


Abstract
Aim: It is possible that macrophages may be effective for cancer treatment because once activated, lung macrophages have enhanced contact with lung tumor cells and have a cytotoxic effect. In this paper, we report that nitric oxide (NO) produced by lung macrophages activated with lipopolysaccharide (LPS) suppressed cell growth of human lung adenocarcinoma cells. Materials and Methods: A549, a lung adenocarcinoma cell line, was cultured with NR8383, a rat alveolar macrophage cell line, in the presence and absence of LPS. The effect of LPS on the growth rate of A549 cells was examined as a function of NO production under cell-to-cell contact conditions. Results: NR8383 cells showed potent cytostatic and cytocidal effects on A549 cells when both cells were co-cultured in the presence of LPS. These effects were mainly due to the production of NO, but another possible mechanism, such as cell-to-cell contact, may also be involved. Conclusion: Activation of alveolar macrophages by LPS suppresses the growth of lung carcinoma cells via NO production under cell-to-cell contact conditions.
- Lipopolysaccharide
- pulmonary inhalation
- lung cancer
- alveolar macrophage
- nitric oxide
- cell-to-cell contact
Non-small cell lung cancer (NSCLC) accounts for 70-80% of lung cancer diagnoses, and the disease is generally diagnosed at an advanced stage. A combination of surgery and chemotherapy is the commonly used treatment for lung cancer, but the long-term survival of patients with NSCLC is low, typically being less than 15% after 5 years (1). Therefore, the development of new therapy against lung cancer is needed.
Macrophages, known to facilitate the action of the immune system in the first step of bio-defense (2), recognize cancer cells as extrinsic materials and try to damage or destroy them. Macrophages are regarded as existing as two phenotypes, i.e. the classical activated macrophages (M1 macrophages) and the alternatively activated macrophages (M2 macrophages) (3). M1 macrophages generate inflammatory cytokines, express high levels of major histocompatibility complex (MHC) molecules, and are cytotoxic towards cancer cells and pathogenic bacteria. In contrast, M2 macrophages, which express scavenger receptors, generate anti-inflammatory cytokines and tissue-repair factors, and play a role in cleaning necrotic tissues, neoangiogenesis, and tissue repair. The phenotype of macrophages changes according to the prevailing conditions. There are tumor-associated macrophages (TAMs) in tumor islets, having migrated there in response to cytokines and chemokines secreted from tumor cells (4). The phenotype of these TAMs is thought to be of the M2 type (5, 6), promoting growth and metastasis of tumor cells (6, 7). On the contrary, it has also been reported that TAMs in tumor islets are favorable for improving prognosis (9, 10). Accordingly, TAMs would be an attractive target of a new cancer immunotherapy. Recently, research on the mechanism of the ‘education’ of M2 macrophages and ‘re-education’ from M2 macrophages to M1 macrophages was attempted (10, 11). Based on the results of these studies, it is expected that appropriate activation of M1 macrophages and conversion of M2 macrophages to M1 macrophages in tumor islets would be helpful for cancer therapy.
In a previous report, we found that activation of lung macrophages (M1 type) by lipopolysaccharide (LPS) increased their adherence to lung tumor cells and caused cytotoxic effect (12). Moreover, this effect was inhibited by the isolation of macrophages from tumor cells using a culture insert tool. These results indicated that the cytotoxic effect of activated lung macrophages on tumor cells occurs by cell-to-cell contact. LPS is known to activate macrophages via toll-like receptor 4 (TLR 4) (13), and to promote the production of nitric oxide (NO) and tumor necrosis factor (TNF)-α by these activated cells (14, 15). Although the usefulness of LPS for cancer therapy has been suggested (16, 17), LPS administered intravenously induces side-effects such as shock and fever. Therefore, activation of TAMs by LPS is worthwhile studying as a treatment for lung cancer.
Materials and Methods
Cell lines. Rat alveolar macrophage cell line NR8383 was purchased from the American Type Culture Collection (ATCC; Manassas, VA, USA; CRL-2192) and was maintained in F-12 Kaighn's Nutrient Mixture (F-12K) medium (Invitrogen Corp., Carlsbad, CA, USA) supplemented with 15% fetal bovine serum (FBS) (Invitrogen Corp.) and 60 μg/ml ampicillin (Meiji Seika Kaisha, Ltd., Tokyo, Japan) at 37°C in a 5% CO2 atmosphere. Human lung adenocarcinoma cell line A549 was obtained from ATCC (CCL-185) and was maintained in F-12K medium supplemented with 10% FBS and 60 μg/ml ampicillin at 37°C in a 5% CO2 atmosphere.
