


Abstract
Background/Aim: The study is directed to the effect of resveratrol on the redox-status and viability of leukemic and normal lymphocytes, as well as its ability to sensitize leukemic lymphocytes to anticancer drugs. Materials and Methods: Cytotoxicity was analyzed by trypan blue staining, apoptosis – by Annexin V test, and oxidative stress – by the intracellular levels of reactive oxygen species (ROS) and protein-carbonyl products. Results: Incubation of resveratrol in combination with the majority of anticancer drugs resulted in higher toxicity than resveratrol or drug alone. In the case of leukemic lymphocytes treated with barasertib and everolimus in the presence of resveratrol, synergistic cytotoxicity was accompanied by strong induction of apoptosis, increased levels of hydroperoxides and insignificant changes in protein-carbonyl products. None of these parameters changed in normal lymphocytes. Conclusion: Resveratrol is a promising supplementary compound for anticancer therapy, that may allow reduction of the therapeutic doses of barasertib and everolimus, minimizing their side-effects.
It has been demonstrated that the effect of conventional anticancer drugs is usually accompanied by the induction of oxidative stress in cancer cells (1-6). However, conventional chemotherapy has harmful side-effects due to increased intracellular ROS production and disruption of redox-homeostasis in normal cells and tissues (5-7). Combined treatment of cancer with chemotherapeutics and natural or synthetic redox-modulators has been proposed to restrain their harmful side-effects (4, 6, 7). Applied alone, some of the redox-modulators show pro-oxidant effects, depending on the environmental conditions, however, the increased ROS production is due to activation of regulatory pathways (cell proliferation, apoptosis, activation of adaptive mechanisms through stimulation of cellular antioxidant systems), rather than direct induction of free radical damages and cell death (8, 9).
In our previous studies, we found that some redox-modulators [e.g., melatonin, 2-deoxy-D-glucose, 6-aminonicotinamide, docosahexaenoic acid, and Apatone® (vitamins C/menadione)] sensitize leukemic lymphocytes to anticancer drugs (10-15). Some combinations (redox-modulator plus anticancer drug) display synergistic cytotoxicity towards leukemic cells, without elevating ROS production. In addition, these redox-modulators did not significantly affect cell viability or the levels of ROS and did not induce apoptosis in normal lymphocytes, when combined with new-generation anticancer drugs (e.g., everolimus and barasertib) (10-15).
Over the last decade, resveratrol has been recognized as an attractive redox-modulator, that can be used as an adjuvant to conventional chemotherapy, reducing its side-effects (6, 16). Resveratrol is a bioactive compound, which is produced by plants as a phytoalexin in response to a stressful stimulus, or to a microbial or fungal infection, providing protection to plants (16-18). Due to its polyphenolic structure (two phenol rings linked to each other by an ethylene bridge and the presence of three phenolic hydroxyl groups), resveratrol exhibits antioxidant properties. They depend on the redox-activity of its phenolic hydroxyl groups and the potential for electron delocalization across the chemical structure (16). Many studies have confirmed that resveratrol has also anti-inflammatory and anticancer activities (16, 19-21). In vitro and in vivo studies have shown that resveratrol (applied alone) suppresses cancer progression in a wide variety of tumor cells, including breast, skin, colorectal and prostate cancers (22-25). Growing evidence have suggested that the anticancer activities of resveratrol are mediated by induction of apoptosis and cell differentiation, cell-cycle arrest, suppression of proliferation, reduction of inflammation and angiogenesis, and inhibition of adhesion, invasion and metastasis (19, 26, 27). Although, the anticancer activity of resveratrol is well established, the exact mechanism by which it interferes with cancer is not well defined. Its therapeutic effects may be due to ROS scavenging activity and modulation of different intracellular signaling pathways. For example, resveratrol could provoke cell death by up-regulating of cyclin-dependent kinase inhibitors (p21), tumor suppressor gene products (e.g., p53) or by inhibiting of cell survival kinases [e.g. mitogen-activated protein kinase (MAPKs), protein kinase B (AKT), phosphoinositide 3-kinase (PI3K), epidermal growth factor receptor (EGFR) kinase] and survival transcription factors [nuclear factor-kappa B (NF-kB), activating protein 1 (AP-1)] (16, 19-22, 26, 28). Resveratrol is used to lower calories, improve health and prevent aging, which is related to its ability to activate sirtuins (6, 29-31).
