


Abstract
Background: High invasiveness of glioma cells is one of the reasons that patients with malignant glioma have a poor prognosis. Resveratrol, a plant compound abundant in the peel of grapes, has been suggested as a potential cancer chemopreventive agent. Therefore, we investigated the effect of resveratrol on glioma cell invasion. Materials and Methods: The effect of resveratrol on U373MG human glioma cell invasion was assessed by Matrigel assay and methylthiazoltetrazolium assay. Western blotting and reverse transcription-polymerase chain reaction were performed to elucidate the action mechanism of resveratrol. Results: Resveratrol reduced tumor necrosis factor (TNF)-α-induced U373MG human glioma cell invasion. In addition, resveratrol repressed nuclear factor kappa B (NF-κB) activation and down-regulated mRNA expression of urokinase plasminogen activator (uPA) and its receptor in TNF-α-treated glioma cells. Conclusion: These findings suggest that resveratrol could prevent glioma cell invasion via inhibiting proteolysis of extracellular matrix.
Gliomas are one of the most common types of malignant brain tumors (1). In particular, glioblastoma multiforme (GBM), WHO grade IV, is highly invasive (2). Because of its high invasiveness, patients with unresectable GBM have a poor prognosis and have median survival of only a few months (3).
Inflammatory cytokines play an important role in cancer progression in many types of malignant tumors (4). During tumor progression, changes of the microenvironment could elicit cytokine expression that stimulates leukocytes, leading to additional cytokine release. Tumor necrosis factor-α (TNF-α) is a pro-inflammatory cytokine whose signaling pathways are associated with both pro-apoptotic and anti-apoptotic responses (5, 6). Although TNF-α causes apoptosis in several tumor types (7), most glioma cells are resistant to TNF-α-induced apoptosis (8). Interestingly, motility and invasion of glioma cells are promoted through the activation of nuclear factor kappa B (NF-κB) by constitutively expressed TNF-α (9). Both induced and constitutively expressed NF-κB has influence on cancer progression and metastasis (10-12). The role of NF-κB induced by TNF-α has been intensively studied as a target of novel cancer therapeutics (13, 14).
The invasiveness of glioma depends on proteolysis of the extracellular matrix, a complex event occurring through the surrounding tissue during tumor cell invasion. Urokinase plasminogen activator (uPA) and its specific receptor (uPAR) play a major role in the infiltrative growth of glioblastoma (15). Expression of genes of the uPA/uPAR system in humans has been reported to be under the regulation of extracellular mediators, such as growth factors and cytokines, as well as intracellular signaling through β-catenin and transcription factors, including NF-κB (16-18).
In numerous efforts to find novel anticancer agents, many plant extracts have been studied. Among them, resveratrol (trans-3,4’,5-trihydrostilbene), a polyphenol found in fruits and vegetables, has been proven to have antioxidant, anti-inflammatory, and antitumor properties (19-21), implying its potential as therapeutic supplement for cancer (22, 23). Indeed, resveratrol has a strong antiproliferative effect on many cancer cell types in vitro (24, 25) and inhibits the manifold process of carcinogenesis at diverse stages in vivo (22). Previous studies also report that resveratrol has an inhibitory effect on glioma (3, 26, 27), Furthermore, it has been shown that resveratrol can pass through the blood-brain barrier, reaching the brain tissues (28). These results suggest the potential of resveratrol as therapeutic supplement for brain tumors, although the effect of resveratrol on glioma is not fully understood. Hence, this study was conducted to investigate the effect of resveratrol on U373MG human glioma cell invasion.
Materials and Methods
Reagents. High glucose Dulbecco's modified Eagle's medium (DMEM), penicillin, streptomycin, and fetal bovine serum (FBS) were obtained from Gibco (Carlsbad, CA, USA). TRIzol Reagent and 4,6-diamidino-2-phenylindole (DAPI) were purchased from Invitrogen (Carlsbad, CA, USA). TNF-α, resveratrol, dimethylsulfoxide (DMSO), and methylthiazoltetrazolium (MTT) were purchased from Sigma (Saint Louis, MO, USA). Anti-phospho-NF-κB, and anti-NF-κB antibodies were purchased from Cell Signaling Technology (Danvers, MA, USA). Anti-α-tubulin antibody was purchased from Pierce (Rockford, IL, USA). Resveratrol was dissolved in DMSO, and MTT was dissolved in phosphate-buffered saline (PBS).
