate filters coated with Matrigel and placed in transwell chambers according to the manufacturer's instructions (BD Biosciences, Bedford, MA, USA). The coated filters were rehydrated using serum-free medium for 2 h in the incubator at 37°C before use. Medium containing 10% (v/v) FBS and 10 μg/ml of fibronectin was applied to the lower chamber as chemoattractant. Cells (2×104 cells/well) in serum-free medium were seeded in the upper part of the transwell plate and incubated with various concentrations of α-mangostin for 48 h at 37°C. After incubation, the cells in the upper surface of the membrane were carefully removed with a cotton swab and cells that invaded across the Matrigel to the lower surface of the membrane were fixed with methanol and stained with 1 μg/ml of DAPI. The invasive cells on the lower surface of the membrane filter were recorded with an Olympus 1×71 inverted fluorescence microscope and the analySIS image capture software, at magnification 200× and the cell number was counted by the Image J software. The data are presented as the average number of cells attached to the bottom surface from 10 randomly chosen fields. The experiments were repeated three times and each experiment was carried out in duplicate.
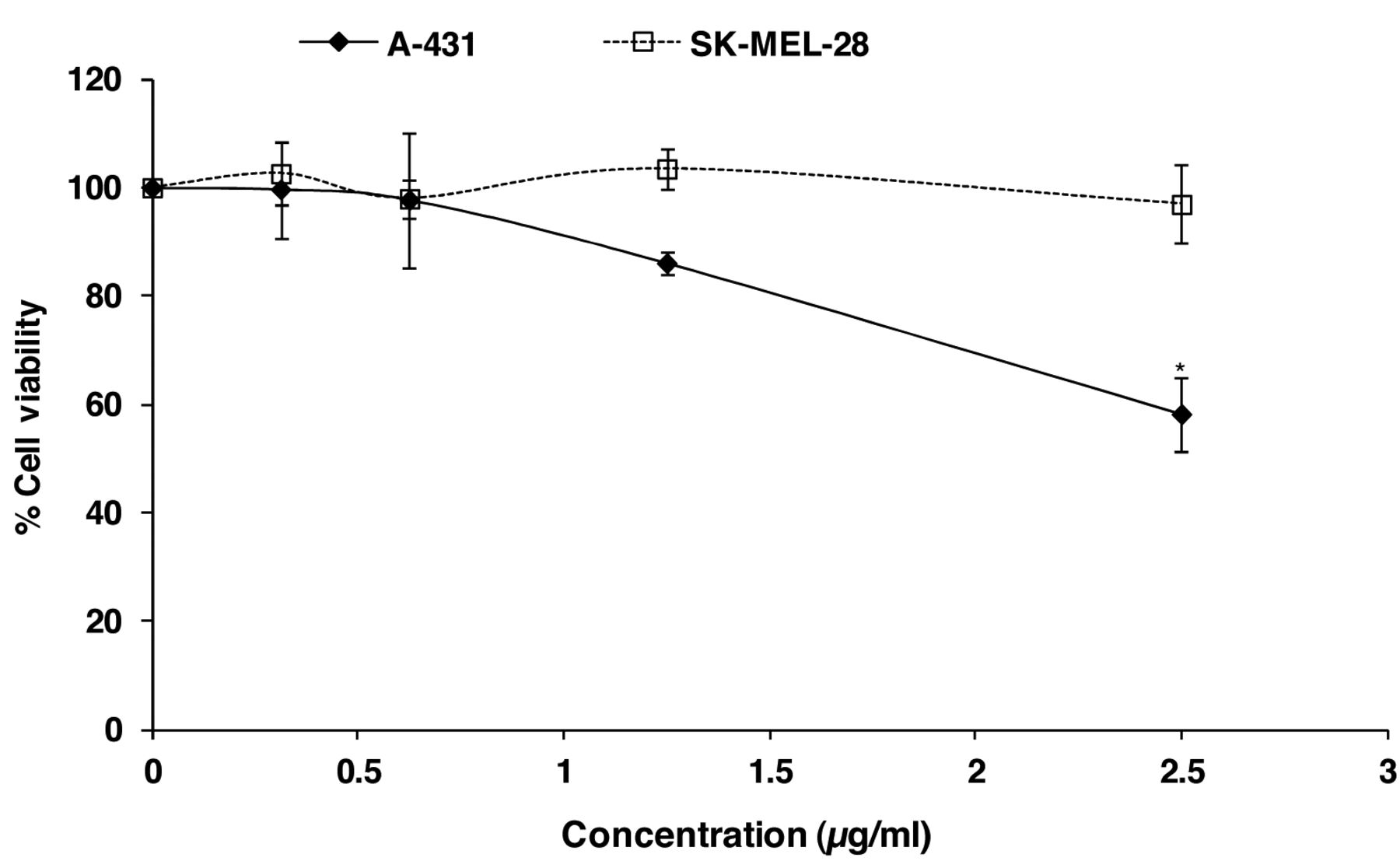
Effect of α-mangostin on the cell viability was determined by the crystal violet assay after 48-h treatment of A-431 and SK-MEL-28 cells with various concentrations of α-mangostin. Data are shown as % viability compared to the untreated control and are presented as the mean±SEM of three independent experiments. Treatments significantly different from the untreated control at p<0.05 are presented as * and at p<0.01 as **.