Evaluation of the effect of NR8383 cells activated by LPS on growth rate of A549 cells. A549 cells were cultured in a 96-well plate at a density of 5×103 cells/well and incubated in F-12K medium with 5% FBS for 4 h at 37°C. After completion of adhesion of the cells to the wells, 10 times as many NR8383 cells were added as the number of A549 cells, with or without 1 μg/ml of Escherichia coli LPS (Merck KGaA, Darmstadt, Germany), and the co-cultures were incubated for different numbers of days at 37°C. Then each well was washed with saline to remove NR8383 cells that had not adhered and dead A549 cells. The growth rate was determined by the crystal violet method. The live A549 cells were stained with 0.1% crystal violet/MeOH (Wako Pure Chemical Industries, Ltd., Osaka, Japan) at room temperature for 10 min. After staining, 0.5% SDS (Wako Pure Chemical Industries, Ltd.) was added to each well to solubilize the stained A549 cells. The viable A549 cells were determined by the optical absorbance at 570 nm read by a microplate reader (Model 680; Bio-Rad Laboratories, Hercules, CA, USA), and the growth rate of A549 cells was expressed as a value relative to the total number of A549 cells without treatment with LPS.
Estimation of anti-TNF-α inhibition. A549 cells were co-cultured with NR8383 cells at the cell number ratio of NR8383:A549 of 10:1 in the presence or absence of LPS and in the presence or absence (control) of 10 μg/ml anti-TNF-α (U-CyTech Biosciences, Utrecht, the Netherlands) or 10 μg/ml IgG (isotype control for confirming non specific response of antibody, R&D Systems, Inc., Minneapolis, MN, USA), and then the growth rate of A549 cells (absorbance in the absence of LPS was set at 100%) was determined.
Evaluation of the growth inhibition of recombinant TNF-α and interleukin (IL)-1β, on growth rate of A549 cells. A549 cells were cultured alone for 1, 2 and 3 days in the presence or absence of 6 ng/ml recombinant TNF-α (R&D Systems, Inc.), or 0.3 ng/ml recombinant IL-1β (Wako Pure Chemical Industries, Ltd.), and the growth rate of A549 cells (absorbance in the absence of a humoral factor was set at 100%) was determined.
Effect of NO on A549 cells co-cultured with NR8383 cells. A549 cells were placed in 24-well plates at a density of 3×104 cells/well and incubated in F-12K medium with 5% FBS for 4 h at 37°C. After completion of A549 cell adhesion to the wells, a cell culture insert (pore size, 0.4 μm; Becton, Dickinson and Co., Franklin Lakes, NJ, USA) was inserted into each well. NR8383 cells, with or without 1 μg/ml LPS, were then added to the inner chamber of the insert at a cell number 10 times greater than that of the A549 cells, and the plates were incubated for different numbers of days at 37°C. NO in the incubation chamber containing the A549 cells was determined in terms of nitrite by the Griess method. The growth rate of A549 cells was determined by the crystal violet method described above. To determine the effect of NO on A549 cells co-cultured with NR8383 cells, these cells were incubated together under the conditions described in the section of ‘Evaluation of the effect of NR8383 cells activated by LPS on growth rate of A549 cells’. To examine the effect of the iNOS inhibitor S-methylisothiourea sulfate (SMT; Merck KGaA) on NO production, we added SMT at a final concentration of 50 or 100 μM to co-cultured A549 and NR8383 cells. The cytotoxic effect of NO was assessed by the crystal violet method, and expressed as the value relative to that of the A549 cells without treatment with LPS.
Determination of the effect of NO itself. A549 cells were added to a well in a 96-well plate at a density of 5×103 cell/well and incubated in F-12K medium with 5% FBS for 4 h at 37°C. The NO donor NOR5 (Dojindo Laboratories, Kumamoto, Japan) was added for a final concentration of 200 μM to generate NO, and the cells were incubated for different numbers of days at 37°C. The NO concentration was determined by the Griess method, and the effect of NO on the growth rate of A549 was assayed by using the crystal violet method.
Statistical analysis. Data were analyzed with Student's t-test. Differences between treatment and control groups were considered statistically significant at p<0.05.