Cell culture and treatment. U373MG human glioma cells (ATCC, Manassas, VA, USA) were maintained in high glucose DMEM supplemented with 10% FBS, 100 U/ml penicillin, and 100 μg/ml streptomycin at 37°C in an atmosphere with 5% CO2. For some experiments, U373MG cells were cultured in DMEM with 0.1% FBS in the presence of designated concentrations of TNF-α (1-50 ng/ml) and/or resveratrol (5-100 μM). Cells were exposed to TNF-α for 24 h prior to cell viability assay and cell invasion assay.
Cell viability assay. U373MG cells (1×104 cells/well) were maintained in a 24-well plate overnight prior to treatment. Following treatment as described above, 100 μl of MTT solution (2.5 mg/ml) was added to each well, and the cells were incubated at 37°C for an additional 4 h. DMSO was added and mixed thoroughly. Absorbance was assessed at 570 nm with a microplate reader (Bio-Rad Laboratories, Hercules, CA, USA).
Cell invasion assay. Briefly, cells were trypsinized and seeded on Matrigel-coated Transwell inserts (Corning Incorporated, Steuben County, NY, USA) at a density of 1×105 cells/well in triplicates in 100 μl of DMEM with 0.1% serum, in the presence of TNF-α or a combination of TNF-α and Resveratrol. After 24 h incubation, the filter inserts were removed. Cells were fixed with methanol at −20°C for 5 min and washed three times with distilled water. Cells were then stained with DAPI (1 μg/ml) for 10 min and rinsed three times with distilled water. The cells on the upper side of the Transwell filter were removed with cotton swabs. The numbers of invading cells were quantified under a fluorescence microscope (Olympus, Tokyo, Japan).
Reverse transcription (RT)-PCR. Total RNA was extracted using TRIzol reagent following the manufacturer's instructions. Total RNA (1 μg) was used for cDNA synthesis using an iCycler thermocycler (Bio-Rad Laboratories). Primers were synthesized on the basis of the reported human cDNA sequences for uPA, uPAR, and hGAPDH in the NCBI data bank. Sequences of the primers used for PCR were as follows; uPA forward, 5’-ACTTCTCCAACATTCACTGG-3’; uPA reverse, 5’-ATTCTTCTGGAGGAGAGGAG-3’; uPAR forward, 5’-GTGCATGAVGTGTAAGACC-3’; uPAR reverse, 5’-CTCTCA CAGCTCATGTCTGA-3’; GAPDH forward, 5’-AGCTGAACGG GAAGCTCACT-3’; GAPDH reverse, 5’-TGCTGTAGCCAAAT TCGTTG-3’ (Bioneer Corporation, Daejeon, South Korea). uPA and uPAR cDNA amplification conditions were 94°C for 30 s, 55°C for 30 s, and 72°C for 1 min for 28 cycles. GAPDH cDNA amplification conditions were 94°C for 30 s, 55°C for 30 s, and 72°C for 1 min for 25 cycles. After amplification, PCR products were electrophoresed in agarose gel, stained with ethidium bromide and visualized with a UV transilluminator. The amplified DNA fragments were 360, 315 and 175 bp for uPA, uPAR, and GAPDH, respectively. The band intensities were assessed by SigmaGel software (Jandel Scientific, San Rafael, CA, USA).