The migration assay was performed in a similar way to the invasion assay, except no Matrigel coating was applied to the upper surface of the transwell filters.
Cell-matrix adhesion assay. The cell adhesion assay was performed as previously described (30). 96-well cell culture plates were coated with type I collagen or fibronectin (200 ng/well) for 2 h at 37°C, followed by blocking with BSA (1 mg/ml) in PBS for 2 h at 37°C and rinsing twice with PBS. After pre-treatment with α-mangostin for 48 h, cells were detached with trypsin-EDTA. The suspended cells were washed once with DMEM culture medium containing 10% (v/v) FBS and twice with serum-free medium before being added to the wells at a density of 4×104 cells/well in 100 μl of serum free medium. Subsequently, the cells were incubated in the incubator for 1 h. After that, floating cells were removed by PBS washes, and adherent cells were stained with 0.5% (w/v) crystal violet and destained with 33% (v/v) acetic acid. The absorbance was recorded on a microplate reader at the wavelength of 570 nm. The effect of α-mangostin on cell adhesion was presented as the relative percentage of attached cells compared to the untreated control cells, as calculated by the following formula:

Quantitative real-time polymerase chain reaction (qRT-PCR). qRT-PCR was performed as described previously by Wang et al. (31) to determine mRNA expressions of different key genes involved in the process of metastasis in the A-431 and SK-MEL-28 cells subjected to the indicated treatments. The primers used are listed in Table I.
Statistical analysis. Data are expressed as mean (±SEM). The experiments were repeated at least three independent times. Statistical analysis of the data was carried out using ANOVA, followed by Tukey's HSD post hoc test (equal variances) or Dunnett's T3 post hoc test (unequal variances). These tests were performed using the SPSS software (version 17). Difference was considered statistically significant when the p-value was less than 0.05 (significant) or 0.01 (highly significant).
Results
The cytotoxicity of α-mangostin on skin cancer cells. The cytotoxicity of α-mangostin was previously demonstrated to be dose-dependent for the SK-MEL-28 cell line (21) and the A-431 cell line (data not shown). α-Mangostin showed less cytotoxicity on non-cancerous human skin-origin cells (primary skin fibroblast CCD-1064Sk) (21). The viability of skin cancer cells was, however, above 80% when treated with α-mangostin at a range of low concentrations (from 0 to 2.5 μg/ml for SK-MEL-28 and from 0 to 1.25 μg/ml for A-431) (Figure 1). To ensure the anti-metastatic activity of α-mangostin is not due to its cytotoxicity, these non-toxic doses were therefore used for all the subsequent experiments on the two types of skin cancer cells for the current study of anti-metastatic activities.

