


Abstract
Liposomes incorporating stearic acid-modified octa-arginine (StA-R8) and conjugated to transferrin (Tf) were synthesized and evaluated for delivery of small interfering RNA (siRNA) against survivin. Characteristics, cytotoxicity, cellular uptake and biological activity of liposomes complexed to survivin siRNA were investigated. Tf-conjugated liposomes were shown to have reduced cytotoxicity and improved delivery efficiency for siRNA into cancer cells compared to non-conjugated StA-R8 liposomes. These data suggest that Tf-conjugated StA-R8 liposomes are promising vehicles for siRNA delivery in cancer therapy.
Cell-penetrating peptides (CPPs) are able to be internalized by mammalian, plant and bacterial cells and to facilitate delivery of molecules with biological activity across the cell membrane (1-3). Many CPPs have been reported, including HIV-Tat (4), penetratin (5), transportan (6) and octa-arginine (R8) (7). R8, is highly cationic and can interact with the plasma membrane and facilitate membrane translocation (8).
Non-viral vectors can be used for delivery of small interfering RNA (siRNA) (9, 10). These include liposomes (11, 12), polymers (13), and polypeptides (14). Liposomes have difficulty crossing the cellular membrane, which greatly limits their applications in vitro and in vivo. Incorporation of CPPs into liposomes is a strategy to improve membrane translocation (15-17).
The transferrin (Tf) receptor is a glycoprotein expressed on the surface of many tumor cells (18). Tf receptor mediates endocytosis of Tf and Tf conjugates (19). Tf conjugation has been used in targeted delivery to tissues including tumor, endothelial cells and the brain (20).
In this article, we report the synthesis and evaluation of Tf-conjugated liposomes containing stearic acid-modified R8 (TSLP) for delivery of survivin siRNA into cancer cells.
Materials and Methods
Materials. 2-Chlorotrityl chloride resins and fluorenylmethyloxycarbonyl-L-arginine (Fmoc-Arg) were obtained from JiEr Biochemical Co. (Shanghai, China). Stearic acid (StA; 98.5%), cholesterol, holo-human Tf, 2-iminothiolane hydrochloride (Traut's reagent) and 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) were purchased from Sigma-Aldrich (St. Louis, MO, USA). Egg phosphatidylcholine (ePC) was purchased from Avanti Polar Lipids (Alabaster, AL, USA). Distearylphosphatidyle-thanolamine-polyethyleneglycol (M.W. 2,000)-maleimide (DSPE-PEG2000-Mal) was purchased from Shanghai Advanced Vehicle Technology Pharmaceutical Ltd. Co. (Shanghai, China). Fetal bovine serum (FBS), Dulbecco's modified Eagle's medium (DMEM) and 0.25% (w/v) trypsin were from HyClone (Logan, UT, USA). Anti-survivin siRNA, and 6-carboxyfluorescein (FAM)-labeled siRNA were synthesized by Ribo Biochemistry (Guangzhou, China). 4’,6-Diamidino-2-phenylindole (DAPI) was purchased from Invitrogen Molecular Probes (Eugene, OR, USA). All other reagents were commercially purchased in reagent-grade.
Synthesis of StA-R8 and Tf-conjugated PEG-DSPE (Tf-PEG-DSPE). StA-R8 was synthesized by the technique of Fmoc solid-phase peptide synthesis on a 2-chlorotrityl chloride resin as reported previously (21, 22). The target peptide was then purified and structurally confirmed. Tf-PEG-DSPE was synthesized using a method modified from a previously reported method (23). Briefly, Tf and Traut's reagent were combined at a molar ratio of 1:10 and incubated for 1 h in dark, and then dialyzed for 2 h to generate thiolated Tf. Next, thiolated Tf and DSPE-PEG2000-Mal were combined at a molar ratio of 1:10 and incubated overnight in the dark (24).
Preparation of liposomes. An ethanol dilution method was used for the preparation of liposomes. First, StA-R8, ePC and cholesterol were dissolved in ethyl alcohol at a molar ratio of 20:35:45. This solution was then injected into HEPES-buffered saline (pH 7.4) under vortexing at a lipid/HEPES volume ratio of 1:3 to generate StA-R8 liposomes (SLPs). The liposomes were then sonicated for 30 s. SLP/siRNA was synthesized by combining SLPs with anti-survivin siRNA and incubating for 20 min at room temperature. A post-insertion method was used to prepare TSLPs. For Tf post-insertion, SLP/siRNA were incubated with Tf-PEG-DSPE at 37°C for 30 min (25).
Agarose gel retardation assay. SLPs were complexed with siRNA at different N/P ratios (with “N” representing positively charged guanidinium group in StA-R8 component of SLP and “P” representing negatively charged phosphates in siRNA) from 0.5:1 to 8:1. TSLP/siRNA complexes were prepared at an N/P ratio of 6:1. Formation of these complexes was confirmed by a gel retardation assay. The samples were incubated at room temperature for 30 min and then loaded onto 1% (w/v) agarose gel in Tris-acetate-EDTA buffer, containing 1 μg/ml ethidium bromide (Bioteke Corporation, Beijing, China). Electrophoresis was performed at 130 V for 12 min and the gel was visualized with a UV lamp using a Vilber Lourmat imaging system (Vilber Lourmat, