


Abstract
Aim: The present investigation aimed to examine the therapeutic potential of the new coumarin derivative bis(4-hydroxy-2H-chromen-2-one) coumarin (4HC) against breast cancer. Materials and Methods: For this purpose, the effects of 4HC treatment on the proliferation of MCF-7 breast cancer cells and on MCF-10a non-cancerous cells were evaluated using a fluorescent assay. Cell cycle distribution and apoptosis were measured by image cytometry. The expression level of aromatase (CYP19A1) and apoptosis-related genes were determined by real-time PCR. Results: MCF-7 mammary cancer cell proliferation was significantly decreased within 24 h after treatment with 4HC at 50 μM, while no effect was observed on the viability of MCF-10a non-cancerous mammary cells. 4HC also increased the percentage of the cells in the G2/M phase, inducing apoptosis. Real-time PCR revealed that 4HC induced MCF-7 mortality through an up-regulation of Bax and a down-regulation of Bcl-2, resulting in an increase in caspase-3 gene expression. The increased expression of apoptosis-related genes was accompanied by a decrease in CYP19A1 gene expression. Conclusion: 4HC selectively inhibits proliferation of MCF-7cells in vitro. Moreover, 4HC has inhibitory effects on aromatase gene expression and promoting effects on apoptosis, in MCF-7 cells.
- Coumarin derivative
- Bis(4-hydroxy-2H-chromen-2-one)
- breast cancer
- cell proliferation
- cell cycle
- apoptosis
- aromatase inhibition
- MCF-7 breast cancer cells
Breast cancer is the first cause of cancer-related mortality in women worldwide. Unfortunately, current chemotherapy is responsible for several side-effects and the urgency of development of new anticancer agents to improve treatment of breast cancer, without affecting normal cells, is imperative (1). Coumarins are naturally occurring constituents of many plants and essential oils such as Tonka beans, sweet clover, woodruff, oil of cassia, and lavender (2). Natural and synthetic coumarin derivatives are recognized as important active compounds with beneficial effects on human health including anticoagulant, vasoprotector, antioxidant, anti-inflammatory and antimicrobial activities (3).
In recent years, a significant number of coumarin derivatives have been studied as potential agents for cancer therapy (4), particularly in breast cancer (5). Depending on their structure, they can inhibit tumor necrosis factor-alpha (TNFα) production, cell cycle and aromatase activities or induce apoptosis through the activation of caspases (6, 7). Special attention has been devoted to the new synthetic bis(4-hydroxy-2H-chromen-2one) coumarin, named 4HC, which has been reported to inhibit proliferation and to promote cell death in tumor cell lines (8).
Based on the diverse biological activity reported by coumarin derivatives, the anti-tumor potential and pro-apoptotic activities of 4HC have been investigated in MCF-7 estrogen receptor-positive (ER+) breast cancer cells, a widely-used model system for the study of breast cancer (9). The induction of apoptosis in tumor cells is an important regulated process tightly involved in therapy and the prevention of cancer.
The impact of 4HC on aromatase activity has also been evaluated. Indeed, breast cancer is known to be under hormonal control and the inhibition of estrogen synthesis by aromatase inhibitors results in the reduction of breast cancer cell proliferation and the induction of apoptosis (10). Several studies reported that coumarin derivatives could be potential aromatase inhibitors and highlighted their efficiency for breast cancer therapy (11). Based on all the above, the expression of aromatase (CYP19A1 gene) was quantified in order to provide insight into the mechanism of the pro-apoptotic effects of 4HC. Finally, we demonstrated that 4HC could be considered as a novel chemotherapeutic agent for breast cancer.
Materials and Methods
Chemistry. Synthesis of bis (4-hydroxy-2H-chromen-2-one) coumarin, namely 4HC, was performed according to the previously described method (8) (Figure 1). Structural identification of 4HC was elucidated by extensive one-dimensional (1D) and two-dimensional (2D) nuclear magnetic resonance (NMR) techniques. The stock solutions (10 mM) of 4HC were dissolved in DMSO, preserved at −4°C and diluted in a culture medium to realize the experiments.
Cell culture. The human breast cancer cell line MCF-7 and the human non-tumorigenic epithelial cell line MCF-10a were obtained from the American Type Culture Collection (Molsheim, France). MCF-7 cells were cultured in RPMI-1640 (GIBCO, Invitrogen, Carlsbad, CA, USA) containing 10% heat inactivated fetal bovine serum (FBS) (Eurobio, Courtaboeuf, France), 1% L-glutamine (2 mM) (GIBCO), 0.5% gentamycin (50 mg/ml) (Fisher Scientific, Strasbourg, France) and 0.04 UI/ml of insulin (Sigma-Aldrich, Saint-Quentin-Fallavier, France). MCF-10a cells were cultured in DMEM/F-12 medium (GIBCO) supplemented with 10% heat-inactivated horse serum (Pan Biotech, France), EGF (0.02 μg/ml), hydrocortisone (0.5 μg/ml), insulin (0.25 UI/ml) and cholera toxin (0.1 μg/ml) (Sigma-Aldrich). All cells were kept at 37°C under a 5% CO2 atmosphere.
Cell proliferation analysis by resazurin assay. Human breast cancer cells MCF-7 and non-cancerous mammary cells MCF-10a were seeded in 96-well plates (1×104/well) in complete media 24 h before treatment with 4HC. 4HC was added at concentrations ranging from 0 to 100 μM for 24, 48, and 72 h. At the end of the treatment, 200 μl of a 25 μg/ml solution of resazurin (Sigma-Aldrich) was added to each well for 2 h of incubation at 37°C. Control wells received the vehicle solvent (0.1% v/v DMSO) and doxorubicin (1 μM) was used as a positive control. Fluorescence was measured on an automated 96-well plate reader (Fluoroskan Ascent FL, Thermo Fisher Scientific, Wilmington, DE, USA) and data were normalized to the control group fluorescence.
Cell cycle analysis. MCF-7 cells were seeded into 12-well plates (1×105 cells/well) for 24 h and synchronized by arresting the cells in the G0 phase, through serum deprivation for an additional 24 h. Cells were then treated for 72 h with 4HC (0, 25 μM or 50 μM). After trypsinization and centrifugation, cell pellets were resuspended in 200 μl of PBS for a final concentration of 5×106 cells/ml. After storage at −20°C, cells were centrifuged and stained with 50 μg/ml of propidium iodide (PI) (Sigma-Aldrich) and 100 μg/ml of ribonuclease A (Sigma-Aldrich). After incubation, cells were analyzed using Cellometer Vision K2 (Nexcelom Bioscience, Lawrence, MA, USA). Data were analyzed using the FCS express 6 image Cytometry software (De Novo software). Experiments were repeated in triplicate.
Primer sequences.
Apoptosis detection by Annexin V-FITC/PI staining. Apoptotic cells were quantified by Annexin V-FITC/PI (Sigma-Aldrich) staining according to the manufacturer's protocol. In brief, cells in a 12-well plate were treated with 4HC at a concentration of 100 μM for 72 h. Then cells were washed with PBS, harvested with trypsin and suspended in 50 μl of Annexin V Binding Buffer. Protected from light, Annexin V-FITC conjugate and PI were added to cells. After 15 min of incubation at room temperature, fluorescence of the cells was read immediately using image cytometer (Cellometer Vision K2, Nexcelom Bioscience, Lawrence, MA, USA). Data were analyzed using the FCS express 6 image Cytometry software (De Novo software). Experiments were repeated in triplicate.
Total RNA extraction. RNA extraction was performed using trizol (Sigma Aldrich). MCF-7 cells were seeded in a 12-well plate (1×105 cells/well) and were treated with 4HC at a concentration of 50 and 100 μM or with vehicle for 72 h. After treatment, trizol was added in each well for total cell lysis. The mixture was separated by chloroform in three phases and RNA was collected in the aqueous phase. The RNA was precipitated with isopropanol then were frozen overnight at −20°C. Centrifugation at 12,000 ×g for 15 min allowed recovering RNA pellets, which were washed with 70% EtOH. After a second centrifugation, the supernatants were carefully removed and samples were suspended in DEPC-treated water. The RNA concentration and purity was determined using a spectrophotometer (Nanodrop 2000; Thermo Scientific).
Real-time polymerase chain reaction (qPCR). After a DNase treatment (DNase I Amplification grade, Invitrogen), 1 μg of RNA was used to synthesis cDNA (HighCap cDNA RT Kit with RNAse inhibitor, Invitrogen) following the manufacturer's protocol. The expressions of Bax, Bcl-2, Caspase 3 and CYP19A1 were evaluated by real-time qPCR. The reverse-transcribed products (2 μl) were added to the PCR mixture (18 μl) for 40 cycles. Each cycle included 15 seconds at 95°C and 1 min at 60°C. Gene-specific primers are listed in Table I. GAPDH mRNA level was used for normalization. The relative quantification method (RQ=2−ΔΔCT) was used to calculate the relative gene expression of given samples with ΔΔCT=[ΔCT (sample1) - ΔCT (sample2)] and ΔCT=[CT(target gene) −CT(reference gene)].

Synthesis of bis(4-hydroxy-2H-chromen-2-one) coumarin (4HC).

Proliferation of MCF-10a mammary non-cancerous cells (A) and MCF-7 mammary cancer cells (B) treated with 4HC. 4HC showed significant anti-proliferative activities in MCF-7 cells in time and concentration-dependent manner. *p≤0.05; **p≤0.01; ***p≤0.001 compared to the control value. Experiments were repeated in triplicate. Ctl, Control; Doxo, doxorubicin.
Data analysis. Data were expressed as the mean±standard deviation (SD) and represented at least three independent experiments. Statistical analysis and significance were measured by the Student's t-test using SPSS software version 23.0. Differences at p≤0.05 (flagged as *) or p≤0.01 (flagged as **) or p≤0.001 (flagged as ***) were considered statistically significant.
Results
4HC inhibited breast cancer cell proliferation. In order to investigate the antiproliferative characteristics of 4HC, MCF-7 breast cancer cells and MCF-10a non-cancerous mammary cells were treated with increasing concentrations of 4HC for 24, 48, and 72 h and cell viability was measured as previously described. Results showed that concentrations up to 50 μM of 4HC did not affect the viability of non-cancerous mammary cells MCF-10a, while a slight decrease was observed at 100 μM of treatment (Figure 2A). On the contrary, 4HC significantly decreased the proliferation of MCF-7 breast cancer cells in a dose and time dependent manner (76.15% and 88.46% inhibition at 50 μm and 100 μm, respectively, after 72 h of treatment; p≤0.001, compared to the control group) (Figure 2B). In addition, the sensitivity of MCF-7 cells exposed at 100 μM of 4HC was similar to that observed with the doxorubicin treatment (88.46% of inhibition with 4HC at 100 μM and 90.73% of inhibition with doxorubicine, compared to the control, p≤0.001).
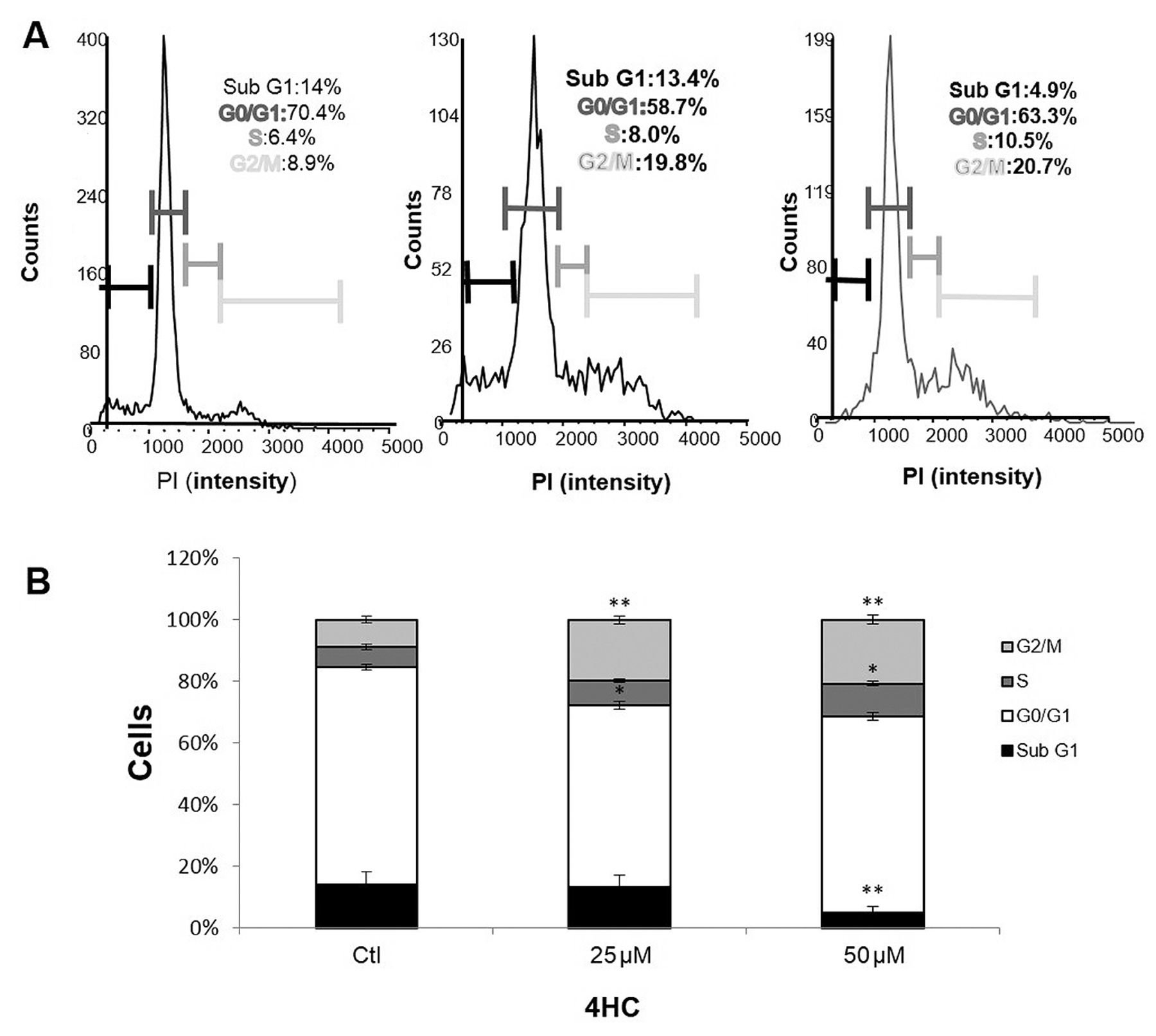
MCF-7 cell cycle distribution analysis (A). The percentage of cells in the G1, S, and G2/M phases of the cell cycle are shown, following a treatment with 25 and 50 μM of 4HC for 72 h (B). *p≤0.05; **p≤0.01 compared to the control group. Experiments were repeated in triplicate. PI, Propidium iodide; ctl, control.
4HC induced cell cycle arrest in G2/M phase. In order to demonstrate that the growth inhibition of MCF-7 cells induced by 4HC could be associated to cell cycle arrest, we analyzed cell cycle progression using the Cellometer K2 Fluorescent Cell Viability Counter (Figure 3A). MCF-7 cells were treated with 0, 25 and 50 μM of 4HC for 72 h. As shown in (Figure 3B), the percentage of cells in the G2/M phase significantly increased from 8±0.98% (control condition) to 19±1.3% (p<0.01) and 20±1.5% (p<0.01) when cells were treated with 25 and 50 μM of 4HC, respectively. These results were accompanied by a decrease in the Sub G1 phase distribution (from 14% in the control group to 4.9% in the 50 μM 4HC-treated group, p≤0.01).
4HC significantly promoted apoptosis in MCF-7 cells. To characterize cell death and assess the pro-apoptotic activity induced by 4HC in MCF-7 cells, we performed PI-annexin-V double staining assays. The results demonstrated a significant increase in the late apoptosis rate in MCF-7 cells after 72 h of treatment with 100 μM of 4HC, compared to the control group (58.75±2.55% and 8.55% respectively, p≤0.01; Figures 4A and 4B).
Activation of the pro-apoptotic genes Bax and Caspase 3 and down-regulation of Bcl-2 were associated with the inhibition of CYP19A1. We performed q-RT-PCR to explore the probable mechanism whereby 4HC induced apoptosis. Analysis revealed (Figure 5) that 4HC treatment induced a significant decrease of CYP19A1 gene expression (0.37±0.1-fold; p≤0.05, compared to the control group), accompanied by a significant up-regulation of pro-apoptotic genes when the concentration of 4HC was equal to 100 μM (Bax: 3.65±0,39-fold; Caspase 3: 5.13±0.41-fold; all p-values were ≤0.05, compared to the control groups), and a significant down-regulation of the anti-apoptotic gene Bcl-2 (0.21±0.1; p≤0.01). These data suggested that inhibition of aromatase may play an important role in inducing apoptosis.

Evaluation of apoptosis in MCF-7 cells treated with 4HC. Images (captured by FCS express 6 image Cytometry, De Novo software) showed apoptosis in MCF-7 cells. Control and 4HC-treated cells for 72 h were stained with Annexin V-FITC and PI. (FITC-/PI-) reflected living cells, fluorescence (FITC+/PI-) represented early apoptotic cells, and fluorescence (FITC+/PI+) showed late apoptotic cells (A). The percentage of apoptosis is represented as the mean±SD (B). *p≤0.05; **p≤0.01, compared to the control. Experiments were repeated in triplicate. Ctl, Control.
Discussion
Previous studies have reported the beneficial therapeutic strategy in the use of cytotoxic drugs, which destroy cancer cells by the inhibition of aromatase and the induction of apoptosis in the cancer cells (12, 13). In this context, our study evaluated the anti-tumor effects of the new synthesized bis(4-hydroxy-2H-chromen-2-one) coumarin (4HC) on human breast cancer cells, in vitro. We aimed to investigate the effect of 4HC on the proliferation and apoptosis of MCF-7 breast cancer cells and non-cancerous cells MCF-10a as well. Moreover, the study focused on the impact of 4HC on the regulation of aromatase and apoptosis-related genes.
Obviously, our results showed an antitumor activity of 4HC with a significant inhibition of the proliferation of human breast cancer cells MCF-7 in a dose and time dependent manner. Significant cytotoxic effects were observed after 24 h of treatment at 50 μM. Previous studies have shown a similar antiproliferative activity related to the presence of hydroxyl group at position 4 against other cancer cell lines (14, 15). Even more interesting, we demonstrated the ability of 4HC to inhibit cell viability without significant cytotoxicity on hyperplastic non-cancerous cells. These data are in agreement with earlier reports demonstrating selectivity of coumarin (16) and especially 4HC toward K-562 and the leukemic JURKAT cell lines (8).
Among the potential mechanisms that have proven effective for the treatment of cancer is the blockade of the cell cycle (17). In our study, cell cycle analysis revealed that 4HC caused arrest in the G2/M phase, in a concentration dependent manner, associated with a decrease in the G1 phase. Many previous studies have demonstrated the ability of coumarin derivatives to induce cell cycle arrest and apoptosis in other cancer cell lines (18-21), supporting our results. More specifically, similar effects on MCF-7 cell cycle progression were induced by the thiazolylpyrazolyl coumarin derivative 9d (20), and a significant increase in the G2/M-phase cell population followed by reduction of cells in G1-phase was also observed in ovarian cancer cells after treatment with the coumarin derivative RKS262 (21).