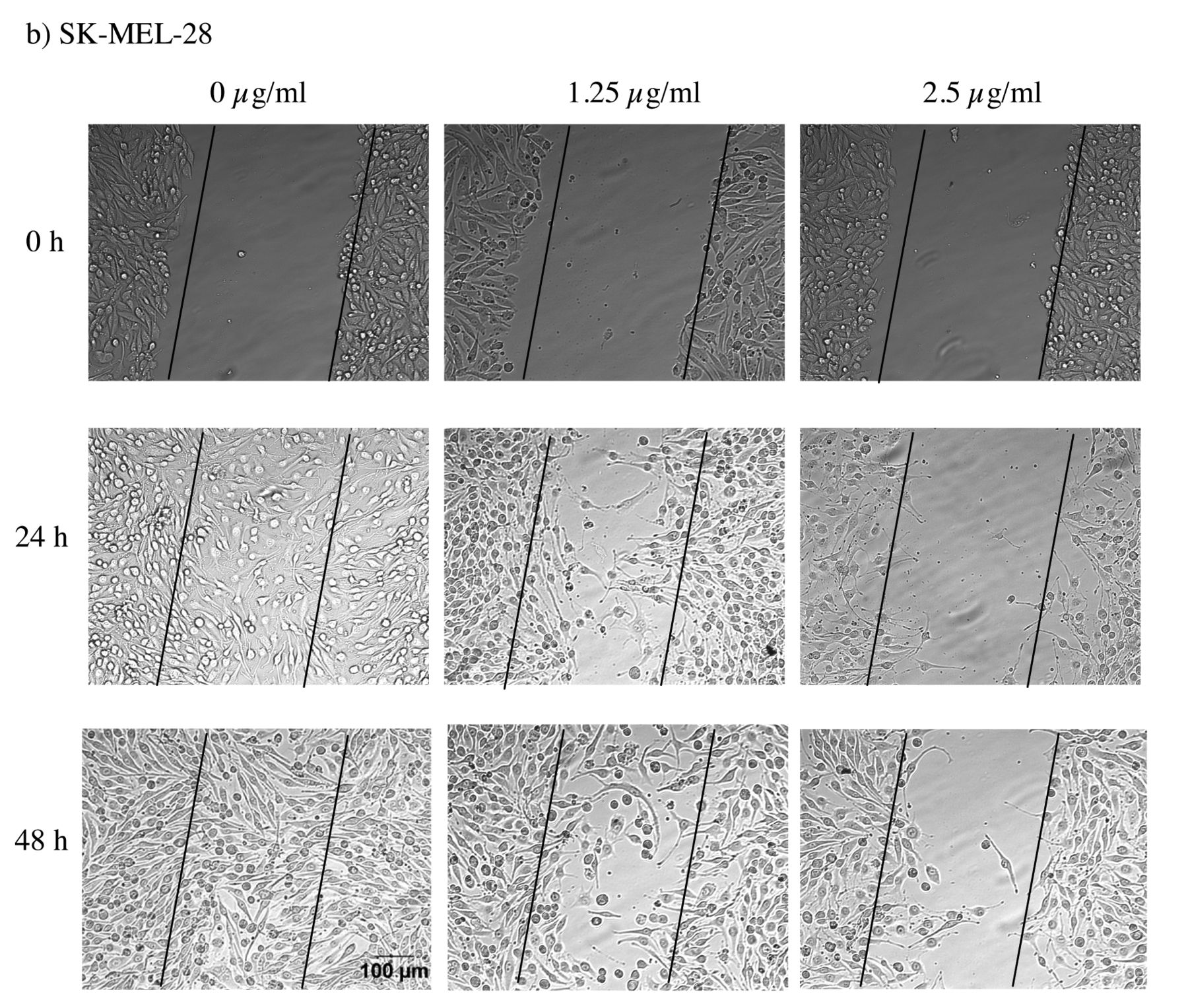
Effect of α-mangostin on the motility of a) A-431 and b) SK-MEL-28 cells was monitored by the wound healing assay as described in Materials and Methods. The wound closure was recorded at 0, 24, and 48 h by the Olympus 1X71 inverted fluorescence microscope and the analySIS image capture software. The images shown here are from a representative experiment repeated three times with similar results (scale bar: 100 μm).
α-Mangostin inhibits the motility of skin cancer cells. α-Mangostin displayed an inhibitory effect on cell motility for both A-431 and SK-MEL-28 cells after 24 h and 48 h of co-incubation at two different concentrations (Figure 2a and b), as evidenced by the lesser wound closure after treatment, compared with the untreated control. The inhibitory effect was found to be concentration-dependent. This was most marked for the concentration of 1.25 μg/ml for A-431 and 2.5 μg/ml for SK-MEL-28.
