


Abstract
Background: Monocytes/macrophages may be affected by tumour cells via cell-to-cell contact, soluble factors and by tumour-derived microvesicles (TMV). Previous observations indicate that TMV interact with monocytes and alter their immunophenotype and activity. This study was designed to determine interactions of TMV with subpopulations (CD14++CD16− and CD14+CD16++) of human monocytes. Methods: Engulfment of TMV by subsets of monocytes was analysed by flow cytometry. Moreover cytokine release and production of reactive oxygen intermediates (ROI) and reactive nitrogen intermediates (RNI) by CD14++CD16− and CD14+CD16++ cells after TMV stimulation was determined. Results: It was found that TMV are engulfed more efficiently by CD14++CD16− than CD14+CD16++ cells. TMV-activated CD14++CD16− cells produce more ROI and interleukin -10 (IL-10) than CD14++CD16+. CD14+CD16++ cells following TMV stimulation showed an increased release of tumour necrosis factor alpha, IL-12p40 and RNI. Conclusion: TMV significantly modulate biological activity of monocyte subsets with a pattern similar to tumour cells. Therefore, TMV mimic the activating effect of tumour cells on monocytes as assessed by release of cytokines, ROI and RNI.
Human monocytes (MO) are heterogeneous population of cells that differ in morphology, phenotype and function (1-5). In particular, the different expression of CD14 (the part of the receptor for lipopolysaccharide) and of CD16 (Fcγ receptor III) was initially used to define two major subsets in peripheral blood, the so called ‘classical’ CD14++ CD16− MO, typically representing up to 90-95% of all MO, and ‘non-classical’ CD14+CD16++ comprising the remaining fraction of MO (6, 7). The CD14+CD16++ subset is the main producer of tumour necrosis factor (TNF) (8) and is expanded during infection and inflammation, in particular in sepsis (9-11). This subset is also formed during in vitro cytokine-induced differentiation from CD14++CD16− (12), supporting the earlier suggestion that it may represent a transitory stage of its differentiation (4). CD14+CD16++ cells constitute the main subpopulation of blood monocytes involved in antitumour response as judged by an enhanced cytotoxicity and production of proinflammatory cytokines (TNF, IL-12) and reactive nitrogen intermediates (RNI) (13). It has been previously suggested that MO and tumour-infiltrating macrophages (TIM) may be affected by tumour cells not only by a direct cell-to-cell contact and soluble factors but also by tumour-derived microvesicles (TMV) (14-16). TMV are small membrane fragments that are released spontaneously by tumour cells during proliferation, migration, activation and apoptosis (16-19). TMV are heterogeneous in shape and size (10 nm - 1 μm) (16, 20). TMV show the expression of several tumour cell determinants such as CD44, CD95L, CD147 (extracellular matrix metalloproteinase inducer, EMMPRIN), CD66 (carcinoembryonal antigen CEA), chemokine receptors (CCR6, CX3CR1) and tumour-associated antigens (Her2/neu) (17, 20-27).
Using an in vitro model, MO interactions with TMV released by three tumour cell lines (pancreatic, lung and colorectal carcinomas) have previously been described (16). TMV, as well as tumour cells, are able to activate MO as judged by production of cytokines (TNF, IL-10, IL-12), ROI (28) and chemokines (unpublished). With this in mind, this study compared interactions of CD14++CD16− and CD14+CD16++ MO subsets with TMV in terms of cytokines, ROI and RNI production during in vitro culture.
Materials and Methods
TMV isolation. TMV were obtained as previously described (16). Briefly, human cell lines: HPC-4 (pancreatic adenocarcinoma, TMVHPC) and DeTa (colorectal adenocarcinoma, TMVDeTa) were cultured in RPMI 1640 medium (Sigma, St. Louis, MO, USA) with 5% foetal bovine serum (FBS; PAA Laboratories, Pasching, Germany). Supernatants from well grown cell cultures were collected and spun down at 2000×g for 20 min to remove cell debris. Then supernatants were again centrifuged (RC28S, Sorvall, Newton, CT, USA) at 50,000×g for 1 hour at 4°C. Pellets were washed several times to remove FBS and finally resuspended in serum-free RPMI-1640 medium. Quantification of TMV proteins was evaluated by the Bradford method (BioRad, Hercules, CA, USA). TMV were tested for endotoxin contamination by the Limmulus test according to manufacturer's instruction (Charles River Laboratories, Inc., Wilmington, MA, USA) and stored at −20°C. TMVHPC were used in all experiments, where TMVDeTa only for RNI testing.