Western blotting. U373MG cells were plated on 100 mm dishes at 1×106 cells/dish. Following treatment as described above, cells were harvested, centrifuged at 3×103 rpm for 1 min, and lysed in lysis buffer [50 mM Tris-Cl (pH 8.0), 150 mM NaCl, 1% NP-40, 0.5% sodium deoxycholate, 0.1% sodium dodecyl sulfate, 0.02% sodium azide, and proteinase inhibitor cocktail (phenylmethylsulphonyl fluoride, 100 μg/ml; aprotinin, 1 μg/ml; leupeptin, 0.5 μg/ml) (Roche, Nutley, NJ, USA)]. Total protein concentration was determined using BCA method (Pierce). An equal volume of 4× SDS sample buffer was added to 30 μg of protein extract and the samples were boiled for 5 min. Equivalent amounts of total protein (20-30 μg) were separated by SDS-PAGE on 8-12% polyacrylamide gel and then transferred to nitrocellulose membrane at 15 V for 40 min using a semi-dry transfer apparatus (Bio-Rad Laboratories) submerged in transfer buffer (25 mM Tris, 192 mM glycine, 20% methanol, pH 8.3). The membrane was blocked with 5% skim milk in 0.1% Tween-20/Tris-buffered saline (TTBS). The membrane was incubated with anti-phospho-NF-κB (1:1000), anti-NF-κB (1:1000), or anti-α-tubulin (1:10000) antibody at 4°C overnight. Subsequently, the blot was washed in TTBS and incubated with the appropriate horseradish peroxide-conjugated secondary antibodies for 30 min. After washing, immunoreactivity was detected by chemiluminescence (ECL; Amersham Biosciences, Piscataway, NJ, USA) using x-ray films. The band intensities were assessed by SigmaGel software (Jandel Scientific, San Rafael, CA, USA).
Statistical analysis. Data are expressed as the mean±SE Statistical significance was determined using the Student t-test (SigmaPlot; Jandel Scientific, Erkrath, Germany). P-values <0.05 were considered statistically significant.
Results
Resveratrol suppresses U373MG human glioma cell invasion. As resveratrol acts as an anticancer agent in various cancer cells (29, 30), we tested whether it has similar effect on glioma cell invasion and viability using Matrigel invasion assay and MTT assay. The results from the Matrigel invasion assay revealed that resveratrol treatment (5-20 μM) significantly inhibited U373MG human glioma cell invasion (Figure 1A and B). The numbers of invaded cells were reduced by approximately 15%, 30%, and 45% by 5 μM, 10 μM, and 20 μM resveratrol, respectively. However, the MTT assay revealed that there was no significant cytotoxic effect on U373MG cells by resveratrol at the concentrations tested (Figure 1C).
Resveratrol represses TNF-α-induced U373MG cell invasion. Inflammatory cytokines are associated with cancer cell invasion (31) and TNF-α is known to play an important role in glioma cell invasion (32). Among the concentrations tested, 10 ng/ml of TNF-α most effectively increased glioma cell invasion by up to approximately 2-fold, compared with vehicle control (Fig 2A and B). Thus, 10 ng/ml of TNF-α was used to induce glioma cell invasion for further study. Resveratrol significantly suppressed TNF-α-induced cell invasion at 5-20 μM in a dose-dependent manner (Figure 2D and E). As 10 μM resveratrol effectively reduced TNF-α-induced glioma cell invasion to a basal level, this concentration was used for further study. The results of the MTT assay showed that TNF-α, resveratrol alone, and combinations of TNF-α and resveratrol have no significant effect on U373MG human glioma cell viability (Figure 2C and F). These results along with the ones presented in Figure 1 suggest that resveratrol reduced not only intrinsic invasiveness but also TNF-α-induced invasion of glioma cells, without affecting cell viability.
Resveratrol reduces TNF-α-induced NF-κB activation in U373MG cells. Previous studies have revealed that NF-κB promotes cancer cell invasion (33, 34) and resveratrol down-regulates NF-κB activation in various cancer cells (35, 36). Thus, we examined whether resveratrol inhibited TNF-α-induced NF-κB activation in U373MG cells. As expected, TNF-α treatment enhanced NF-κB activation, which was detected as phosphorylated NF-κB (Figure 3A). The amount of increased phosphorylated NF-κB reached plateau in 1 h and did not significantly change by 24 h post TNF-α stimulation. Resveratrol (10 μM) reduced TNF-α-induced NF-κB phosphorylation by approximately 38%, 42%, and 58% at 1, 3, and 6 h following TNF-α stimulation, respectively (Figure 3B). In summary, resveratrol inhibited TNG-α-induced NF-κB signaling pathway in glioma cells
Resveratrol down-regulates TNF-α-induced uPA and uPAR mRNA expression in U373MG cells. Previous studies have shown that NF-κB is one of the upstream regulators of the plasminogen activator system (37). Thus, the effect of resveratrol on uPA and uPAR gene expression was assessed by RT-PCR. mRNA expression of both uPA and uPAR was significantly up-regulated by TNF-α (Figure 4A and B). uPA and uPAR transcripts appeared to rapidly increase in 1 h to 370% and 330%, respectively, compared with negative control (0 h) and to gradually decrease afterwards. Resveratrol (10 μM) down-regulated TNF-α-induced uPA mRNA expression by approximately 43% at an early time point (1 h), level at which the expression remained unchanged (Figure 4C and D). Down-regulation of uPAR expression by resveratrol appeared to occur slightly later so that uPAR transcripts decreased by 36% at 1 h and by 60% at 3 h following TNF-α and resveratrol treatment (Figure 4C and D). Taken together, the inhibitory effect of resveratrol on glioma cell invasion was mediated by inhibiting the NF-κB pathway, leading to down-regulation of uPA and uPAR gene expression, which are required for proteolysis of extracellular matrix.