α-Mangostin inhibits the migration and invasion of skin cancer cells. Boyden chamber assay was used to measure the migratory and invasive potential of skin cancer cells after 48-h treatment with α-mangostin. α-Mangostin induced a dose-dependent decrease in migration and invasion with increasing concentration of α-mangostin on both A-431 and SK-MEL-28 cell lines (p<0.01; Figure 3a and b). After treatment of A-431 cells with 1.25 μg/ml of α-mangostin, the migration and the invasion were reduced to 6% and 4% of the untreated control, respectively (Figure 3a). After treatment of SK-MEL-28 cells with 2.5 μg/ml of α-mangostin, the migration and the invasion were reduced to 23% and 20% of the untreated control (Figure 3b).
α-Mangostin inhibits the adhesion of skin cancer cells. As detected by the cell-matrix adhesion assay, α-mangostin was found to decrease the adherence of the skin cancer cells to fibronectin and type I collagen. The adhesive capability on fibronectin decreased to 54% for A-431 cells after treatment with 1.25 μg/ml α-mangostin (p<0.05), and to 63% for SK-MEL-28 cells after treatment with 2.5 μg/ml α-mangostin (p<0.05). The adhesion ability on type I collagen decreased to 57% at 1.25 μg/ml of α-mangostin for A-431(p>0.05) and 59% at 2.5 μg/ml of α-mangostin for SK-MEL-28 (p<0.01), respectively (Figure 4).
α-Mangostin modulates the metastasis-related genes of skin cancer cells. As detected by qRT-PCR, α-mangostin was observed to inhibit metastasis of the skin cancer cells by down-regulating the target genes (Figure 5a and b). In A-431 cells, compared with untreated control, a 1.8-, 2.2-, 1.9-, and 2.8-fold decrease was observed after treatment with α-mangostin in the mRNA expression of MMP-2 (p<0.05), MMP-9 (p<0.05), NF-κB (p<0.05), and Akt1 (p<0.01), respectively (Figure 5a). The mRNA expression of IκBα was not significantly changed after treatment with α-mangostin in this cell line (Figure 5a). In SK-MEL-28 cells, treatment with α-mangostin induced a 3.6-, 3.2-, and 3.2-fold decrease in the mRNA expression of MMP-2 (p<0.05), NF-κB (p<0.05), and IκBα (p<0.05), respectively (Figure 5b). α-Mangostin also inhibited the mRNA expression of MMP-9 and Akt1, but this was not statistically significant (Figure 5b).
Discussion