Isolation of MO subpopulation. Human peripheral blood mononuclear cells were isolated from EDTA-blood of healthy donors by the standard Ficoll/Isopaque (Pharmacia, Uppsala, Sweden) density gradient centrifugation. MO were separated from mononuclear cells by counter-flow centrifugal elutriation with the JE-6.B elutriation system equipped with a 5 ml Sanderson separation chamber (Beckman, Palo Alto, CA, USA), as previously described (29). MO were suspended in RPMI-1640 culture medium supplemented with L-glutamine and gentamycin (25 μg/ml) (Sigma). The CD14++CD16− and CD14+CD16++ monocyte subpopulations were isolated from elutriated MO by cell sorting with the use of a FACS Aria cell sorter (BD Biosciences Immunocytometry Systems, San Jose, CA, USA). MO were labelled by using CD14 APC (allophycocyanin) and CD16 PE-Cy7 (phycoerytrin-cyanine 7) conjugated monoclonal antibodies (mAbs; BD Biosciences Pharmingen, San Diego, CA, USA). To exclude natural killer (NK) cells contamination surface expression of HLA-DR antigens was checked for by using peridinin chlorophyll protein complex (PerCP) conjugated mAb (BD Biosciences). Sorted cells (CD14++CD16− and CD14+CD16++) were collected into tubes precoated with FBS to avoid plastic charging and cell attachment to the wall. The purity of sorted cells was checked by flow cytometry and exceeded 95%.
Transfer of TMV to MO subpopulations. TMV were incubated for 5 min with red PKH26 dye (Sigma) according to the manufacturer's instructions. Then TMV were washed with 1% bovine serum albumin and several times in serum-free RPMI-1640 medium. Sorted cells CD14++CD16− and CD14+CD16++ (1×106 cells/ml) were incubated with PKH26-labelled TMV (30 μg/ml) for 18 hours at 37°C in the medium supplemented with 5% FBS. Engulfment of PKH26-labelled TMV by MO subsets was determined by flow cytometry (FACSCanto, BD Bioscience). Red fluorescence intensity and the percentage of positive cells were analysed after quenching extracellular fluorescence with crystal violet as described before (16).
Determination of cytokine secretion. Control MO (elutriated but not sorted) and sorted CD14++CD16− and CD14+CD16++ cells were cultured for 18 hours in 96 well microplates (1×105 cells/well) at 37°C, 5% CO2 in a humidified atmosphere in the presence of TMV (30 μg/ml). Supernatants were collected and the concentrations of TNF, IL-10 and IL-12p40 were measured by ELISA. The following matched mAbs pairs for ELISA were used (PharMingen, San Diego, CA, USA): for TNF Mab1 (capture) and Mab11 (detection), for IL-10, JES3-9d7 (capture) and JES-12G8 (detection) and for IL-12p40 C8.3 (capture) and C8.6 (detection). Recombinant human cytokines (all from PharMingen) were used as standards. Tests were performed according to the manufacturer's protocol and results were determined with ELISA reader (BioTek Instruments, Vinooski, VT, USA) at 492 vs. 630 nm wavelength. Detection level for TNF was 20 pg/ml, and 10 pg/ml for IL-10, IL-12p40. The tested cytokines were not detected in TMV.

Transfer of PKH26-labelled TMVHPC to MO subsets. Flow cytometry of CD14++CD16− (right panel) and CD14+CD16++ (left panel) cells exposed to PKH26-labelled-TMVHPC for 18 hours followed by quenching of extracellular fluorescence with crystal violet. One representative experiment of four performed is presented.