Resveratrol suppresses the oxidative stress in isolated cells and in vivo due to its ROS scavenging activity and up-regulation of some antioxidant enzymes (15). Three different mechanisms of the antioxidant effect of resveratrol are described: (i) competition with coenzyme Q and decrease of ROS production by the mitochondrial electron-transport chain; (ii) scavenging of superoxide radicals formed in mitochondria; and (iii) inhibition of the lipid peroxidation induced by Fenton's reactions (24, 32). On the other hand, it has been reported that resveratrol (in high doses) reduces cell growth and induces apoptosis in normal cells, which confirms its biphasic effects (18, 33).
The present study aimed to investigate the effect of resveratrol on the redox-status and viability of leukemic and normal lymphocytes, as well as its ability to sensitize leukemic lymphocytes to a variety of anticancer drugs (conventional and new-generation).
Materials and Methods
Cells and treatment protocol. The experiments were performed on leukemic lymphocytes (Jurkat), derived from patients with acute lymphoblastic leukemia, as well as on normal lymphocytes, isolated from healthy blood donors using Lymphosepar-I (Immune-Biological Laboratories Co., Fujioka, Japan). The cells were cultured in RPMI-1640 medium (Sigma-Aldrich, Steinem, Germany), supplemented with 10% heat-inactivated fetal bovine serum (Gibson, Nashville, TN, USA) and antibiotics (100 U/ml penicillin and 100 μg/ml streptomycin) (Gibson) in a humidified atmosphere at 37°C with 5% CO2. Before treatment with anticancer drugs, all cells were collected by centrifugation (1,000 × g, 10 min) and refed with fresh medium without antibiotics.
The drugs were dissolved in dimethyl sulfoxide (DMSO; Sigma-Aldrich) or phosphate-buffered saline (PBS; 10 mM, pH 7.4). The final concentration of DMSO in the cell suspension did not exceed 1%. At this concentration, DMSO did not influence cell viability.
The cells (1×106 cells/ml) were incubated with resveratrol, drug, or drug plus resveratrol at the following concentrations: 12.5 μM resveratrol (Sigma-Aldrich), 0.25 μM palbociclib (Selleckchem, Houston, TX, USA), 0.5 μM PI-103, 5 μM (Selleckchem) everolimus, 0.5 μM lonafarnib (Selleckchem), 0.1 μM ABT-737 (Selleckchem), 0.1 μM doxorubicin (Sigma-Aldrich), 0.5 μM bleomycin (Nippon Kayaku Co., Tokyo, Japan), 0.1 μM AZD-7762 (Sigma-Aldrich), 0.01 μM MLN-2238 (Selleckchem), 0.025 μM MG-132 (Wako, Tokyo, Japan), 10 μM lomustine (Sigma-Aldrich), 2.5 μM cisplatin (Selleckchem), 0.025 μM BEZ-235 (Selleckchem), 0.01 μM bortezomib (Selleckchem), 0.05 μM or 0.01 μM barasertib (Selleckchem).
Cell viability assay. Cell viability was analyzed using trypan blue staining and Countess™ Automated Cell Counter (Invitrogen, OR, USA). Three independent experiments (with two repetitive measurements) were performed for each sample. Non-treated cells were used as controls. The data are presented as the mean±SD.
Intracellular ROS assay. The amount of ROS was analyzed using OxiSelectTM In vitro ROS/RNS Assay Kit – Green Flourescence (Cell Biolabs, Inc., San Diego, CA, USA). The method is based on the use of fluorogenic probe 2’,7’-dichlorodihydrofluorescin DiOxyQ (DCFH-DiOxyQ). In the cytosol, the probe is deacetylated to the non-fluorescent 2’,7’-dichlorodihydrofluorescin (DCFH). DCFH reacts with ROS and reactive nitrogen species (RNS) (predominantly H2O2, ROO., NO, ONOO−) with formation of fluorescent product 2’,7’-dichlorodihydrofluorescein (DCF). The intensity of DCF fluorescence (λex=480 nm, λem=530 nm) is proportional to the amount of ROS/RNS in the biological sample.
The amount of ROS/RNS was calculated by a calibration curve based on DCF standard solution in PBS. The measurements were performed on a Tecan Infinite F200 PRO (Tecan Austria GmbH, Mannedorf, Austria) microplate reader.
Briefly, cells (1×106 cells/ml) were collected by centrifugation (1,000 × g for 10 min) and lysed by using 300 μl of 0.1% sodium dodecylsulfate (SDS; dissolved in PBS) followed by a 30 min incubation on ice. The lysates were adjusted to equal protein concentration (range=1-10 mg/ml) using PBS. Protein concentration was analyzed by Bradford assay. Each sample was subjected to ROS/RNS assay, according to the manufacturer's instructions.
Protein–carbonyl assay. The amount of protein–carbonyl products was analyzed using OxiSelect™ Protein Carbonyl Spectrophotometric Assay Kit (Cell Biolabs, Inc.). The most common products of protein oxidation in biological samples are protein–carbonyl derivatives of proline, arginine, lysine and threonine, which are chemically stable and serve as markers of oxidative stress. The analysis of these products is based on derivatization of the carbonyl groups with dinitrophenylhidrazine and the formation of protein–hydrazone. The amount of protein–hydrazone was detected spectrophotometrically at 375 nm.
Briefly, cells (1×106 cells/ml) were collected by centrifugation (1000 × g, 10 min) and lysed using 300 μl of 0.1% SDS (dissolved in PBS) followed by a 30 min incubation on ice. The lysates were adjusted to equal protein concentration (range=1-10 mg/ml) using PBS. Protein concentration was analyzed calculated by the Bradford assay. Each sample was subjected to protein–carbonyl assay, according to the manufacturer's instructions. Oxidized bovine serum albumin was used as a standard.
Apoptosis assay. The induction of apoptosis was analyzed by the expression of phosphatidylserine (PSer) on the cell surface, using FITC-Annexin V Apoptosis Detection Kit (BioVision, Milpitas, CA, USA). Briefly, the cells (1×106 cells/ml) were incubated with drug, resveratrol, or their combination, under the conditions mentioned above. At different time-points, cells were collected by centrifugation (1000×g, 10 min), washed twice with PBS containing 2.5 mM CaCl2 (annexin V-binding buffer) and re-suspended in the same buffer. One hundred microliters of the suspension were incubated with 5 μl of fluorescein isothiocyanate (FITC)-annexin V for 10 min at room temperature in the dark. The cells were washed three times with annexin V-binding buffer and were finally re-suspended in 500 μl of the same buffer. FITC-annexin V bound to PSer exposed on the cell surface was detected spectrofluorimetrically at λex=488 nm and λem=535 nm, using a Tecan Infinite F200 PRO (Tecan Austria GmbH) microplate reader.
Results and Discussion
Effect of resveratrol on the viability of leukemic and normal lymphocytes. The cells were incubated with different concentrations of resveratrol in order to select the optimal concentration of the redox-modulator, which induces ~20-30% cytotoxicity in leukemic lymphocytes after 48-h incubation. This level of cytotoxicity enables assessment of synergism, additivity or antagonism after combined application of resveratrol with anticancer drugs.
The data on Figure 1 demonstrate that administration of 12.5 μM resveratrol decreased the viability of leukemic lymphocytes ~15-20% (Figure 1A), but did not influence the viability of normal lymphocytes (Figure 1B). Even at 50 μM, resveratrol did not induce cytotoxicity in normal lymphocytes.
This data are consistent with many studies, demonstrating the anticancer effects of resveratrol on cultured cells (15, 19-28). Gautam et al. have reported that resveratrol in concentrations 10-80 μM inhibits cell proliferation of human leukemic cell lines U937 and HL-60 in a dose-dependent manner (34). Jiao et al. have also shown that resveratrol in concentrations 25-250 μM suppresses cell growth and induces apoptosis in leukemic lymphocytes derived from patients with acute lymphoblastic leukemia (CEM-C-1-15, Jurkat, Molt-4 and CEM-C7-14) (35). The authors have observed that after 48-h treatment with 200 μM resveratrol, the inhibition rate of CEM-C-1-15, Jurkat, Molt-4 and CEM-C7-14 cells was ~52%, ~69%, ~58%, and ~42%, respectively. A similar anti-proliferative effect of resveratrol has been described by Ferry-Dumazet et al. (33). The authors have found that resveratrol inhibits proliferation and induces apoptosis in all tested lymphoid and myeloid leukemic cells, with IC50 values between 5 and 43 μM. However, in contrast to our data, they have observed that resveratrol induces apoptosis in normal lymphocytes at concentrations (IC50 <8 μM) lower than those required for most leukemic cells. Resveratrol also induces apoptosis in CD34(+) cells and decreases the number of colonies generated by these precursor cells in a dose-dependent manner (IC50 ~60 μM). Billard et al. have detected a slight (negligible) cytotoxicity of four resveratrol derivatives (trans-resveratrol, its dimmer ε-viviferin, and two vineatrols) on the viability of normal peripheral blood mononuclear cells (PBMC) (36). One possible reason of the observed differences in the effect of resveratrol on normal lymphocytes is the presence of free iron in cell suspensions as a result of the isolation process. Even traces of this transition metal ions can cause Fenton's reactions and compromise the results.

Effect of resveratrol on viability of leukemic lymphocytes Jurkat (A) and normal lymphocytes (B) at different concentrations. The data are the mean±SD of six independent experiments. The red arrow indicates the concentration of resveratrol (12.5 μM), selected for further experiments in combination with anticancer drugs.


Effect of resveratrol and anticancer drugs alone or in combination on proliferation of Jurkat cells after incubation for: (A, B) 24-h; (C, D) 48-h; (E, F) 72-h. Incubation conditions: 1x106 cells/ml, resveratrol (12.5 μM) and/or drug (at concentrations given in the Materials and Methods), at 37°C in a humidified atmosphere. The data are the mean±SD of six independent experiments. B: Effect of each combination (drug plus resveratrol) on cell proliferation as a percentage of the effect of drug applied alone. The red line indicates the effect of resveratrol on the proliferation of cancer cells as a percentage of the control (untreated cells). *In this experiment, the concentration of barasertib was 50 nM.
Effect of resveratrol on the cytotoxicity of anticancer drugs towards leukemic lymphocytes. Next, the study aimed to clarify the effect of resveratrol (in concentration 12.5 μM) on the cytotoxicity of anticancer drugs towards leukemic cells Jurkat.
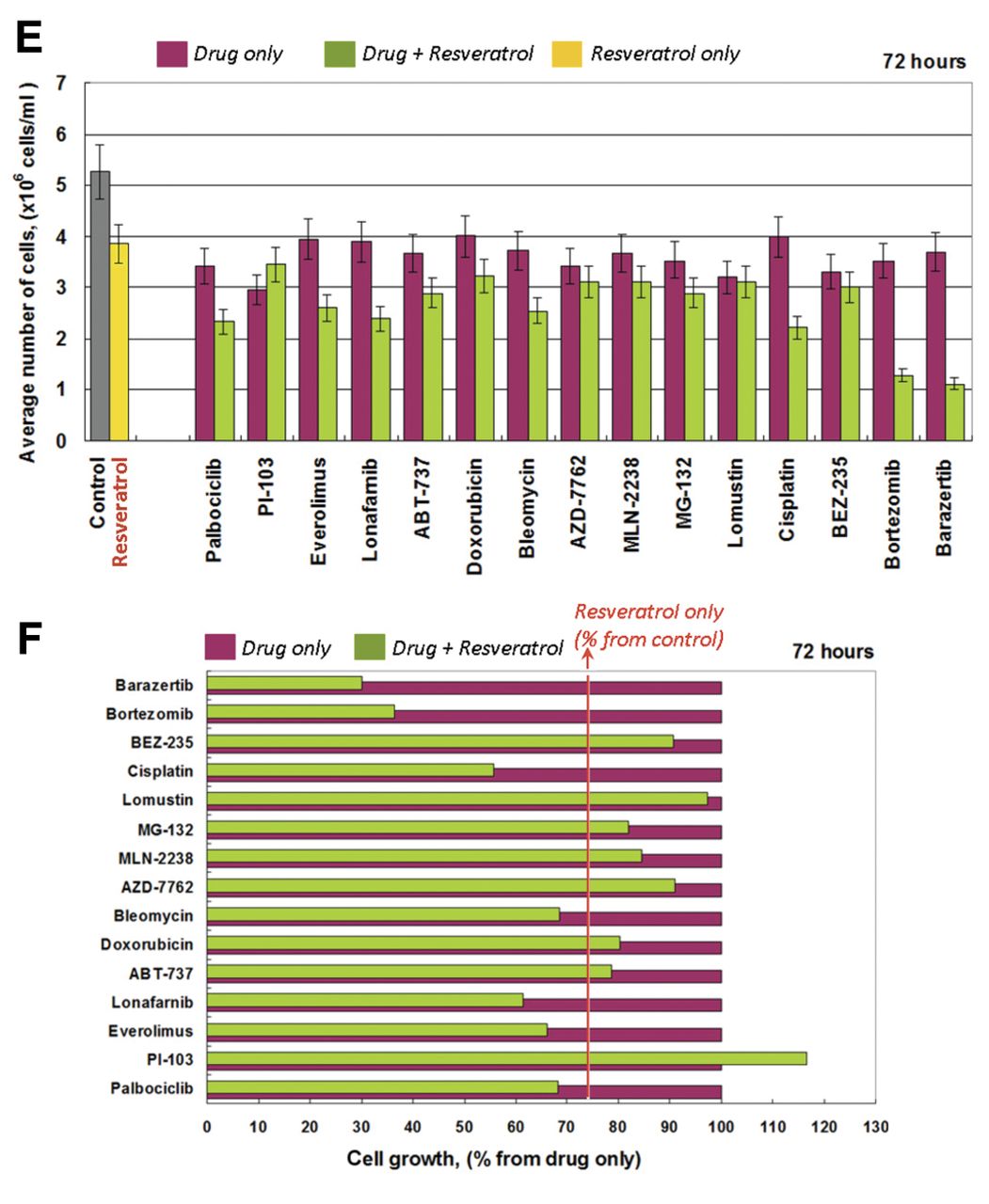
Cells were treated with resveratrol and anticancer drugs, separately and in combination, for different time intervals (24-, 48- and 72-h) (Figure 2). The assessment of cytotoxicity has been described in detail in our previous article (11). The effect of each combination on cell proliferation was calculated as a percentage of the effect of the respective drug applied alone. Then, this effect was compared to the effect of resveratrol, applied alone. The effect of resveratrol is indicated by the red line in Figure 2B. In the case of drug plus resveratrol, the data located to the left of the red line represent synergistic cytotoxic effects, while the data located to the right of the red line represent antagonistic effects. All data matching the red line reflect additive effects. The cytotoxicity of each drug (applied separately at the select concentration) ranged from ~10-20% after 24-h incubation and to ~20-30% after 48- and 72-h incubation.
After 24-h incubation, the majority of combinations (anticancer drug plus resveratrol) displayed additive cytotoxic effects towards Jurkat cells. Three combinations made an exception: resveratrol plus barasertib, bortezomib, or BEZ-235 (Figure 2B). Barasertib and bortezomib showed an increased synergistic effect. The synergistic effect of BEZ-235 disappeared with an increase of incubation time (Figure 2D). After 48- and 72-h incubation, the best synergistic effects were observed with the combination of resveratrol with barasertib, bortezomib, cisplatin, lonafarnib, or everolimus (Figure 2D/F).

Effects of resveratrol (12.5 μM), doxorubicin (0.1 μM) and their combination on cell viability, induction of apoptosis, levels of reactive oxygen species (ROS) and levels of protein-carbonyl products in Jurkat leukemia cells (A) and normal lymphocytes (B), after 24- and 48-h incubation at 37°C in a humidified atmosphere. The data are the mean±SD from three independent experiments.
Effect of resveratrol on doxorubicin-induced oxidative stress and apoptosis in leukemic and normal lymphocytes. The third stage of our study aimed to investigate whether the cytotoxicity of resveratrol in combination with anticancer drugs towards leukemic lymphocytes was accompanied by induction of oxidative stress. Three anticancer drugs were selected: doxorubicin (conventional), everolimus and barasertib (new generation).
It is generally accepted that the cytotoxic effect of doxorubicin is accompanied by overproduction of ROS in cancer cells and this determines its therapeutic effect (4, 6, 12). However, doxorubicin induces also oxidative stress in normal cells, and this is the reason for its harmful side-effects.