The metastasis of cancer cells is a complex multistep process, including altered adhesion ability between cells and the extracellular matrix (ECM), degradation of the environmental ECM and basement membrane, detachment of cancer cells from the primary site, invasion into new blood vessels, and establishment of a new tumor at distant sites (32-34). Disruption of any of these metastatic components is a target for anti-metastasis therapeutics (35, 36). Based on the current results, the potential mechanisms of α-mangostin induced anti-metastatic activity are depicted in Figure 6. The changes in gene expression or metastatic activities are indicated. It is acknowledged that there are additional genes involved in some of these pathways. These are yet to be fully elucidated.
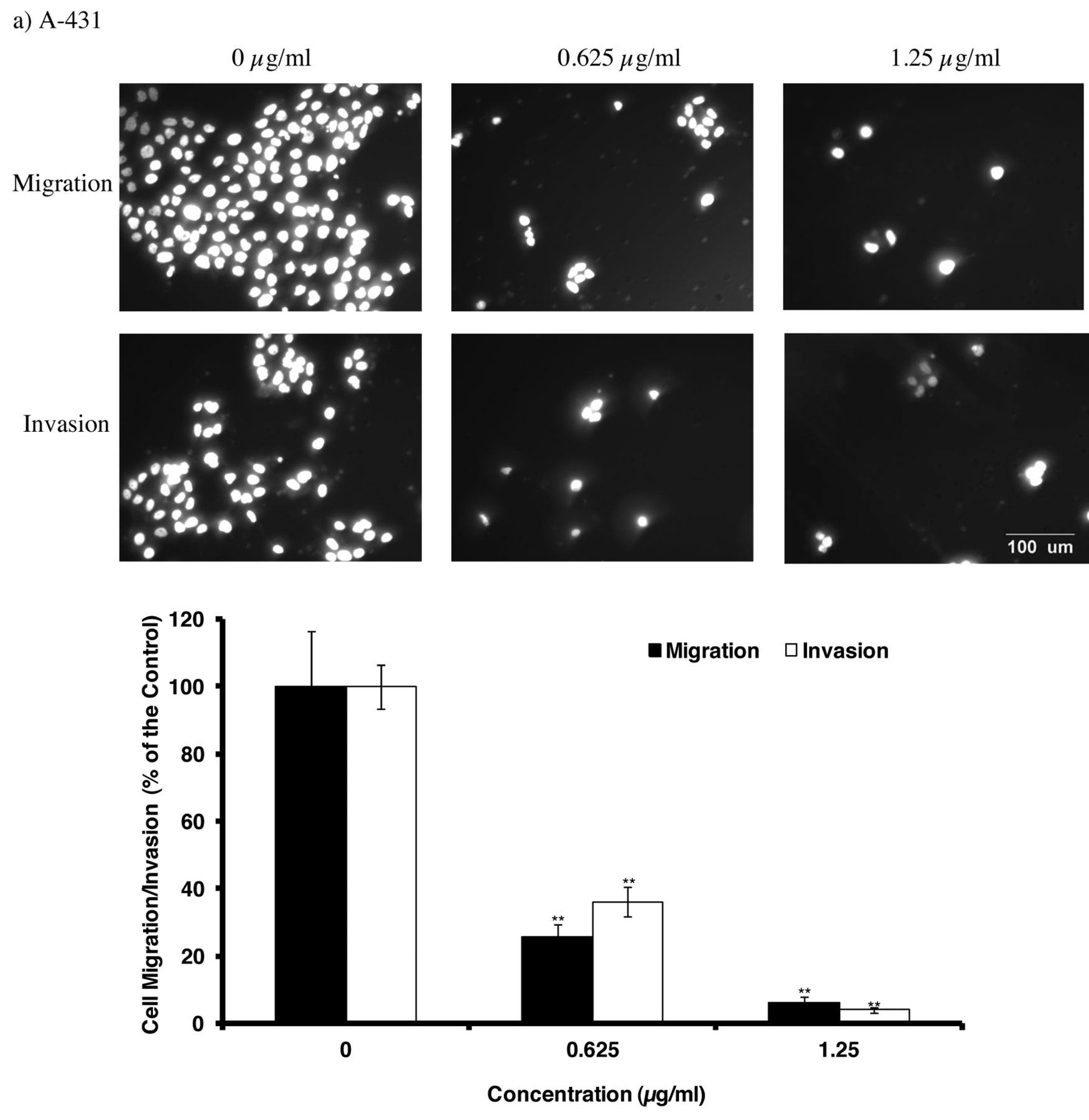

Effect of α-mangostin on migration and invasion of a) A-431 and b) SK-MEL-28 cells was detected by the Boyden Chamber assay after 48-h treatment, as described in Materials and Methods. The migration and invasion of cells were recorded by the Olympus 1X71 inverted fluorescence microscope and the analySIS image capture software and the cell number was counted by the Image J software (scale bar: 100 μm). The values are shown as the mean±SEM of three independent experiments. Treatments significantly different from the untreated control at p<0.05 are presented as * and at p<0.01 as **.
The present study demonstrated that α-mangostin inhibited the motility, migration and invasion of the two skin cancer cell lines at non-toxic concentrations, in a dose-dependent manner (Figures 2 and 3). The current results are consistent with our study, α-mangostin inhibits the motility of human prostate cancer PC-3 and breast cancer MCF-7 cells (22, 23). Migratory and invasive ability are important characteristics of metastatic cancer cells. The current results showed that α-mangostin induced a dose-dependent decrease in migration and invasion with increasing concentration of α-mangostin (Figure 4; p<0.01). In agreement with our study, α-mangostin inhibited migration and invasion of human prostate cancer PC-3 cell line, breast cancer MCF-7 cell line, and lung cancer A-549 cell line (22-24). However, these activities were found to be cell type-dependent. For example, in our study, α-mangostin at 1.25 μg/ml reduced the invasion to 4% and migration to 6% in A-431, and at 2.5 μg/ml reduced the invasion to 20% and migration to 23% in SK-MEL-28. In the previous study, at 2.1 μg/ml (in A-549 cells) reduced the invasion to 20% and migration to 40%, at 5 μg/ml (in PC-3 cells) it reduced the invasion to 60% and migration to 40%, and at around 2.5 μg/ml (in MCF-7 cells), it reduced the invasion to 55% and migration to 61% compared with the untreated controls (22-24). Therefore, α-mangostin appears to be more effective on the two skin cancer cells, especially A-431 cells, compared to other cancer cell types.