The effect of TMV on cytokine release by MO subpopulations. MO subsets were incubated for 18 hours with TMVHPC (30 μg/ml). Cytokine levels in the culture supernatants were determined by ELISA. Cytokines were not detected in the culture supernatant of control monocytes (medium) or TMV alone. Data represent the mean±SD of six independent experiments. *p<0.05.
Determination of ROI production by flow cytometry. The intracellular production of O2- and H2O2 was measured by flow cytometry using oxidation-sensitive fluorescent probes hydroethidine (HE, Sigma) and dihydrorhodamine 123 (DHR123), respectively (30). MO and sorted subsets of MO were incubated at 37°C for 2 hours alone or with TMV (30 μg/ml) in the presence of 10 μM HE and 40 μM DHR123 (both from Sigma). Analysis was performed with FACSCanto cytometer (BD Biosciences).
Nitric oxide secretion. MO and their subpopulations (2×105 cells/well) were cultured with TMVDeTa (30 μg/ml) for 18 hours in the presence of fluorescent dye diaminofluorescein-2 (DAF-2). TMVDeTa were chosen because only DeTa tumor cells induce NO in our hands (31). TMVHPC were used as a negative control. NO production was analysed by flow cytometry.
Statistical analysis. Statistical analysis was performed by nonparametric Mann-Whitney test. Differences were considered significant at p<0,05.
Results
Determination of TMV engulfment. It has been previously shown that TMV can adhere to MO surface and this adherence is followed by the engulfment of TMV. The present study investigated whether engulfment of TMV was similar for both subsets (CD14++CD16− and CD14+CD16++) of MO. The MO subsets were incubated with PKH26-labelled TMV for 18 hours. Data showed that after an overnight incubation TMV were located intracellulary, mainly in CD14++CD16− cells. Only a small percentage of CD14+CD16++ cells showed red fluorescence inside (Figure 1). No differences in TMV attachment to monocyte subsets were observed (data not shown).