Results
Effect of activated alveolar macrophages on lung carcinoma cells. In our previous report, LPS-activated rat alveolar macrophages NR8383 cells attached to human lung carcinoma A549 cells and showed growth inhibition (12). First, we have confirmed the effect in the same system. As shown in Figure 1a, the growth rate of A549 cells in the presence of LPS at 1 μg/ml decreased in a time-dependent manner, after a culture period of 3 days. As shown in Figure 1b, the concentration of nitric oxide (NO) produced by NR8383 cells activated with LPS increased until 3 days, and then leveled off.

Influence of LPS on growth rate of tumor cells and production of NO from alveolar macrophages. A549 cells were cultured with or without NR8383 cells in the presence of LPS for 1, 2 and 3 days, and the growth rates of A549 cells relative to those in the absence of LPS were determined (a). A549 cells were co-cultured with NR8383 cells in the presence or absence (control) of LPS for 1, 2, 3, 5, and 7 days, and the production of NO was determined. The NR8383 cell-to-A549 cell ratio was 10:1. The data are expressed as the mean±S.D. of (a) 9 experiments and (b) 9-17 experiments (*p<0.05 vs. control).
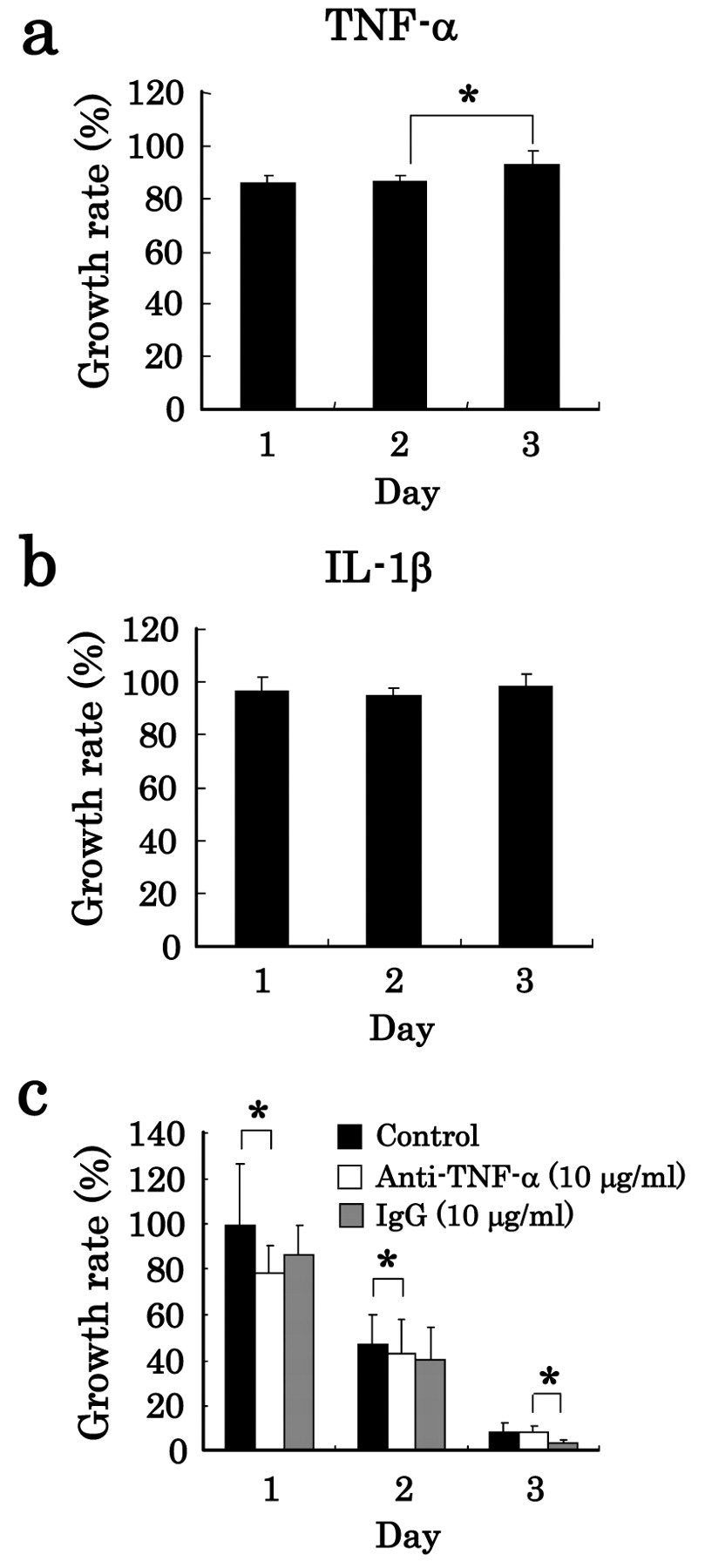
Influence of humoral factor on cytotoxic effect. We already reported that LPS-stimulated NR8383 cells produce significant amounts of TNF-α, IL-1β, and NO (12). Thus, to clarify the influence of cytotoxic factors produced in LPS-stimulated NR8383 cells, we examined the effect of TNF-α, IL-1β, and NO which induce apoptosis of