Effect of α-mangostin on the adhesion of a) A-431 and b) SK-MEL-28 cells was evaluated by the cell-matrix adhesion assay. After 48-h treatment with or without α-mangostin, cells were seeded to fibronectin or collagen-coated wells. The cell-matrix adhesion assay was carried out as described in Materials and Methods. Data are shown as % relative attached cells compared to the untreated control. The values are shown as the mean±SEM of three independent experiments. Treatments significantly different from the untreated control at p<0.05 are presented as * and at p<0.01 as **.
The attachment of cancer cells to ECM is an important step in the process of metastasis (32, 37). To further investigate the possible mechanism of anti-metastatic effects of α-mangostin, the effect of α-mangostin on the ability of A-431 and SK-MEL-28 to adhere to fibronectin and type I collagen, the major components of ECM, was examined. α-Mangostin significantly decreased the adhesive ability on fibronectin on the two skin cancer cell lines. However, α-mangostin only significantly inhibited the adhesive capability on type I collagen on SK-MEL-28 cells, but not on A-431 cells. Consistent with our study, α-mangostin inhibited the adhesion to type I collagen of PC-3 (22) and MCF-7 cells (23) with a decrease to around 75%, at 5 μg/ml, and around 40%, at 2.5 μg/ml, respectively. An increased capacity for collagen binding is one of the important features of metastatic cancer cells (38). Therefore, the inhibition of type I collagen cell adhesion by α-mangostin may indicate its anti-invasive effects. Although the role of fibronectin in the metastasis of skin cancer cells is not clear, it is likely that the inhibition of fibronectin cell adhesion contributed to the anti-invasive effects induced by α-mangostin. In the future, the effect of α-mangostin on other ECM proteins (e.g. laminin, type IV collagen, and vitronectin), as well as on the integrins which mediate the ECM protein adhesion, should be studied.
Degradation of the ECM components is a key step for the metastasis process. Matrix metalloproteinases (MMP)-2 and MMP-9 are highly expressed in various malignant tumors and activation of these two enzymes is required for the degradation of ECM (39, 40). Therefore, inhibition of MMPs is an important target for the anti-metastasis therapy. To further explore the exact mechanism of α-mangostin for inhibition of invasion and migration, qRT-PCR was performed to detect the mRNA levels of MMP-2 and MMP-9. In this study, we found that α-mangostin could significantly down-regulate the mRNA levels of MMP-2 on both skin cancer cell lines, while that of MMP-9 only had an effect on A-431 cells. Likewise, inhibition of MMP-2 and MMP-9 by α-mangostin was also observed on PC-3, MCF-7, and A549 cells at different concentrations, as detected by various methods, such as gelatine zymography, western blotting, and RT-PCR (22-24). It would be interesting as a future study to evaluate the production of interleukine-8 from these two skin cancer cell lines due to its important role in regulation of the expression of MMP-2 and MMP-9 (41).
Elevated activity of NF-κB was shown to stimulate cell migration and angiogenesis by increasing expression of interleukin-8 and vascular-endothelial growth factor (42, 43). Additionally, the MMP-2 and MMP-9 genes were reported to be regulated through the transcriptional level interaction of NF-κB with their binding sequences in the MMP-2 and MMP-9 gene promoter (44). Therefore, NF-κB is an important target for inhibiting metastasis of cancer cells (44, 45). In the current study, down-regulation of NF-κB mRNA expression was found after treatment with α-mangostin on both types of skin cancer cells studied. Similar findings on other cell lines were reported in previous studies (22-24). Moreover, an increase in the mRNA level of IκBα was found after treatment with α-mangostin on A-431 cells (Figure 5a; p>0.05). This was consistent with the study of Lee et al. (23), which reported that α-mangostin induced an increase in the protein level of IκBα as detected by western blotting. In contrast, a significant decrease in the mRNA expression of IκBα was observed after treatment of SK-MEL-28 cells with α-mangostin (Figure 5b; p<0.05). The relationship between NF-κB and IκBα is complex (46). IκBα can inhibit activation of NF-κB by trapping NF-κB in the cytoplasm. Meanwhile, IκBα is one of the major downstream targets of NF-κB, hence activation of NF-κB can switch on IκBα gene expression (46). Therefore, in SK-MEL-28 cells, suppression of NF-κB may lead to the down-regulation of IκBα. Additionally, a regulatory role of ERK in the expression of NF-κB has been reported (47). Therefore, apart from IκBα, the ERK pathway may also contribute to the down-regulation of NF-κB expression induced by α-mangostin. This requires further investigation.