Cytokine secretion. Previous studies have shown that TMV, as well as tumour cells, stimulate production of TNF, IL-10, IL-12 by MO (28, 32). The experiments in this study were undertaken to determine cytokine secretion by the subpopulations of MO stimulated with TMV. Figure 2 shows that the whole population of MO and CD14++CD16− cells after stimulation with TMV secreted comparable amounts of TNF, IL-10 and IL-12p40. Secretion of TNF and IL-12p40 by CD14+CD16++ cells was significantly higher in comparison to initial MO or CD14++CD16− cells. In contrast, secretion of IL-10 by CD14+CD16++ cells was significantly lower than by MO and CD14++CD16−. Neither TMV alone or control monocytes released measurable amounts of tested cytokines (data not shown).
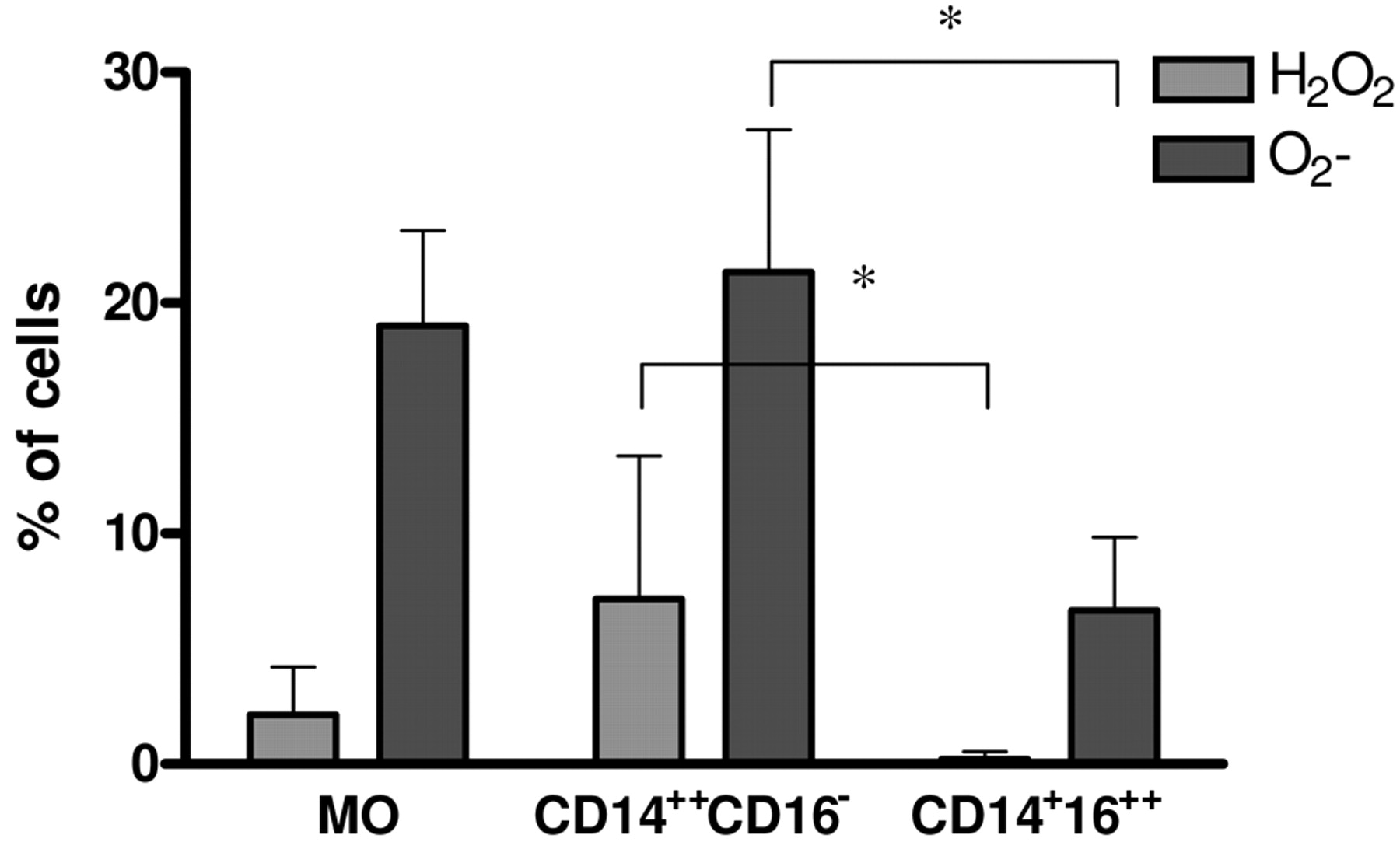
Production of ROI. Tumour cells and TMV are able to induce ROI production by MO (28, 32). As tumour cells induce ROI production mainly by CD14++CD16− cells (13), this study examined whether this subpopulation is the main producer of ROI after TMV stimulation. Initial MO and their subsets were stimulated for 2 hours with TMV and production of H2O2 and O2− was measured by flow cytometry. Figure 3 shows that ROI production by MO and CD14++CD16− cells was comparable and significantly higher than by CD14+CD16++ cells.
Production of RNI. Production of RNI (NO) was assessed by intracellular staining with DAF-2 in MO and their subsets after 18 h stimulation with TMVDeTa. A significantly higher percentage of DAF-2 positive cells was found among CD14+CD16++ cells in comparison to CD14++CD16− cells (Figure 4), suggesting that the former are main producers of NO.