Effect of resveratrol on U373MG human glioma cell invasion. A and B: U373MG cells were treated with resveratrol at the indicated concentrations for 24 h. Cell invasion was examined by Matrigel cell invasion assay. Data are presented as the mean±SE *p<0.05 (vs. CTL). CTL, Control; Res, resveratrol. C: U373MG cells were treated with resveratrol at the indicated concentrations for 24 h. MTT assay was performed to determine the cytotoxic effect of resveratrol. Cell survival relative to that of the control (%) is presented as the mean±SE (n=8).

Effect of resveratrol on TNF-α-induced U373MG cell invasion. A and B: U373MG cells were treated with TNF-α at the indicated concentrations for 24 h. Cell invasion was assessed by Matrigel cell invasion assay. Data are presented as the mean±SE. *p<0.05 (vs. CTL). C: U373MG cells were treated with TNF-α at the indicated concentrations for 24 h. MTT assay was performed to determine the cytotoxic effect of TNF-α. Cell survival relative to that of the control (%) is presented as the mean±SE (n=8). D and E: U373MG cells were treated with TNF-α (10 ng/ml) in the presence of resveratrol (5, 10, and 20 μM) for 24 h. Matrigel cell invasion assay was performed to determine the effect of resveratrol on cancer cell invasion. (i) CTL, (ii) TNF-α (10 ng/ml), (iii) TNF-α (10 ng/ml) + resveratrol (5 μM), (iv) TNF-α (10 ng/ml) + resveratrol (10 μM), and (v) TNF-α (10 ng/ml) + resveratrol (20 μM). Data are presented as mean±S±SE *p<0.05 (vs. TNF-α). F: U373MG cells were treated with TNF-α in the presence of resveratrol at the indicated concentrations for 24 h. MTT assay was performed to determine the cytotoxic effect of TNF-α and resveratrol. Cell survival relative to that of the control (%) is presented as the mean±SE (n=8). CTL, Control; Res, resveratrol.
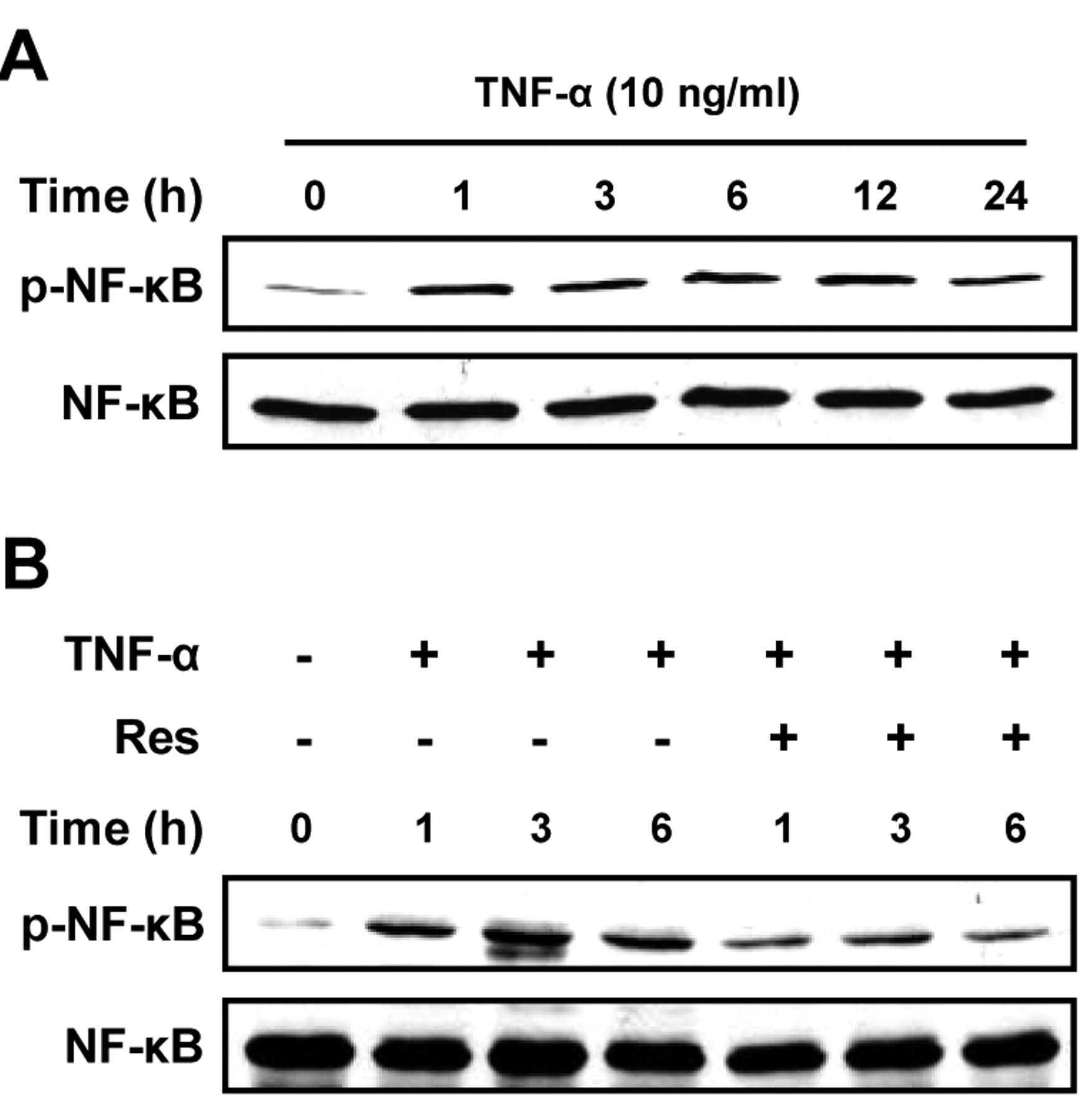