The relative expression of the pro-apoptotic Bax, and caspase 3 genes, the anti-apoptotic gene Bcl-2 and the aromatase gene CYP19A1 levels in 4HC-treated and control MCF-7 cells (for 72 h) were analyzed using quantitative real-time reverse transcription polymerase chain reaction (qRT-PCR). *p≤0.05; **p≤0.01, compared to the control group. Experiments were repeated in triplicate. Ctl, Control.
Moreover, based on previous evidence about the essential role of the cell cycle in the regulation of cell growth and mediation of apoptosis (22-24), we decided to explore whether the cell cycle arrest induced by 4HC led to changes in apoptosis. Apoptosis is the process of programmed cell death and constitutes a target for variety of drugs (25-27). Referring to previous studies (18, 28), we used Annexin-V/PI staining to detect apoptosis and, a significant increase in apoptosis was observed in MCF-7 cells after treatment with 4HC for 72 h.
Bax and Bcl-2 proteins represent the most important effectors of apoptosis and play a pivotal role in the regulation of cell death (29, 30). Alteration in the ratio of Bax/Bcl2 is a crucial factor in apoptosis induction (31, 32). A significant down-regulation of Bcl-2 and an up-regulation of the Bax gene expressions after 4HC treatment reflect a decrease in the Bcl-2/Bax ratio which could result in the activation of the caspase apoptosis proteins (33). Caspases, like caspase 3, are cysteinyl aspartate-specific proteinases playing a critical role in apoptosis and provide a target for therapies (34). Our study results support these findings and demonstrated that 4HC induced overexpression of caspase-3 gene, which is required for regulation of apoptosis.
Based on all of the above, and in order to define the underlying mechanisms of 4HC apoptosis, we focused on the inhibition of aromatase. Aromatase inhibitors have been reported to block cell proliferation and promote signal transduction pathways of cell death (35). In addition, the detailed mechanism under which aromatase blocking induced cell cycle arrest, activation of caspases and promotion of apoptosis has already been defined (36). Ours results revealeed that 4HC has also anti-aromatase potency by inducing down-regulation of CYP19A1 gene expression, which could lead to the cell cycle arrest and promotion of apoptosis observed in MCF-7 cancer cells. These findings further support a previous report which demonstrated the aromatase inhibitor potential of coumarin derivatives in breast cancer cell lines (37).
In summary, our study demonstrated for the first time that 4HC selectively inhibited the proliferation of MCF-7 breast cancer cells and induced cell cycle arrest leading to the activation of apoptosis. Furthermore, our results suggested that down-regulation of aromatase gene may be a potential mechanism responsible for 4HC-induced cell death, resulting in a decrease in Bcl-2, an increase in Bax, and caspase-3 gene expression. Based on these encouraging findings, the inhibition of aromatase activity using 4HC might be a promising strategy in breast cancer treatment.
Acknowledgements
The Authors would like to thank Université Clermont Auvergne, INRA, UNH, Unité de Nutrition Humaine, CRNH Auvergne, the University of Aveiro, Fundação para a Ciência e a Tecnologia (Portugal), EU, QREN, FEDER, COMPETE and Scientific and Technical Research Center in Physico-Chemical Analysis (CRAPC) for allowing us the achievement of this work.
Funding: This work was financially supported by Université Clermont Auvergne, Equipe ECREIN (France) and the University of Aveiro (Portugal). Algeria's General Directorate for Scientific Research and Technological Development (DGRSDT) also made it contribution.
Footnotes
↵* These Authors contributed equally to this work.
Authors' Contributions
OT, AS and KB performed the chemical experiments, LHR, MV and AB performed the biological experiments. LHR, CD, MPV, LD and FCC analyzed the data and wrote the manuscript.
Conflicts of Interest
The Authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
- Received September 10, 2019.
- Revision received October 11, 2019.
- Accepted October 16, 2019.
- Copyright© 2019, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}