


Abstract
Background/Aim: The rabbit auricular VX2 carcinoma is an established animal model for human head and neck squamous cell carcinoma (HNSCC). Previously, we observed that intraperitoneal oxidative (O3/O2) stress induced tumor remission. Our aim was to evaluate candidate genes associated with tumor regression. Materials and Methods: For identification of tumor remission-related genes, microarray analysis was performed with subsequent validation by polymerase chain reaction (PCR), in situ hybridization, immunohistochemistry and western blot analysis. Results: Microarray analysis indicated a prominent reduction of epidermal growth factor receptor (Egfr, Erbb1) expression levels in regressing tumors. Quantitative PCR confirmed a significant (p<0.005) down-regulation of Erbb1-3 mRNA in regressing VX2 tumors. Histological localization of Erbb1-3 mRNA transcript and protein indicated reduced Erbb gene expression occurring at the level of individual VX2 tumor cells rather than solely being an effect of tumor shrinkage. This study highlights changes in the Erbb gene signature of regressing VX2 carcinomas as a predictor for therapy response. The VX2 carcinoma animal model, therefore, appears suitable for the identification and evaluation of new diagnostic, prognostic and therapeutic biomarkers prior to their application in patients with HNSCC.
The epidermal growth factor receptor (EGFR) family comprises four distinct receptor tyrosine kinases (RTKs): EGFR (also known as ERBB1/HER1), ERBB2 (HER2, neu), ERBB3 (HER3) and ERBB4 (HER4). ERBB receptors trigger intracellular signals promoting cellular differentiation, proliferation, survival and migration thereby being critical in embryonic development, tissue repair and renewal, as well as in cancer biology. Since first reports demonstrated EGFR to be related to the transforming protein encoded by the v-erb-B oncogene of avian erythroblastosis virus (1), the role of ERBB receptors in cancer development has been studied intensely (2). In fact, aberrant ERBB activity has been described in a variety of human epithelial cancer types (2, 3) including head and neck squamous cell carcinomas (HNSCC) (4), which are the most frequent malignancies of the upper aerodigestive tract. Overexpression of EGFR has been verified in up to 90% of all HNSCC (5, 6) and numerous studies have demonstrated that EGFR can be used as a prognostic factor to predict tumor growth, therapy sensitivity and risk of recurrences (7-9). Overexpression of other ERBB family members, such as ERBB2 and ERBB3 have been described in ovarian and breast cancer as well as HNSCC (10-13).
In order to identify candidate genes and unravel molecular and cellular changes present in regressing VX2 tumors, we experimentally deployed the auricular VX2 carcinoma animal model of the New Zealand White (NZW) rabbit. This animal model exhibits analogous growth features to those observed in HNSCCs (14, 15). Similar to human papillomavirus-related HNSCCs, the VX2 carcinoma is also associated with a papillomavirus, the so-called Shope cottontail rabbit papillomavirus (16, 17). In the VX2 model, a squamous cell tumor develops after implantation of VX2 tumor cell suspension into the subcutis of the rabbit auricle, which later tends to metastasize regionally to cervical lymph nodes, as well as distantly to organs such as the lung. This pattern of metastatic spread of the VX2 tumor strongly resembles the biological behavior of human HNSCC and is one main reason why the VX2 tumor has become a valuable tool in head and neck cancer research.
Previously, we reported that a strong oxidative stress applied intraperitoneally (O3/O2-pneumoperitoneum, O3/O2-PP) to VX2 carcinoma-bearing rabbits resulted in remission of the tumor and prevention of associated metastasis in a significant number of animals (18). The exact underlying mechanisms of this therapeutic approach are still unknown, but given that NZW rabbits are equipped with an intact immune system, the body's own immune system is most likely involved since reimplantation of the VX2 tumor in cured rabbits was only possible after immune suppression with dexamethasone and cyclosporine A (19). The role of the immune system was further supported by the observation that adoptive transfer of leukocytes from rabbits with regressive VX2 carcinomas into those with progressive tumor led to tumor remission in the recipient animals (16).
The aim of the study was therefore to evaluate in more detail the expression level of EGFR and other members of the ERBB family of RTKs in progressive and regressive VX2 tumors.
Materials and Methods
Animals. Animal experiments were approved by the regional board in Giessen, Germany [V54-19c20-15(1) MR20/26 Nr.A22/2008 and V54-19c20-15(1) MR20/26 Nr.34/2011] according to the German Animal Protection Law. Overall 28 adult Iffa Credo NZW rabbits (Charles River, Wiga, Germany) with a body weight (BW) range of 2.0 to 3.0 kg were used; four animals served as donors of the VX2 cell suspension used for the induction of auricular tumors. Rabbits were kept in individual steel cages under standardized air conditioning (20-22°C, 50-60% humidity) under a 12 h artificial day/night rhythm with access to food and water ad libitum. Animals were acclimatized for at least 7 days in the hutch prior to starting the experiment. For surgical ablation of the auricular tumor, animals were sedated by Robinul® (0.1 ml/kg BW; Riemser Arzneimittel AG, Greifswald, Germany) subcutaneously (s.c.) and anesthetized by a mixture of Rompun® (5 mg/kg BW; Bayer Vital GmbH, Leverkusen, Germany)/Ketavet® (70 mg/kg BW; Pharmacia GmbH, Berlin, Germany) intramuscularly. Analgesic treatment s.c. with Temgesic® (Essex Pharma GmbH, Munich, Germany) was started 2 h before surgical intervention and maintained for at least 2 days and dependent on wound-healing progression. Signs of distress, pain or cachexia, defined as weight loss above 20%, were criteria for euthanasia.
Experimental design. Auricular VX2 tumors were implanted into the left ear of 24 NZW rabbits by s.c. injection of 5×107 tumor cells derived from a donor rabbit as described elsewhere (19). Tumor volume was measured daily with a digital caliper and growth was allowed to continue until the solid auricular tumor had reached a volume of >2,500 mm3. At this stage, the animals were alternately assigned to one of two experimental groups: i) animals that received intraperitoneal oxidative stress (O3/O2-PP); or ii) sham-treated animals that received anesthesia and a puncture into the peritoneum but without gas insufflation.
The O3/O2-PP therapy consisted of daily intraperitoneal insufflation of a medical O3/O2 gas mixture (MedozonIP; Herrmann Apparatebau GmbH, Kleinwallstadt, Germany) under anesthesia (70 mg/kg ketamine/20 mg/kg xylazine) over a period of 5 consecutive days. The O3/O2 gas mixture was applied in a standardized volume of 80 ml/kg BW containing 50 μg O3 per ml gas mixture corresponding to 2.5% O3 and 97.5% medical O2. For a detailed description see Schulz et al. (19).
Tumor size was documented daily with a digital caliper and tumor development was classified as progressive when the tumor volume reached more than 6,000 mm3, or designated as in remission when the volume dropped below 50% of its maximal value. These volumes were based on previous personal observations that once a tumor decreased or increased to one of these levels it either totally disappeared or progressively grew leading to the animal's death within 2 months due to massive tumor bleeding or abundant lung metastases. Once an auricular VX2 tumor reached these criteria it was surgically removed as described earlier (18) and tissue samples were stored in RNAlater (Sigma-Aldrich, Darmstadt, Germany) to be used in downstream applications such as quantitative reverse transcription (qRT) - PCR and microarray analysis. In addition, tissues were cryoprotected in Tissue-Tek® O.C.T. Compound (Sakura Finetek, Torrance, CA, USA) for in situ hybridization (ISH) analysis, or fixed in 4% formaldehyde solution (i.e. formalin) to be used in formalin-fixed paraffin-embedded (FFPE) immunohistochemistry.
Microarray analysis. Total mRNA was isolated from two progressive and two regressive auricular VX2 tumors using RNeasy Mini kit (Qiagen, Hilden, Germany) according to the manufacturer's protocol. RNA was validated with the Experion™ Automated Electrophoresis System (BioRad Laboratories Inc., Dreieich, Germany) and subsequently deployed for microarray analysis (one slide, four microarrays/slide; Format: 4×44K, product #: G2519F, Design ID: 020908; Agilent Technologies, Santa Clara, CA, USA). This microarray represents 42,034 gene fragments that are based on gene information obtained from the RefSeq (Release 29, May 2008), Unigene (Build 11, Mar 2008) and Ensembl (Release 49, Feb 2006) databases. Microarray analysis, as well as pre-evaluation of the raw gene-expression data, were carried out at the EMBL GeneCore Facility (European Molecular Biology Laboratory, Heidelberg, Germany).
Quantification of Erbb and mitogen-activated protein kinase kinase 1 (Mapk2k1) mRNA in VX2 tumor tissues. All tumor tissues were homogenized with Precellys® Homogenizer (Peqlab Biotechnologie GmbH, Erlangen, Germany). Total RNA was isolated using RNeasy Mini kit (Qiagen) according to the manufacturer's protocol and single-strand cDNA was synthesized from 0.5 μg total RNA using RT2 first strand kit (Qiagen). For Erbb and Mapk2k1 mRNA quantification by qRT-PCR, Power SybrGreen PCR Master Mix (Applied Biosystems, Darmstadt, Germany) was used in combination with the respective primer pairs (Invitrogen, Darmstadt, Germany) as listed in Table I. PCR was performed with 1 cycle at 95°C for 10 min, 40 cycles at 95°C for 15 s and 60°C for 15 s) afterwards followed by a dissociation curve with 95°C for 15 s, increasing temperature from 60 to 90°C and afterwards 1 cycle at 95°C for 10 min. Rabbit Erbb mRNA was normalized to the respective amount of rabbit glyceraldehyde 3-phosphate dehydrogenase (Gapdh; NM_001082253.1) mRNA in each sample. Samples were run in triplicate. The cycle threshold (Ct) cut-off value for all measurements was 35. For relative quantification, an efficiency of 100% was assumed for all primer pairs. For absolute quantification, copy numbers of Egfr, Erbb2, Erbb3, Erbb4 and, Gapdh were quantified by using a standard curve of known cDNA copy numbers.
Summary of oligonucleotide primer pairs used in this study.
Cloning of Erbb DNA probes. Egfr-, Erbb2-, and Erbb3-specific amplicons were generated by RT-PCR from RNA derived from auricular VX2 tumor tissue using Phusion Hot Start II DNA Polymerase (New England Biolabs, Frankfurt, Germany) in combination with the respective ISH primers (eurofins, Nuremberg, Germany) as listed in Table I. The resulting amplicons were gel purified using NucleoSpin™ Gel and PCR Clean-up Kit (Macherey & Nagel GmbH, Düren, Germany) and cloned into pGEM-T easy vector using the pGEM®-T Vector System I (Promega GmbH, Mannheim, Germany). The vector containing the respective Erbb-specific amplicon was used to transform competent NEB®10-beta Escherichia coli cells (New England Biolabs) following the manufacturer's protocol. Single clones were generated and Egfr, Erbb2, and ErbB3 recombinant vectors were purified (QIAprep Spin Miniprep Kit; Qiagen). The level of sequence identity of the cloned Egfr, Erbb2, and Erbb3 amplicons to the respective reference sequences was confirmed by sequence analysis (Seqlab, Goettingen, Germany).