Effect of resveratrol on TNF-α-induced NF-κB activation in U373MG cells. A: Protein lysate was extracted from U373MG cells treated with TNF-α (10 ng/ml) for the indicated times and western blotting was performed to detect phosphorylation of NF-κB. Blots were reprobed for total NF-κB. B: Protein lysate was extracted from U373MG cells treated with TNF-α (10 ng/ml) in the presence or absence of resveratrol (10 μM) for the indicated times and western blotting was performed to detect phosphorylation of NF-κB. Blots were reprobed for total NF-κB.
Discussion
In this study, we showed that resveratrol reduced TNF-α-induced U373MG human glioma cell invasion. Resveratrol repressed NF-κB activation and down-regulated mRNA expression of uPA and its receptor uPAR in TNF-α-treated glioma cells.
It is well documented that TNF-α is a strong stimulatory factor in various cancer cell lines (38). While TNF-α induces cytotoxic effects in many types of cancer, glioblastoma has been shown to be resistant to TNF-α-induced cell death (8). Interestingly, TNF-α elicits tumor cell motility and invasion through the activation of NF-κB in glioblastoma (39). In this study, TNF-α enhanced U373MG cell invasion and induced NF-κB activation, as expected. In previous studies, resveratrol inhibited TNF-α-induced cell invasion in many types of cancer cells through inhibition of NF-κB signaling (32, 40). Resveratrol represses the expression of interleukin-6, B-cell lymphoma 2 (BCL-2), BCL-xL, X-linked inhibitor of apoptosis protein, cellular inhibitor of apoptosis protein, vascular endothelial growth factor (VEGF), and matrix metalloproteinase-9 (MMP-9), the syntheses of which is regulated by NF-κB (36, 41). Furthermore, resveratrol is also known to inhibit the NF-κB pathway (42). In this study, we show that resveratrol suppressed glioma cell invasion and NF-κB activation in TNF-α-treated U373MG human glioma cells.