cancer cells (18, 19) on the growth rate of tumor cells. As shown in Figures 2a and 2b, TNF-α and IL-1β did not affect the growth rate of A549 cells. Moreover, we examined the possibility of membrane-bound TNF-α for the growth inhibition by cell-to-cell contact, because membrane-bound type TNF-α is known to behave differently from soluble TNF-α in biological activity and cytocidal activity against several tumor cells in a juxtacrine manner (cell-to-cell contact) (20, 21). As shown in Figure 2c, anti-TNF-α antibody and IgG (isotype control) did not affect the growth rate of A549 cells during co-culture with NR8383 cells, indicating that membrane-bound TNF-α did not affect A549-cell growth.
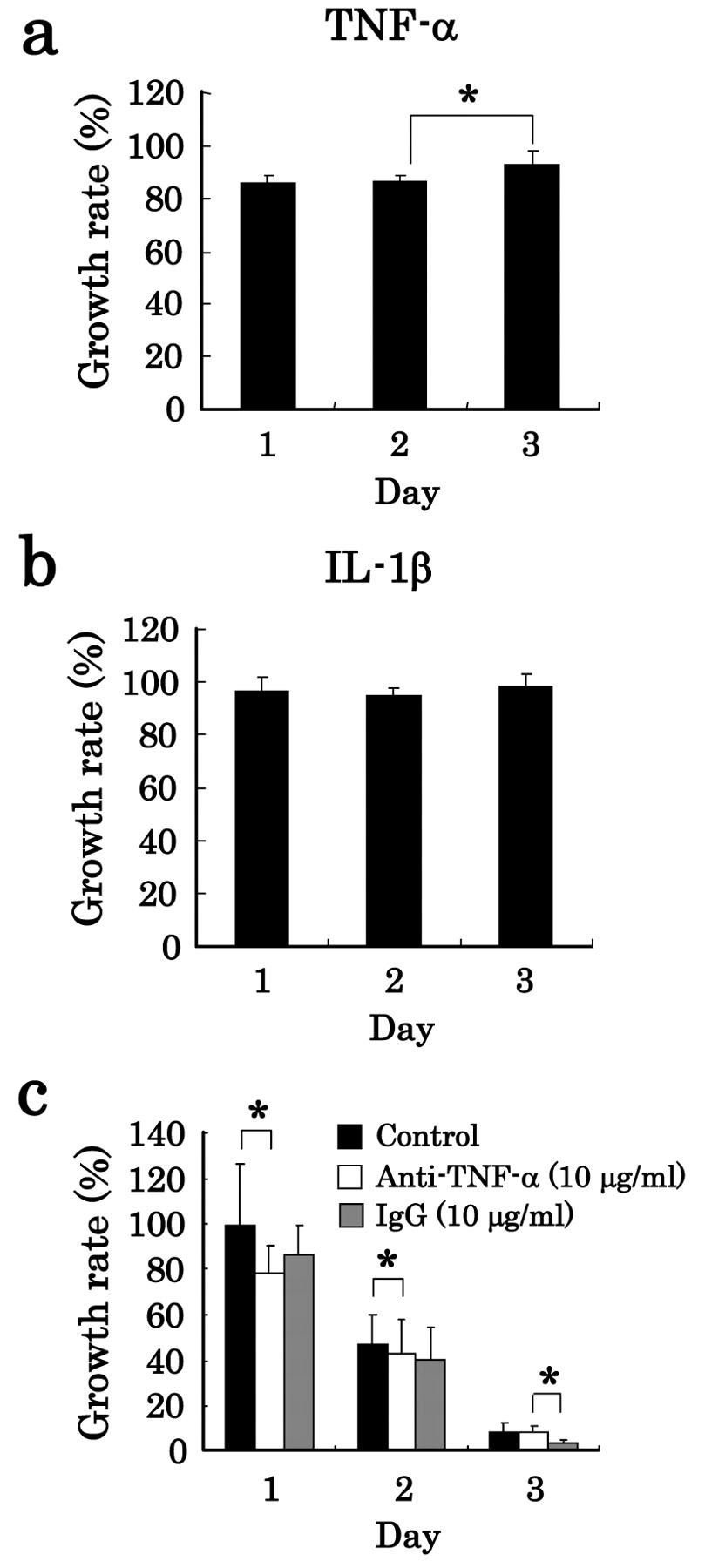
Effects of humoral factors on growth rate of tumor cells. Effects of TNF-α and IL-1β on the growth rate of lung carcinoma cells were determined. A549 cells were cultured alone for 1, 2 and 3 days in the presence or absence of 6 ng/ml TNF-α (a), or 0.3 ng/ml IL-1β (b), and the growth rate of A549 cells (absorbance in the absence of a humoral factor set at 100%) was determined. Furthermore, A549 cells were co-cultured with NR8383 cells at a ratio of NR8383:A549 of 10:1 in the presence or absence of LPS and in the presence or absence (control) of 10 μg/ml anti-TNF-α or 10 μg/ml IgG (isotype control), and then the growth rate of A549 cells (absorbance in the absence of LPS was set at 100%) was determined (c). The data are expressed as the mean±S.D. of (a, b) 6 experiments and (c) 15 experiments (*p<0.05).