Preparation of Egfr-, Erbb2-, and Erbb3-specific cRNA probes. Recombinant plasmids were linearized with the appropriate restriction enzymes (NotI or NcoI; New England Biolabs) and transcribed to antisense or sense cRNA using SP6 or T7 polymerase, respectively. In vitro transcription was performed in the presence of 35S-UTP (New England Nuclear, Dreieich, Germany) followed by treatment with RNase-free-DNase (Qiagen) for 15 min at 37°C. To generate fragments of approximately 250 bp (for optimal intracellular translocation of the probes), 35S-Labeled cRNAs were hydrolyzed in sodium carbonate (final concentration 0.1 M, pH 10.2) at 60°C according to the protocol provided by Angerer et al. (20). Hydrolysis was stopped by the addition of acetic acid to a final concentration of 0.5% (v/v), followed by immediate purification of the probes using Micro Bio-Spin Chromatography columns as described in the instruction manual (Bio-Rad, Munich, Germany). For ISH, all labeled cRNA probes were diluted in hybridization buffer (100 mM Tris pH 7.5, 600 mM NaCl, 1 mM EDTA, 0.5 mg/ml tRNA, 0.1 mg/ml sonicated salmon sperm DNA, 1x Denhardts, 10% dextran-sulfate, 50% formamide) to a final activity of 50,000 cpm/μl. All ISH assays were performed with both antisense and sense cRNA probes.
In situ hybridization. Serial 14 μm cryostat sections of tumor tissues were fixed in 4% formaldehyde at 4°C for 1 h, washed three times in phosphate-buffered saline (PBS), penetrated by 0.4% Triton X-100 in PBS for 5 min and acetylated for 10 min in 0.1 M triethanolamine (pH 8.0) with 0.25% acetic anhydride. Tissues were washed 2x in saline-sodium citrate (SSC) and dehydrated in ethanol. Hybridization of cellular mRNA with Egfr-, Erbb2-, and Erbb3-specific cRNAs, prepared by in vitro transcription, was achieved after incubating 35 μl of cRNA on tissue sections for 16-18 h at 52°C in a moist chamber. Sections were washed in 2x SSC and 1x SSC for 10 min, followed by digestion of single-stranded non-hybridized RNA with 10 μg/ml RNAse A and 1 U/ml T1 RNAse (both Roche, Mannheim, Germany) in the presence of Tris/EDTA (pH 8.0), and 150 mM NaCl for 1 h at 37°C. Subsequently, sections were desalted with 1x SSC, 0.5x SSC and 0.2x SSC for 10 min followed by incubation in 0.2x SSC for 1 h at 60°C. Finally, the tissue sections were washed in water for 10 min, dehydrated with ethanol and air dried. Autoradiograms were generated by exposing the sections to autoradiography film (Hyperfilm-MP; Amersham, Dreieich, Germany) for 1-3 days, followed by coating of sections with Kodak NTB2 film emulsion (Fisher Scientific GmbH, Schwerte, Germany). Coated tissues were developed with D19 Kodak developer (Fisher Scientific GmbH) and fixed with Ilford Hypam Fixer (llford, Cheshire, UK) after 15-25 days and counterstained with hematoxylin/eosin. Sense probes served as a negative control and never generated any signal above the background level.
Antibodies. Anti-EGFR (cetuximab, Erbitux®; 2 μg/μl, dilution 1:100; Merck KGaA, Darmstadt, Germany); anti-ERBB2 [Neu (F11)] and anti-ErbB3 [ErbB3 (G-4)] (both from Santa Cruz Biotechnology Inc. Heidelberg, Germany) and anti-mitogen-activated protein kinase, activated diphosphorylated extracellular signal-regulated kinase 1/2 (ERK1/2) (monoclonal, clone MAPK-YT; Sigma-Aldrich) were deployed in immunohistochemistry. Anti-human IgG [Biotin-SP-AffiniPure F(ab')2 Fragment Goat anti-Human IgA + IgG + IgM (H+L); Dianova GmbH, Hamburg, Germany] was used to detect the humanized chimeric antibody cetuximab. The EnVision+ System horseradish peroxidase (HRP)-labeled polymer (K4001; Dako, Glostrup, Denmark) was deployed to detect mouse antibodies directed against ERBB2 and -3 and active ERK1/2 during immunohistochemistry. Primary antibodies directed against active ERK1/2 and beta-actin (mouse monoclonal; Sigma) were used for western blot analysis followed by their detection with HRP-conjugated mouse IgG kappa binding protein (Santa Cruz Biotechnology).
Immunohistochemistry. Surgically removed tumors were fixed in 4% formaldehyde/PBS (pH 7.2). After dehydration in a graded series of 2-propanol solutions, tissues were embedded in paraffin. Serial 7 μm-thick tissue sections were deparaffinized in Roti®-Histol (Carl Roth, Karlsruhe, Germany) and endogen peroxidase activity was blocked by incubation of tissue slices in a 3% H2O2 methanol solution for 30 min followed by rehydration in serially diluted 2-propanol solutions. To preserve optimal antigen retrieval, sections were incubated for 15 min at 95°C in 10 mM sodium citrate buffer, pH 6.0. After cooling for 30 min, slices were blocked for 30 min with 10% normal goat serum (Dako) in PBS prior to incubation with antibodies to ERBB2, ERBB3 or ERK1/2, and 5% bovine serum albumin/PBS prior exposure to cetuximab for EGFR, then washed and incubated with the respective primary antibodies overnight at 4°C, followed by incubation at 37°C for 1 h the next day. Subsequently, slices were washed three times for 5 min in PBS followed by detection of cetuximab binding which was achieved by incubation with a biotinylated secondary anti-human IgG antibody and treatment with avidin/biotin/peroxidase complex (ABC Elite). Detection of anti-ERBB2, anti-ERBB3 and anti-ERK1/2 binding was achieved with the EnVision+ System HRP-labeled polymer (Dako, Glostrup, Denmark). Chromogen signals (brown precipitates) were generated by peroxidase activity after incubation with (3,3’-diaminobenzidine. Tissue slices were counterstained with Mayer's hemalaun solution (Merck). No non-specific reactivity was detected when using corresponding normal IgG (Dako) instead of the primary antibody. Staining results were documented qualitatively with an Olympus Provis AX70 bright field microscope (Olympus Optical, Tokyo, Japan).
Sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and western blot analysis. To evaluate protein expression levels, SDS-PAGE (10%) and western blot analysis were performed with the antibodies indicated above under standard laboratory conditions as described elsewhere (21).
Statistical analysis. GraphPad Prism Software 6.0 (GraphPad Software Inc., San Diego, CA, USA) was used for statistical evaluation. The two-tailed, unpaired Student's t-test was implemented to evaluate differences between the O3/O2PP-and sham-treated groups. A value of p<0.05 was considered to represent significant differences between groups. Welch's correction was applied whenever variances of the group means differed significantly.
Results
Progressive growing of VX2 carcinomas were compared with regressing tumors. One feature of the VX2 animal model is that spontaneous remission of the tumor is observed in up to 25% of cases, pointing to the inherent potential of the NZW rabbit to reject the VX2 carcinoma. The remission rate of auricular VX2 tumor can be increased to 70% when applying O3/O2-PP (18). Here, the O3/O2-PP treatment was used to generate a high number of regressing tumors. Two regressive and two progressive tumors were used as a first approach to compare gene-expression profiles between the two groups, which should indicate possible candidate genes associated with tumor remission.
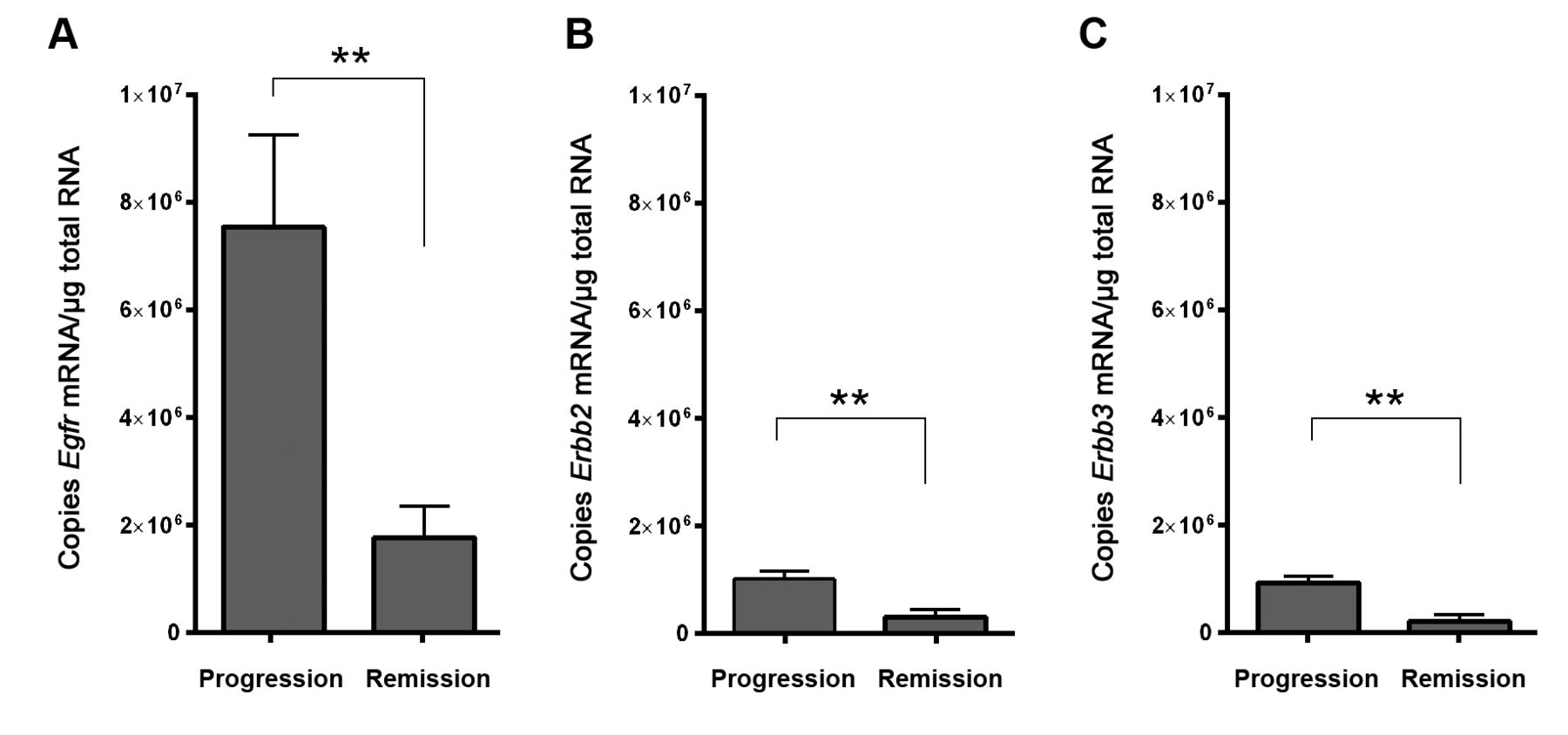
Gene-expression profiles of progressive compared with regressive VX2 tumors. Figure 1A and B depict the macroscopic view and growth course of two progressive and two regressive VX2 tumors, respectively. Total mRNA derived from these tumors was used for microarray analysis leading to the detection of 540 distinctive genes (Figure 1C). More detailed analysis of this gene profile focusing on vascular-associated genes pinpointed six candidate genes that significantly differed in their expression levels between the two groups (Figure 1D). It was remarkable to note that only the level of expression of Egfr mRNA demonstrated major (30-fold) down-regulation in regressive tumors, which is in line with the concept that EGFR, as well as other RTKs, promote tumor growth and development.