Effect of α-mangostin on the metastasis-related gene expression was detected by qRT-PCR after 48-h treatment on a) A-431 and b) SK-MEL-28 cells. The values are shown as the mean±SEM of three independent experiments. Treatments significantly different from the untreated control at p<0.05 are presented as * and at p<0.01 as **.

Proposed model of mechanisms involved in α-mangostin-induced anti-metastatic effect on human skin cancer cells. The genes involved are indicated by the common abbreviations (Akt, protein kinase B; IκB, inhibitor of kappa B; IKK, inhibitor of kappa B Kinase; MMP, matrix metalloproteinase; NF-κB, nuclear factor kappa B). Adjacent to each gene is indicated the direction of expression level change as ↓=decreased expression, ⇅=increased and decreased expression, after treatment with α-mangostin on different cell lines. For motility, adhesion, migration, and invasion, the ↓ indicates a decrease in these activities.
Akt has been reported to be involved in the metastasis of various tumors (48, 49). In particular, activation of Akt is found in approximately two thirds of melanomas and regulates melanoma cell migration, invasion and metastasis (50-53). In this study, α-mangostin decreased the mRNA levels of Akt1 on both A-431 (p<0.05) and SK-MEL-28 (p>0.05) cells. This suggests that the inhibition of Akt1 was involved in the anti-metastatic activity of α-mangostin. Akt was reported to regulate NF-κB via activation of IKK and consequent release of NF-κB from the inhibitory complex (54, 55). The current results indicate that suppression of Akt contributes to the down-regulation of NF-κB induced by α-mangostin. Likewise, Shibata et al. (25) reported that phospho-Akt-threonine 308 was inhibited after treatment with α-mangostin in human mammary carcinoma MDA-MB231 cells. In contrast, Akt alterations were not found after treatment with α-mangostin of A-549, PC-3, and MCF-7 cells (22-24). It is possible that the effects of α-mangostin could partially exert through different molecular targets in different cancer cell lines. This phenomenon needs to be studied further.
There are some lines of investigation that could build on the current findings. The mRNA expression patterns, as detected by qRT-PCR, provide important information for the quantitative description of biological systems (56). The mRNA expression of the target gene is not indicative of the subcellular location of the protein (57-59). Hence, gene analysis in complement with proteome analysis is needed to assess the quantitative description of the state of a biological system (60). Proteome analysis includes the quantity of target protein expression, their subcellular location, and the modification state. Therefore, in the future, the results from the current study can be confirmed and clarified by proteome analysis (e.g. two-dimensional gel electrophoresis, western blotting, and ELISA). In addition, the current model for the metastasis assays is a simple two-dimensional one, but in the future, a three-dimensional model, constructed using isolated cutaneous cell populations could be used to better mimic the human disease than the two-dimensional invasion assay (61). Furthermore, animal models could be used to study skin cancer progression and spread, and provide an indication of the therapeutic efficacy of α-mangostin.
In conclusion, the present study demonstrates that α-mangostin inhibits the processes involved in the metastasis of A-431 and SK-MEL-28 cells, as reflected in results for cell motility, adhesion, migration, and Matrigel invasion. The anti-invasive activity of α-mangostin may involve inhibition of Akt1 and NF-κB, thereby decreasing the activities of MMP-2 and MMP-9. For the anti-invasive and anti-metastatic activities of α-mangostin, significant difference of anti-metastatic activity and mechanisms were found between different cancer cell types in previous studies and those of skin cancer in the current study. Thus, our study produced novel results that contribute new knowledge in this field by providing important evidence and mechanistic explanations regarding the anti-invasive activity of α-mangostin in the two tested skin cancer cell lines.
Acknowledgements
Jing J. Wang gratefully acknowledges Flinders University for awarding an EIPRS scholarship. The research was supported by Klein Research Institute Ltd, Adelaide, Australia and Flinders University Underwriting Grant. The Authors are thankful to George Mayne from the Department of Surgery, Flinders Medical Centre, for his kind help with qRT-PCR methods and analysis.
Footnotes
-
Conflicts of Interest
The Authors have declared no conflict of interest.
- Received June 12, 2012.
- Accepted July 13, 2012.
- Copyright© 2012 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}