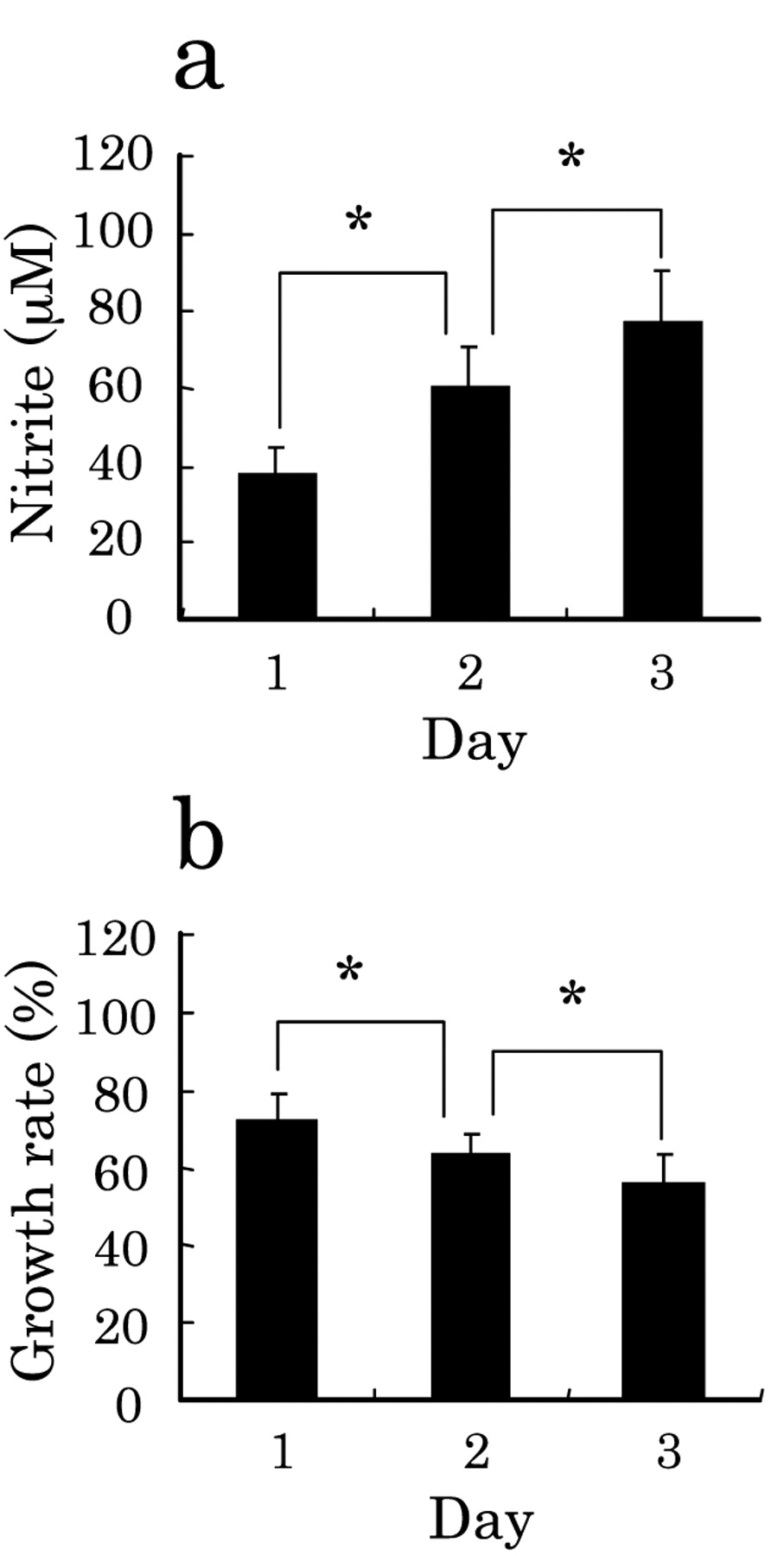
Effects of NO on growth rate of tumor cells. Effect of the humoral factor NO on the growth rate of lung carcinoma cells was determined by culture of A549 cells alone for 1, 2 and 3 days in the presence or absence of the NO generator NOR5, and the production of NO in terms of nitrite (a) and the growth rate of A549 cells (b) (absorbance in the absence of NOR5 set at 100%) were determined. The data are expressed as the mean±S.D. of 15 experiments (*p<0.05).
To determine the effect of NO on the cell growth, we next cultured A549 cells in the presence of NO donor NOR5 at a final concentration of 100 μM for 1-3 days. The concentration of NO produced by NOR5 was determined in terms of nitrate. As shown in Figure 3a, NOR5 produced NO in the incubation medium in such a way that the generation of NO increased with an increase in the duration of culture. In accordance with the production of NO, the growth rate of the A549 cells decreased, falling to approx. 56% on day 3 (Figure 3b). These results suggest that A549 is sensitive to NO, but not to TNF-α and IL-1β.
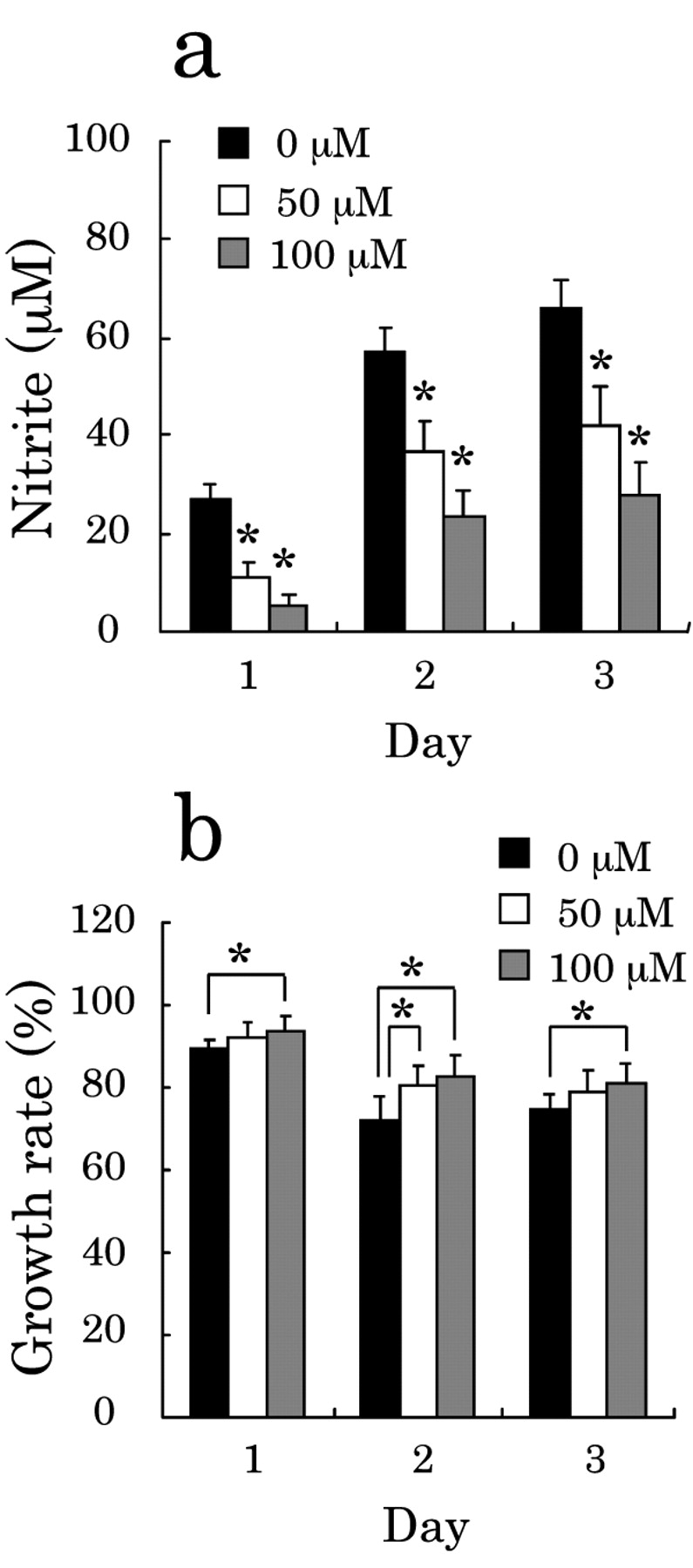
Effect of inhibition of NO production on the growth rate. The results shown in Figures 2 and 3 suggest that the effect of NR8383 cells activated by LPS on the growth rate of A549 cells is due to NO produced from the activated NR8383 cells. Hence, we next examined whether the growth rate of A549 cells was affected by inhibiting the NO production from the LPS-activated NR8383 cells. Namely, A549 cells were cultured with NR8383 