Quantification of mRNA for ERBB receptors in VX2 tumor tissues. Observation of the major down-regulation of Egfr mRNA in regressive VX2 tumor tissues prompted us to evaluate the expression of all ERBB RTK family members. qRT-PCR was deployed to confirm down-regulation of Egfr in regressive VX2 tumor tissues and to analyze the expression pattern of other ERBB receptor family members. To analyze independent samples, we used tumor tissues from an independent VX2 animal study in which O3/O2-PP was also deployed to generate tumor remission. However, in that study, the criteria for tumor remission were even more stringent (reduction to <25% of the tumor's maximal size) than in the present animal study.
Absolute quantification of Egf mRNA confirmed the significant down-regulation of the receptor (Figure 2A) as observed from the microarray analysis. Similarly, although generally having lower expression than Egfr, Erbb2 and Erbb3 mRNAs were also significantly down-regulated in regressive tumors (Figure 2B and C). In our experimental setting, no Erbb4 mRNA was detected in any of the tumors.

Growth kinetics and gene profiles of progressive and regressive VX2 tumors. A: Macroscopic view of VX2 tumors in progression or remission (day 10 after treatment). B: Development of tumor volume in sham-(sham A and sham B) and O3/O2-PP-treated (T 91 and T 92) animals. C: Hierarchical clustering of 540 selected genes after microarray analysis of VX2 tumor samples. Each column represents one tumor tissue sample and each row refers to one gene. Relative gene-expression levels are depicted according to a color scale in which blue represents the lowest and red the highest level of expression. D: Gene-profile analysis focusing on vascular-associated genes revealed six differentially regulated candidate genes. S1pr1: Sphingosine 1-phosphate receptor 1 (NM_001198946); Ednra: endothelin receptor type A (NM_001105674); Egfr: epidermal growth factor receptor (AF243516); Enos3: endothelial nitric oxide synthase (NM_001082733); Sell: selectin L, (NM_001082352); Vcam1: vascular cell adhesion molecule (NM_001082152).
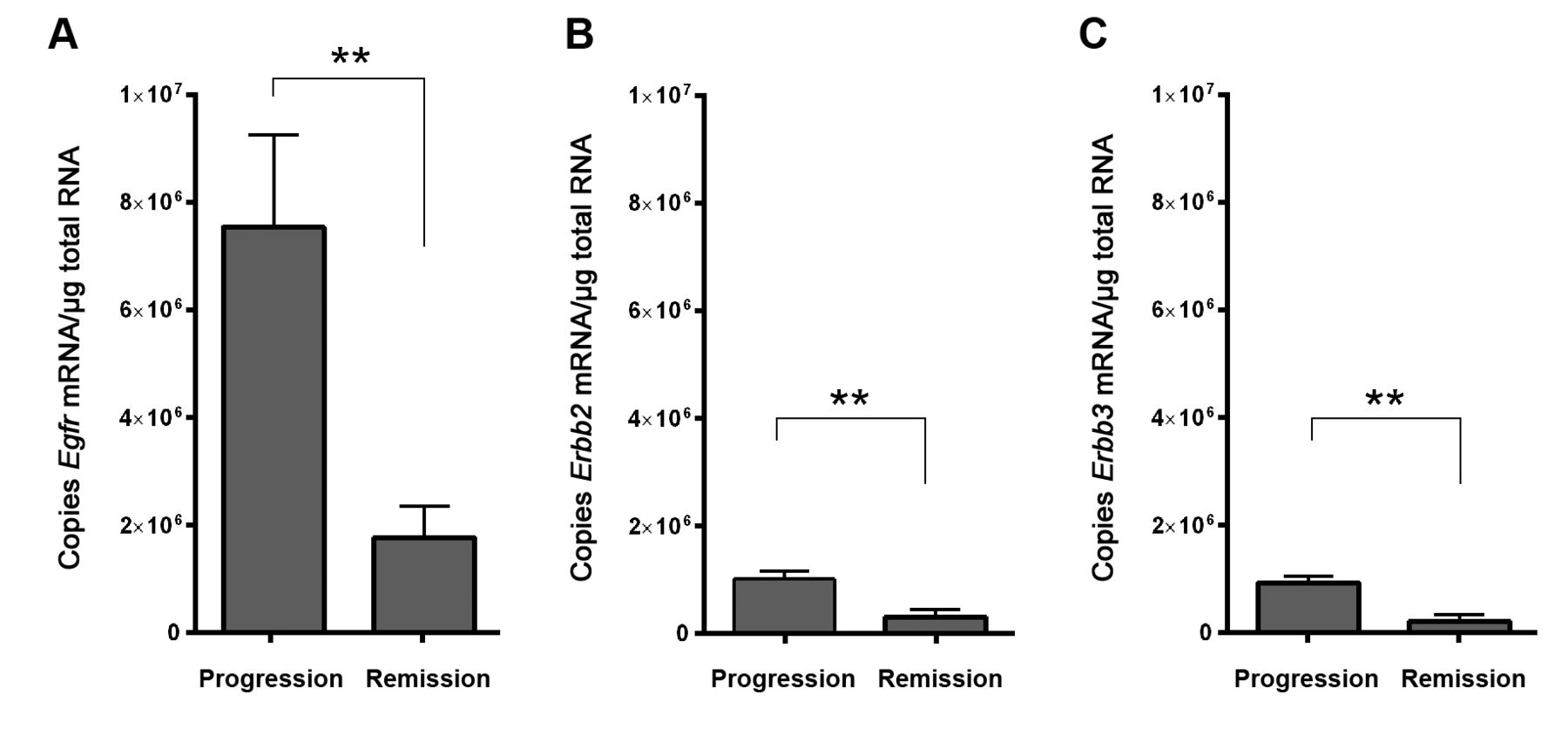
Expression of erythro-blastic leukemia viral oncogene (Erbb) family members in VX2 tumor tissues. Graphs depict the absolute number of A: Egfr, B: Erbb2 and C: Erbb3 mRNA copies per μg total RNA in O3/O2-PP-treated progressive (n=6) or regressive (n=7) tumors. Data are the mean±SEM. **Significantly different at p<0.005.

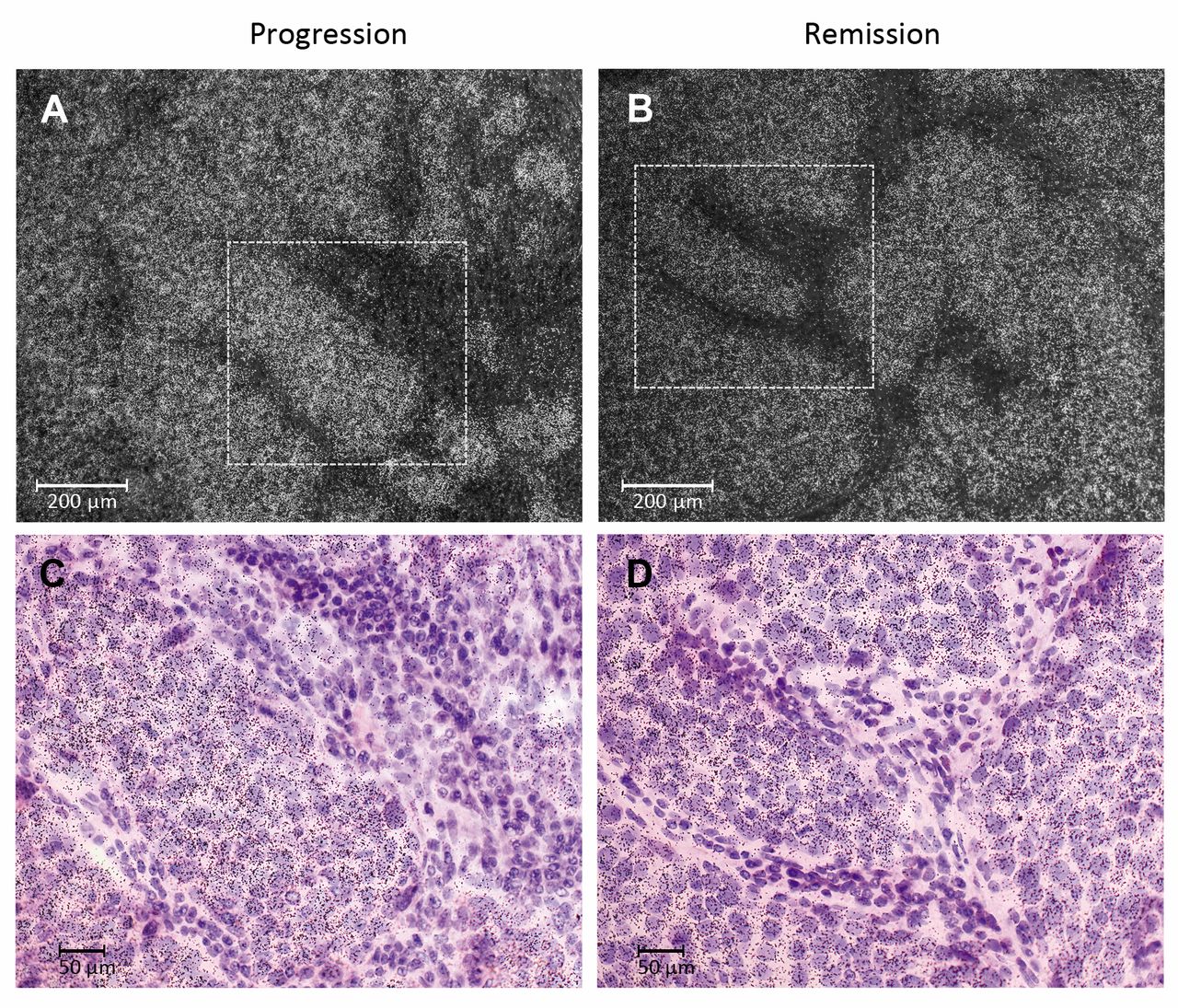
Epidermal growth factor receptor (Egfr) mRNA expression in auricular VX2 tumor tissue. A: Histological overview of an implanted VX2 carcinoma of the rabbit auricle. The epidermis of the ventral skin is shown at the bottom of the image. B: and C: Dark-field microscopy of radioactive-labelled Egfr mRNA (white dots) in tumor cells from progressive and regressive tumors, respectively. D and E: Documentation using bright-field analysis of Egfr mRNA in VX2 tumoral, but not stromal cells from progressive and regressive tumors, respectively. Areas in B and C (dotted boxes) are shown at higher magnification in D and E.
ISH of VX2 tumor tissues using Erbb-specific probes. In order to evaluate which cell types in the tumor express the Erbb mRNAs detected by microarray and qRT-PCR, we generated rabbit Erbb DNA probes for ISH. For PCR cloning, RNA isolated from the original VX2 tumor tissues was deployed. Using the primer pairs listed in Table I we were able to generate rabbit Egfr, Erbb2 and Erbb3 but not Erbb4 DNA fragments. Sequence analysis of the cloned DNA fragments revealed high identity to the published rabbit (rb) reference sequences (rbEgfr clone #4: 99.7% to XM_008261844.2; rbErbb2 clone #8; 99.0% to XM_002719343.3; rbErbb3 clone #3; 100% to XM_002711067.1. We were unable to isolate rbErbb4 DNA from VX2 tumor tissues in our experimental settings.
It was found that Erbb receptor mRNAs were predominately expressed by the epithelial portion of the VX2 tumor (Figures 3, 4 and 5), which identifies these cells as the major source of the Erbb mRNA measured. In addition, robust expression of Erbb2 mRNA was also seen in normal epithelial cells of the epidermis and sebaceous glands (Figure 4). Similarly, these structures also expressed Egfr and Erbb2 mRNA, although at lower levels (data not shown).
Erythroblastic leukemia viral oncogene 3 (Erbb3) mRNA expression in VX2 tumor cells. A and B: Dark-field analysis; C and D: bright-field detection of areas highlighted in A and B at higher magnification.
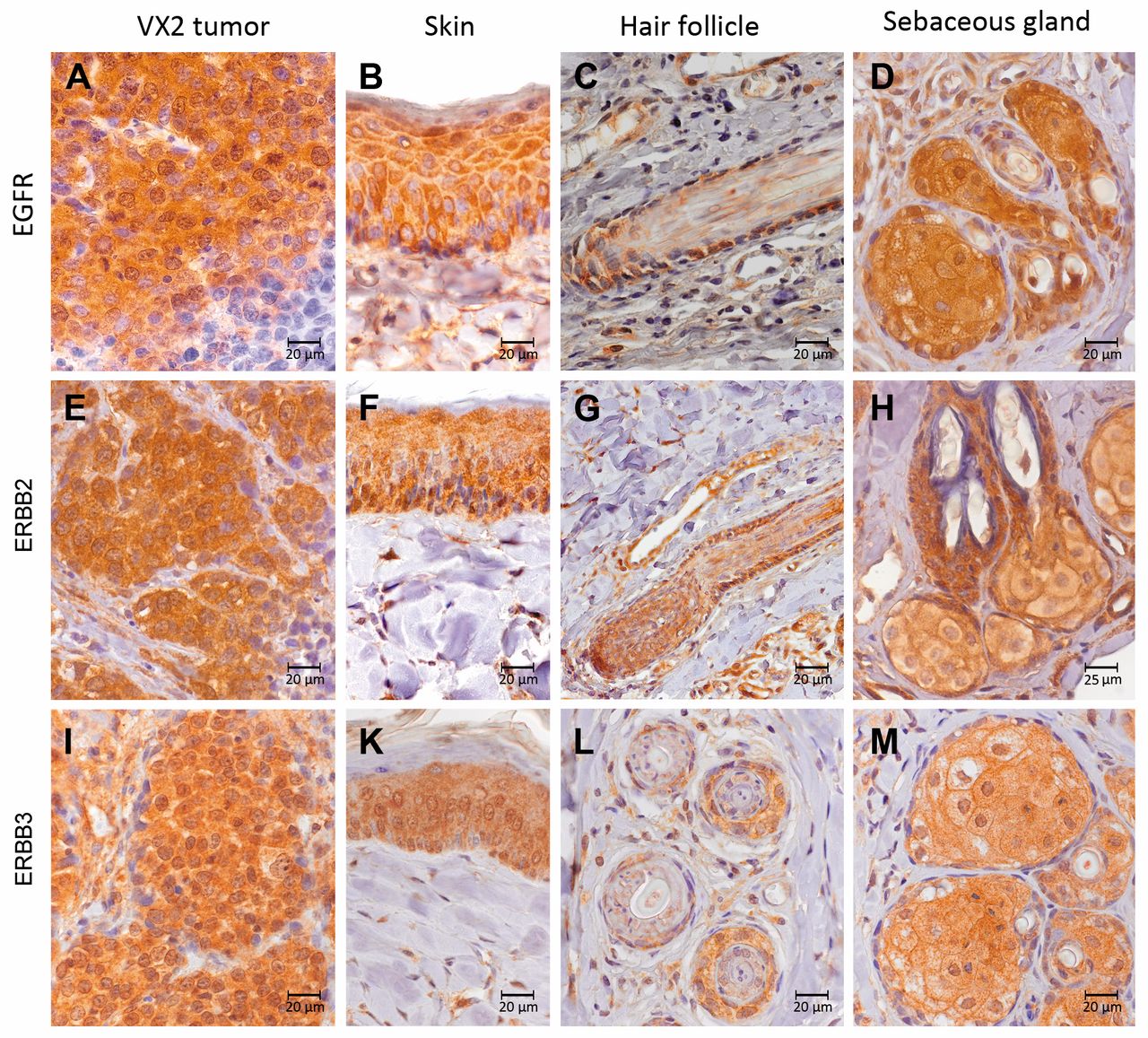
Immunohistochemical analysis of FFPE tissue slices was consistent with the ISH data revealing protein expression of EGFR, ERBB2, and ERBB3 in VX2 tumor cells, as well as in epidermal cells of the skin, outer cells of the hair follicle and in sebaceous glands (Figure 6).
ERBB downstream signaling in VX2 tumors. In order to assess if ERBB signaling is functional in VX2 tumor cells, we evaluated expression of major ERBB downstream signaling intermediates, such as MAPK2K1 as well as the presence of active ERK1/2. Active ERK1/2 was detected by western blot analysis and immunohistochemistry. Consistent with its being in its active form, ERK1/2 exhibited major nuclear localization and was also detected in normal epithelial cells such as cells of the epidermis, hair follicles and sebaceous glands (Figure 7) that also expressed EGFR, ERBB2 and ERBB3. Similarly, Mapk2k1 transcript levels were detected in progressive and regressive tumors at similar expression levels by qRT-PCR analysis.
Discussion
RTKs of the ERBB family are essential for the regulation of numerous biological outcomes such as cell proliferation, differentiation, adhesion and migration. EGFR (ERBB1), the most prominent representative of the RTK family, is particularly responsible for the maintenance of epithelial cell growth (22). Mutation or overexpression of ERBB receptors are often associated with the development and progression of human malignancies in which distinct ERBB RTK members exhibit a decisive role depending on the tumor entity (3, 23-25). Mutation or overexpression of EGFR (e.g. by gene amplification) has been implicated in the carcinogenesis and disease progression of more than 90% of all HNSCC cases (26-28). Furthermore, several studies demonstrated EGFR to be associated with an increased tumor size, risk of recurrence, and decreased therapeutic sensitivity (29-33). However, less attention has been paid to its role as a diagnostic marker.

Erythroblastic leukemia viral oncogene 2 (Erbb2) expression in rabbit skin and VX2 tumor tissues. Detection of Erbb2 mRNA in the epidermis (A, B) and sebaceous glands (C, D) by dark-field (A, C) and bright-field (B, D-F) microscopy. Erbb2 mRNA was expressed in VX2 cells of progressive (E) and regressive (F) tumors.

Protein expression of epidermal growth factor receptor (EGFR) and erythroblastic leukemia viral oncogene (ERBB) receptors. A-D: EGFR; E-H: ERBB2 and I-M: ERBB3 were expressed in VX2 tumor cells, skin, hair follicle and sebaceous glands.
Here we present data of ERBB receptor expression in an experimental animal tumor (VX2 carcinoma of the NZW rabbit) which serves as a model system for human HNSCC. In previous studies, we reported that application of O3/O2-PP is an effective approach to initiate processes leading to remission of solid auricular VX2 squamous cell carcinomas in the NZW rabbit animal model (18, 19). Deploying the O3/O2-PP treatment we were able to induce VX2 tumor remission. To identify possible candidate genes that may contribute to such remission, here the mRNA expression profile of progressive and regressive tumors was evaluated by microarray analysis. Amongst other candidate genes, we identified Egfr as a major differentially regulated gene. Egfr mRNA expression level dropped by a factor of 30 in regressive tumor tissues compared to tumors in progression.
Further evaluation of Egfr mRNA expression levels by qPCR confirmed its significant down-regulation in regressive tumors. This implies that progressively growing VX2 tumors exhibit significantly higher Egfr mRNA levels, which is in line with the observed overexpression of EGFR in up to 90% of human HNSCC (26). In this context, elevated EGFR copy numbers were found to be associated with reduced survival in patients with HNSCC (34). However, other, studies failed to prove a correlation of altered EGFR mRNA or protein levels and clinical outcome (7).
The role of ERBB2 in HNSCC is less understood, but studies have demonstrated an increased ERBB2 expression in a large percentage of patients with oral HNSCCs (35-37). Western blot and immunohistochemical analyses highlight variable ERBB2 expression which, however, appears to be limited to a subset of HNSCC tumors and cell lines. Expression of ERBB2 was found to be highly regulated at the transcriptional or post-transcriptional level (35). Published data of ERBB2 overexpression and its correlation with HNSCC disease progression and survival are controversial (38, 39) and its role as a reliable marker for HNSCC is incompletely understood. This in part appears to be due to the lack of a ligand-binding domain in ERBB2 and its function rather as an amplifier of ERBB receptor activation (40), thereby masking its possible impact on HNSCC. Upon ligand binding, monomeric ERBB receptors are activated through homo-or heterodimerization with other members of the ERBB family (41). ERBB2 is unique since it does not homodimerize under normal conditions and its ectodomain does not bind ligands (42, 43). There is evidence that ERBB2 is the preferred dimerization partner for all other members of the ERBB receptor family after ligand binding (42, 44, 45), which has led to the suggestion of it having a key function in ERBB RTK activation (43).

Expression of erythroblastic leukemia viral oncogene (ERBB) downstream signaling intermediates. A: Comparison of mitogen-activated protein kinase kinase 1 (MAPK2K1) expression in tumor tissues. B: Western blot analysis of active (phosphorylated) extracellular signal-regulated kinase 1/2 (ERK1/2) in homogenates of different tumors in progression (lanes 1-5) and remission (lanes 6-11). Arrowheads point to phospho-ERK1/2-specific bands. Immunohistochemical detection of active ERK1/2 in C: VX2 tumor, D: epidermis, E: hair follicles and F: sebaceous gland. Note the predominant nuclear localization of active ERK1/2.
Especially when overexpressed, ERBB2 can maintain tumorigenesis caused by other ERBB receptor family members. Many tumors which express a high level of ERBB2 also express ERBB3 (12), suggesting that ERBB2–ERBB3 heterodimers are required for ERBB-mediated signaling in tumor cells. Indeed, the existence of ERBB2–ERBB3 heterodimers forming functional oncogenic units that drive tumor cell proliferation have been experimentally verified by Holbro and colleagues using different breast cancer cell lines (46).
In VX2 rabbit tumor, we observed expression of EGFR, ERBB2 and ERBB3. ERBB4 was not detected in VX2 carcinoma during our analyses. VX2 cells thereby carry the potential to form heterodimers between these three ERBB representatives. Detection of Mapk2k1 mRNA in VX2 tumor tissues, as well as immunohistochemical detection of activated ERK1/2, points to an active rat sarcoma (RAS) – rapidly accelerated fibrosarcoma (RAF)–MAPK2K1–ERK1/2 signaling pathway. However, we did not detect obvious changes in the activation status of this pathway in progressive and regressive tumors. In human cancer, this pathway is frequently up-regulated and over activated, thereby serving as an attractive target for anticancer drugs (47).
This study highlights changes in the Erbb gene signature of rabbit VX2 carcinomas as a possible predictor for therapy response. Further studies will have to evaluate if and how this information is applicable to diagnostic, prognostic and therapeutic procedures in the human patient.
Acknowledgements
The Authors thank Mrs. R. Peldszus and G. Sadowski for preparation of the VX2 tumor cell suspension, and Mrs. M. Zibuschka and H. Hlawaty for assistance with the histological analysis. This research was funded in part by the Wirtschafts-und Infrastrukturbank Hessen (TM MBF-P_04/2009).
- Received June 22, 2018.
- Revision received July 18, 2018.
- Accepted July 19, 2018.
- Copyright© 2018, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.