Doxorubicin, applied alone on leukemic cells for 48-h, induced apoptosis, accompanied by increased production of ROS, but insignificant changes in the protein-carbonyl level (Figure 3A). Doxorubicin also induced cytotoxicity and apoptosis in normal lymphocytes following 48-h incubation (Figure 3B). In this case, doxorubicin induced a significant increase in protein carbonyl levels without affecting significantly ROS production. These data confirm that the anticancer effect of doxorubicin is accompanied by induction of oxidative stress in both types of cells, but probably through different mechanisms. Resveratrol did not affect doxorubicin-induced ROS production in leukemic lymphocytes, but it decreased the expression of phosphotidylserin on the cell surface of leukemic lymphocytes, as well as protein-carbonyl levels in normal lymphocytes. Resveratrol also abolished the cytotoxicity of doxorubicin on leukemic and normal cells.
It has been shown that resveratrol plays an important protective role on doxorubicin-inducing cardiotoxicity, which is linked to increased ROS production in cardiac cells (6). The authors suggested that the cardioprotective role of resveratrol is due to its effect on mitochondria and the increase in MnSOD activity, as well as in the activation of sirtuin (SIRT1)-dependent pathway (6). Activation of SIRT1 and cardioprotective role of resveratrol was has also been reported by Lou et al. (29). The authors have found that resveratrol (25 μM) significantly reduces cytotoxicity of doxorubicin (5 μM) on H9c2 cells. They have explained the protective effect of resveratrol on doxorubicin-induced cardiotoxicity by the activation of SIRT1 pathway and endoplasmic reticulum stress stabilization (29). Xu et al. have shown that autophagy suppression mediates the ability of resveratrol to protect against doxorubicin-induced cardiotoxicity (37). Rezk et al. have demonstrated that resveratrol plus doxorubicin exhibit additive inhibitory effect on the growth of human ovarian (OVCAR-3) and uterine (Ishikawa) cancer cells (38).

Effects of resveratrol (12.5 μM), everolimus (5 μM) and their combination on cell viability, induction of apoptosis, levels of reactive oxygen species (ROS) and levels of protein-carbonyl products in Jurkat leukemia cells (A) and normal lymphocytes (B), after 24- and 48-h incubation at 37°C in a humidified atmosphere. The data are the mean±SD from three independent experiments.
A number of studies have reported a synergistic cytotoxicity after treatment of leukemic cells with doxorubicin plus resveratrol (39-41), at concentrations much higher than those used in the current study. Resveratrol has been shown to induce cell growth arrest and apoptotic death trough down-regulation of the multidrug resistance-associated protein-1 (MRP1) expression on doxorubicin-resistant leukemic cells, derived from patients with acute myeloid leukemia (39). The authors suggested that resveratrol may facilitate cellular uptake of doxorubicin. Gatouillat et al. have also demonstrated that resveratrol (25 μM) suppresses the growth of doxorubicin-resistant B16 melanoma cells and potentiates the cytotoxicity of doxorubicin (40). Rai et al. have reported synergistic cytotoxicity, inhibition of inflammatory response [NF-kB; cyclooxygenase-2 (COX-2)], redox regulation [nuclear respiratory factor-2 (Nrf2)] and induction of apoptosis (Bax/Bcl-2 ratio and Caspase-9) in breast cancer cell lines (MCF-7, MDA-MB-231), after treatment with resveratrol (IC30) plus doxorubicin (IC20) (41).
Effect of resveratrol on everolimus- and barasertib-induced oxidative stress and apoptosis in leukemic and normal lymphocytes. Everolimus, which is a derivative of rapamycin and also an inhibitor of the mammalian target of rapamycin (mTOR) pathway, plays a fundamental role in the regulation of cell viability, translational initiation and cell cycle progression. This drug is usually used in the clinic as an immunosuppressant to prevent rejection of organ transplants. In the past 10 years, it was found that everolimus also possesses anticancer activity (42-45). Barasertib belongs to a new class of pyrazoloquinazolines, a selective inhibitor of aurora B kinase, discovered and described in 2007 (10, 46). Little is known on the molecular mechanisms of action of barasertib, except that it provokes cell-cycle arrest and apoptosis, and increases the response to chemotherapy (10, 47, 48). In our previous studies, we established that everolimus and barasertib in combination with redox-modulators induced apoptosis of leukemic lymphocytes via ROS-independent (in case of 2-deoxy-D-glucose, 6-aminonicotinamide, melatonin, and Apatone®) or ROS-dependent mechanisms (in case of docosahexaenoic acid) (11-15).
After treatment of leukemic cells with everolimus plus resveratrol for 48 h, we observed strong cytotoxicity and induction of apoptosis, accompanied by an increased production of ROS, but insignificant changes in the protein-carbonyl level (Figure 4A). This combination did not alter the viability of normal lymphocytes, and did not induce oxidative stress in these cells (Figure 4B).