cells in the presence of the iNOS inhibitor SMT at a concentration of 50 or 100 μM together with LPS for 1-3 days. As shown in Figure 4a, SMT inhibited the NO production from NR8383 cells in a dose-dependent manner. As a result, the growth rate of A549 cells relative to that without treatment with SMT was increased dose-dependently, and the growth rate of 14% on day 3 in the absence of SMT became approx. 50% in the presence of 100 μM SMT (Figure 4b). Hence, NO was confirmed to be associated with the suppression of the growth rate induced by LPS-activated NR8383 cells on A549 cells.

Effect of NO inhibitor on growth rate of A549 cells co-cultured with NR8383 cells. a: Inhibition of NO production by NR8383 cells by SMT. b: The recovery of growth of A549 cells. Cytostatic effects were evaluated by the growth rate. A549 cells were co-cultured with NR8383 cells in the presence or absence of LPS and in the presence or absence (control) of 50 μM or 100 μM SMT for 1, 2 and 3 days at an NR8383 cell-to-A549 cell ratio of 10:1. The data are expressed as the mean±S.D. of 12 experiments (*p<0.05 vs. control).
Relationship between NO production and its effect on A549 cells cultured separately from NR8383 cells. The previous our results demonstrated that close contact between NR8383 cells and A549 cells was important for induction of a potent suppression of the cell growth (12). In order to clarify this, we next examined the effect of attachment of NR8383 cells to A549 cells on the action of NO, because NO is reported to persist for only a few seconds (22). Namely, A549 cells were cultured separately with NR8383 cells in the presence of LPS and the iNOS inhibitor SMT by using the cell culture insert, and the effect of NO on the growth of A549 cells was determined. As shown in Figure 5a, the production of NO was inhibited dose-dependently by SMT, the NO concentrations thus produced being very similar to those produced in the co-culture of A549 cells with NR8383 cells in the presence of SMT at the corresponding incubation day (see Figure 4a). However, dose-dependent increase in the growth rate of A549 cells by inhibition of NO production was very low, and recovery of the growth rate was only approx. 5% for each day (Figure 5b). Hence, the adhesion of NR8383 cells to A549 cells was also important for the inhibitory effect of NR8383 cells mediated by NO on the growth of A549 cells.

The influence of NO inhibitor on growth rate of A549 cells cultured separately from NR8383 cells. The effects of the iNOS inhibitor SMT treatment using an insert tool on NO production by LPS-stimulated NR8383 cells (a), and the growth rate of A549 cells (b) are shown. A549 cells were cultured separately from NR8383 cells in the presence of LPS and the iNOS inhibitor SMT by using the cell culture insert for 1, 2 and 3 days. After the culture period, the NO concentration in terms of nitrite and growth rate of A549 cells were determined. The results of growth rate are shown by the values relative to those without treatment with LPS. The data are expressed as the mean±S.D. of 12 experiments (*p<0.05 vs. control).