Upon binding to uPAR, uPA converts inactive plasminogen to active plasmin. Plasmin can degrade extracellular matrix protein components and change other proteins into active forms, including MMP-2, -3, -9, and 13 (43, 44). Both uPA and uPAR have been identified as key factors that regulate cancer cell motility, invasion and metastasis (45-47). Furthermore, uPA and MMP-9 have been proven be expressed at high levels in human malignant brain tumors in vivo (48, 49) and these proteases act as key regulators in glioblastoma invasion and pregression (50, 51). In addition, NF-κB is an important upstream regulator of MMPs and the plasminogen activator system.
As TNF-α-induced U373MG cell invasion is regulated by NF-κB signaling, it was shown herein that TNF-α enhanced uPA and uPAR mRNA expression, supposedly through NF-κB activation, in U373MG cells. Therefore, the inhibition of cancer cell invasion mediated by uPA and uPAR may be a key feature for the prevention of U373MG cell invasion. Similar results have been found for epigallocatechin-3-gallate, the most abundant polyphenol in green tea, which inhibits migration and invasion of OC2 cells by inhibiting the activation of MMP-2, MMP-9, and uPA enzymes (52, 53). Importantly, TNF-α-induced uPA and uPAR mRNA expression was inhibited by resveratrol in U373MG cells. Therefore, we propose that the anti-invasive effect of resveratrol is associated with the inhibition of NF-κB activity. It is known that suppression of NF-κB expression restrains cancer cell invasion by down-regulating the expression of several molecules, including MMP-2, MMP-9, membrane type 1-MMP, uPA and VEGF at the mRNA level (54). In particular, resveratrol was shown to reduce MMP-2 mRNA levels (3). Resveratrol inhibits the NF-κB pathway and consequently should have effects on proteolysis of extracellular matrix protein at multiple levels, e.g. by inhibiting MMP-2 activation and reducing MMP-2 transcripts.
In summary, these results demonstrate that resveratrol inhibits U373MG cell invasion by down-regulating NF-κB activation and uPA/uPAR expression. At the concentrations we tested, we did not find significant cytotoxicity of resveratrol. In a previous report, resveratrol induced apoptotic cell death of human U251 glioma cells at higher concentrations (27). Thus, we do not exclude the possibility that higher concentrations of resveratrol could affect U373MG glioma cell viability. It is worth mentioning that resveratrol functions as an anticancer agent at multiple steps of cancer progression (22). In particular, resveratrol can sensitize cancer cells to other conventional anticancer therapeutics (42). In conclusion, resveratrol as a therapeutic supplement, would be beneficial to patients with invasive glioma, although more research on various effects of resveratrol on glioma is required.

Effect of resveratrol on the TNF-α-induced mRNA expression of uPA and uPAR in U373MG cells. A: U373MG cells were treated with TNF-α (10 ng/ml) for the indicated times. The transcripts of uPA and uPAR were detected by RT-PCR. GAPDH was chosen as a housekeeping gene. B: The band intensities of uPA and uPAR transcripts were compared to those of GAPDH. n=3. C: U373MG cells were treated with TNF-α (10 ng/ml) in the presence or absence of resveratrol (10 μM) for 1 and 3 h. The mRNA level of uPA, uPAR was examined by RT-PCR. GAPDH was used as loading control. D: The band intensities of uPA and uPAR transcripts were compared with those of GAPDH. Data are presented as the mean±SE *p<0.05 (vs. TNF-α). n=3. AU, Arbitrary units; Res, resveratrol.
Acknowledgements
This research was supported by Basic Science Research Program through the National Research Foundation of Korea funded by the Ministry of Education, Science and Technology (20110006200).
- Received March 31, 2011.
- Revision received November 9, 2011.
- Accepted November 10, 2011.
- Copyright© 2011 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved





{kind=link}
{kind=link}
{kind=link}
{kind=link}