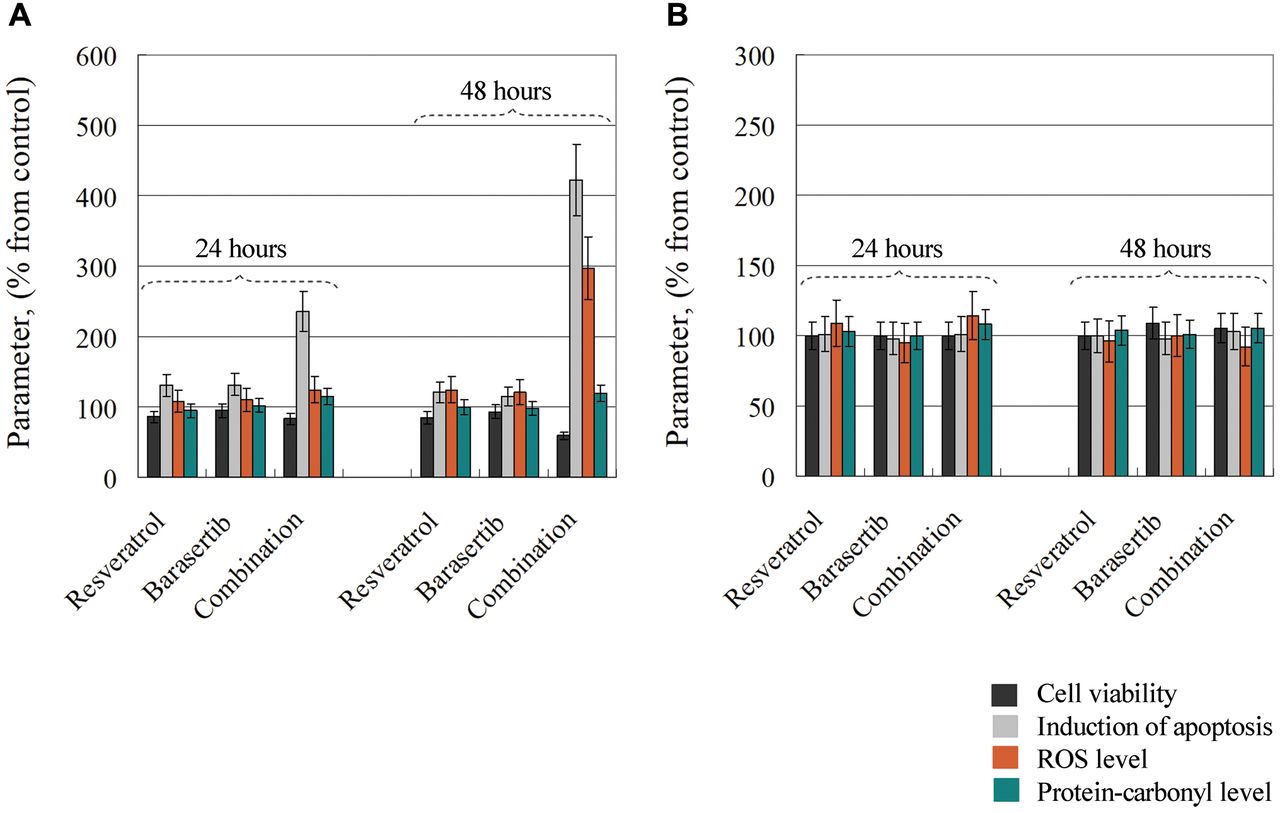
Effects of resveratrol (12.5 μM), barasertib (0.01 μM) and their combination on cell viability, induction of apoptosis, levels of reactive oxygen species (ROS) and levels of protein-carbonyl products in Jurkat leukemia cells (A) and normal lymphocytes (B), after 24- and 48-h incubation at 37°C in a humidified atmosphere. The data are the mean±SD from three independent experiments.
The synergistic effect could be explained (at least partially) by the effect of resveratrol on the mTOR pathway and multiplication (amplification) of the anticancer effect of everolimus. Alayev et al. have reported that in combination with rapamycin, resveratrol inhibits the mTOR pathway and blocks rapamycin-induced Akt activation in human bladder and breast cancer cell lines (49, 50). Recently, Roger-Broadway et al. demonstrated that resveratrol suppresses the endometrosis in ovarian cancer cells through inhibition of the mTOR pathway (51). The same authors have also investigated the effect of everolimus on endometrosis, but not in combination with resveratrol.