Discussion
In our previous report, we investigated cytotoxic activities including cytostatic effects and cell-killing effects against lung carcinoma cells to establish the therapy for lung cancer through pulmonary administration. Interestingly, LPS-activation of lung macrophages enhanced the attachment to lung tumor cells, resulting in the growth inhibition of the latter (12). It is known that LPS, the main component of the outer membrane of Gram-negative bacteria, is a strong activator of immune systems (23-25). In this study, we examined the effector molecules of alveolar macrophage NR8383 cells activated by LPS on the in vitro growth of the human lung adenocarcinoma A549 cells under cell-to-cell contact conditions.
As a result, LPS had a significant effect on the growth of A549 cells co-cultured with NR8383 cells, showing that activation of alveolar macrophages suppressed cell growth of the lung cancer cells (Figure 1a). When A549 cells were cultured with the supernatant of the incubation medium from cultures of NR8383 cells that had been activated with LPS, the growth of A549 cells decreased considerably, but no growth inhibition was observed at all when the A549 cells were incubated with the supernatant of the incubation medium after culture of A549 cells in the presence of LPS (12). Therefore, it is possible that humoral factors, such as TNF-α, IL-1β and NO, derived from NR8383 cells, are associated with the growth suppression of A549 cells. We found that TNF-α and IL-1β did not suppress the growth of A549 cells co-cultured with NR8383 cells (Figure 2), but NO produced artificially by NOR5 suppressed the growth of A549 cells (Figure 3). Hence, suppression of growth rate of A549 cells is suggested to be due to NO. NO is known to be produced by macrophage cells on stimulation with LPS, and to induce tissue injury, inflammation, and shock, as well as apoptosis (26, 27).
Accordingly, we examined the NO production from NR8383 cells activated by LPS and its effect on A549 cells under cell-to-cell contact conditions. As expected, incubation of NR8383 cells with LPS induced NO production (Figure 1), and the NO produced suppressed the growth of A549 cells, when these cells were incubated either together or separately with NR8383 cells (Figures 4 and 5). However, the decrease in the growth rate induced by LPS was much more significant for A549 cells co-cultured with NR8383 cells than for those cultured with NR8383 cells separately (Figures 1, 4 and 5). In addition, inhibition of NO production by the iNOS inhibitor SMT restored the growth of A549 cells (Figures 4 and 5). These results suggest that close contact of the tumor cells with macrophages was very effective in decreasing the growth of the tumor cells. Activated macrophages are known to migrate to and adhere to tumor cells, where the former suppresses the growth of the latter (12, 28). Therefore, the cellular interaction, such as cell-to-cell contact, should be important for induction of the effect of the LPS-activated NR8383 cells on the growth of A549 cells.
As activated macrophages are reported not to attack normal cells (29), treatment with LPS should be effective for cancer therapy. However, LPS is known to induce tissue injury. In this connection, the effect of LPS from Pantoea agglomerans (IP-PA1) is noteworthy (30, 31). This LPS showed cytostatic and cytocidal effects on A549 cells similar to those obtained by LPS, when it was incubated with NR8383 cells co-cultured with A549 cells, and it was confirmed microscopically that NR8383 cells activated by LPS were localized around A549 cells (12). The polysaccharide part of IP-PA1, containing rhamnose and glucose, is a 5-kDa, being smaller than that of E. coli LPS, whose molecular mass is heterogenous (approx. 10-50 kDa). IP-PA1 is not toxic in animals by oral administration (31, 32), and shows very low toxicity in humans by intracutaneous administration (33, 34). Hence, IP-PA1 is expected to be effective in the treatment and prevention of certain adult diseases, allergy, and infections. Accordingly, IP-PA1, by activating macrophages, could also be a good candidate substance for the therapy of tumors via lung delivery apparatus.
Acknowledgements
This research was partially supported by a ‘High-Tech Research Center’ Project for Private Universities with a matching fund subsidy from the Ministry of Education, Culture, Sports, Science and Technology (MEXT) of Japan, 2004-2008.
- Received June 8, 2010.
- Revision received July 27, 2010.
- Accepted July 27, 2010.
- Copyright© 2010 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}