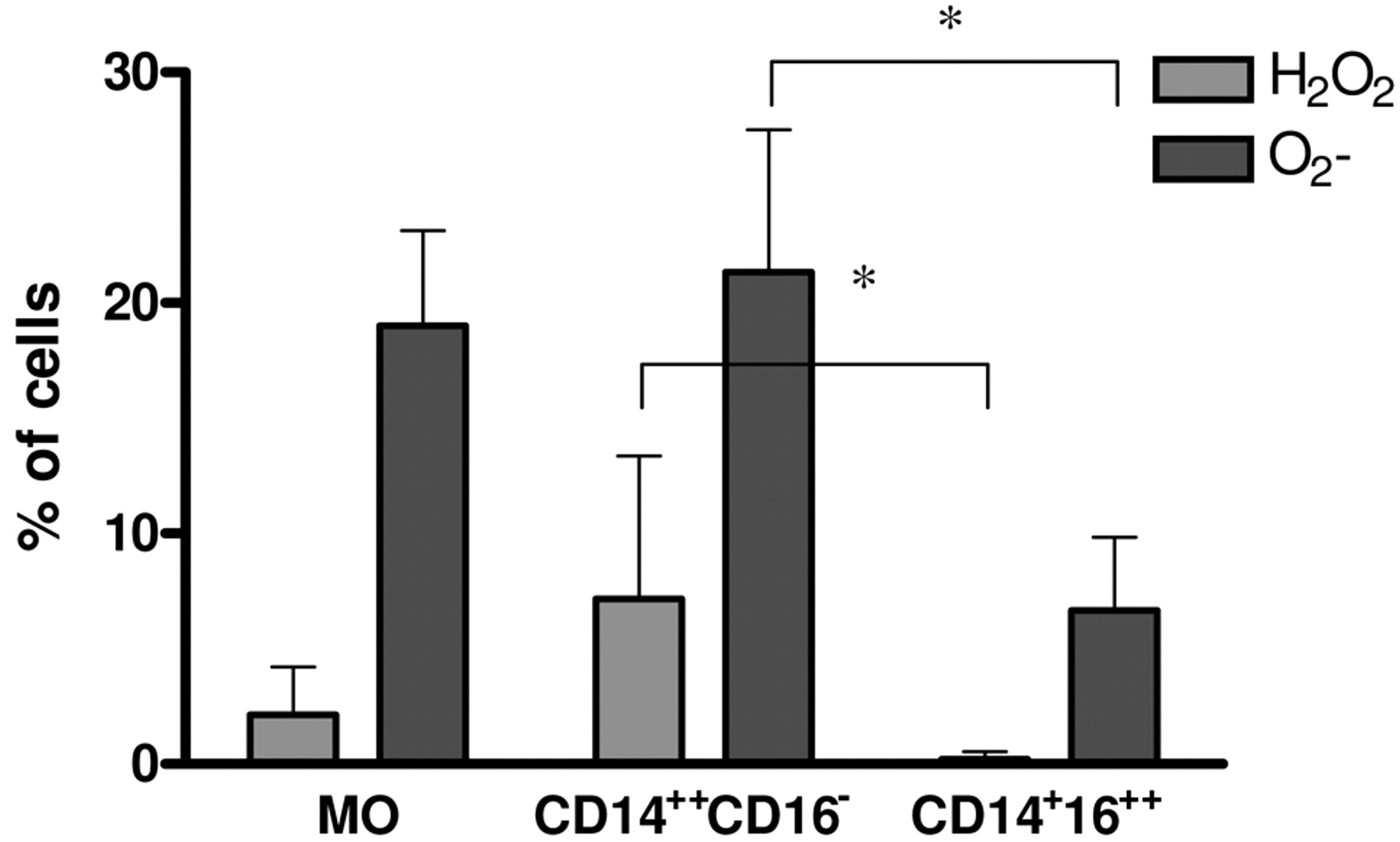
The effect of TMV on intracellular production of O2- and H2O2 by MO subpopulations. The level of O2- and H2O2 production was determined by flow cytometry using oxidation-sensitive probes HE and DHR123, respectively. MO subsets were incubated alone or with TMV (30 μg/ml) in the presence of HE or DHR123 for 2 hours. Data represent mean±SD of five independent experiments; *p<0.05.

The effect of TMV on production of NO by MO subpopulations. The level of NO was determined by flow cytometry using DAF-2 probe. MO subsets were incubated alone or with TMV (30 μg/ml) in the presence of DAF-2 for 18 h. Data represent mean±SD of six independent experiments; *p<0.05.