There is still no evidence about the effect of resveratrol on barasertib-induced cytotoxicity in cancer cells, as well as the effect of the combination on the viability of normal cells. In our study, both cell lines were treated with a very low concentration of barasertib (10 nM). At this concentration, barasertib applied alone had no affect on the viability of either cell type (Figure 5). However, after treatment of leukemic lymphocytes with barasertib plus resveratrol for 48 h, we observed strong cytotoxicity and induction of apoptosis, accompanied by an increased production of ROS, but insignificant changes in the protein-carbonyl levels (Figure 5A). This combination did not alter the viability of normal lymphocytes, and did not induce oxidative stress in these cells (Figure 5B).
It has been demonstrated that aurora B kinase, which exists in a complex with survivin and mTOR, synergistically regulates survival and proliferation of leukemia and lymphoma cells via cross-talk with AKT, mTOR and Notch signaling pathways (52). Moreover, several studies have reported that aurora kinase inhibitors induce cell cycle arrest and apoptosis in cancer cells through p38 MAPK and AKT/mTOR signaling (53). Therefore, the synergistic cytotoxicity of the combination of barasertib and resveratrol towards leukemic lymphocytes could be explained by multiplication (amplification) of the effects of the two substances on multiple molecular targets including aurora B kinase, AKT/mTOR, and MAPK.
In the literature, there is an abundance of data regarding sensitization of cancer cells to conventional anticancer drugs (such as doxorubicin, bleomycin, cisplatin) by resveratrol (6, 7, 21, 22, 25, 28). A limited number of studies describe the effect of resveratrol on the cytotoxicity of new-generation anticancer drugs such as ABT-737 (a selective inhibitor of B cell lymphoma 2 proteins), MG132, and bortezomib (proteasome inhibitors) (54, 55). There are no data on the effect of resveratrol on the efficiency of AZD7762 (a selective inhibitor of checkpoint kinases), barasertib (a selective inhibitor of aurora B kinase), everolimus (mTOR inhibitor), lonafarnib (a farnesyltrasferase inhibitor), MLN-2238 (proteasome inhibitors), and palbociclib (a selective inhibitor of cyclin-dependent kinases). To our knowledge, our study is the first to report synergistic cytotoxicity of resveratrol in combination with everolimus or barasertib, which is specific for cancer cells and particularly for acute lymphoblastic leukemia cells Jurkat. Both combinations are harmless for normal lymphocytes. The data suggest that resveratrol may be a promising supplementary component for anticancer therapy that may allow reduction of the therapeutic doses of barasertib and everolimus, minimizing their side-effects. This statement can also be made for all investigated anticancer drugs, whose cytotoxicity increases by combining them with resveratrol.
Acknowledgements
This study was partially supported by the following projects: Japanese Society for the Promotion of Science (JSPS) (Grand-in-aid “Kakenhi-C” granted to R.B.) and JSPS Fellowship (granted to S.S.), Japan Agency for Medical Research and Development (AMED) (Project for Cancer Research and Therapeutic Evolution, P-CREATE, no. 16 cm0106202h0001), and Bulgarian National Scientific Program ”Young Scientists and Postdoctoral Students“ (granted to D.I.).
Footnotes
Authors' Contributions
ZZ and RB conceived the idea for the manuscript and produced the first draft. DI, SS and RB were involved in the experiments. RB, ZZ and IA were involved in the critical review of subsequent drafts. All Authors read and approved the final manuscript.
Conflicts of Interest
No potential conflict of interest was reported by the Authors.
- Received March 29, 2019.
- Revision received May 5, 2019.
- Accepted May 14, 2019.
- Copyright© 2019, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Docosahexaenoic Acid Potentiates the Anticancer Effect of the Menadione/Ascorbate Redox Couple by Increasing Mitochondrial Superoxide and Accelerating ATP Depletion
- The Complex Composition of Trans-resveratrol, Quercetin, Vitamin E and Selenium Inhibits the Growth of Colorectal Carcinoma
- Effect of Alpha-tocopheryl Succinate on the Cytotoxicity of Anticancer Drugs Towards Leukemia Lymphocytes