Discussion
The present study showed that subpopulations (CD14++CD16−, CD14+CD16++) of MO interact with TMV in different manners. First, there were differences in TMV engulfment by MO subsets, as CD14++CD16− internalized more TMV than CD14+CD16++ cells. Second, CD14+CD16++ cells were the main subpopulation of MO, which, when stimulated with TMV, produce pro-inflammatory cytokines (TNF, IL-12) and RNI. In contrast, CD14++CD16− cells exposed to TMV released more IL-10 and ROI, but less TNF and IL-12p40.
MO and macrophages play a dual role in the host's response to the growing tumour as they both enhance or inhibit proliferation of malignant cells (14). The final outcome of MO effect may depend on the production of various mediators including cytokines, growth factors, proangiogenic and angiostatic factors, etc. On the other hand, tumour cells may affect MO by direct cell-to-cell contact, via soluble factors or by TMV (14, 28, 33, 34). Contact of MO with TMV results in cytokine (TNF, IL-10, IL-12) and ROI production with the pattern similar to tumour cell stimulation (28, 32, 35).
Since the response of MO subsets to tumour cells is different, it was investigated whether MO subsets interact with TMV in a pattern similar to tumour cells. CD14++CD16− cells are characterised by a high phagocytic activity and generation of ROI (13, 36). This is in keeping with the current observations that after TMV stimulation CD14++CD16− cells release less TNF and IL-12 and show enhanced ROI production (H2O2 and O2) in comparison to the CD14+CD16++ subset. These data were supported by analysis of TMV engulfment by subpopulation of MO. Approximately 60% of CD14++CD16− showed PKH26-labelled TMV red fluorescence not quenched by crystal violet, which indicates intracellular localization of TMV. Strong engulfment of TMV by CD14++CD16− cells, known as precursors of professional phagocytes, may suggest the role of phagocytosis/receptor mechanism in TMV internalization. On the other hand, CD14+CD16++, which are rather poor phagocytes, expressed only 10% of cells with intracellular red fluorescence after incubation with PKH26 labelled-TMV. CD14+CD16++ cells represent only 5-10% of total human MO (37), and showed an increased release of TNF, IL-12 and RNI after lipopolysaccharide or tumour cell stimulation, in comparison to CD14++CD16− cells (4, 7, 13, 36). Also stimulation with TMV induces significantly higher secretion of proinflammatory cytokines (TNF, IL-12p40) by CD14+CD16++ than CD14++CD16− cells. This is in concordance with previous data suggesting that CD14+CD16++ cells are the main subpopulation of MO involved in antitumour response as judged by cytotoxicity and production of proinflamatory cytokines (13). In addition, it has previously been shown that HLA-DR is involved in signalling for TNF production by monocytes stimulated with tumour cells (38). This may be relevant also in the present study, as an increased expression of HLA-DR is observed in the CD14+CD16++ subpopulation. TMV, which are present in culture supernatant may possibly induce NO production in monocytes via CD29 or CD44 molecules, which are expressed on MO. The current results clearly showed that TMV may mimic tumour cell effect on MO subsets. The precise mechanism of interactions of TMV with MO subset is not clear, but ligand-receptor binding should be taken into consideration as TMV may carry several antigens presented on tumour cells (16, 20). This hypothesis was partially proven, as blocking CD44 molecules on MO was sufficient to diminish production of TNF after TMV stimulation (28).
In summary, the exact activity of human MO in malignancy remains not fully understood, but the data presented here increase the knowledge about MO–tumour cell interactions, highlighting the role of TMV in monocyte-mediated antitumour response.
Acknowledgements
This study was supported by the Jagiellonian University Medical College (grant no. K/ZDS/001481). The authors thank Ms B. Hajto and I. Ruggiero for skilful technical assistance.
- Received May 31, 2010.
- Revision received June 21, 2010.
- Accepted June 25, 2010.
- Copyright© 2010 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved





{kind=link}
{kind=link}
{kind=link}
{kind=link}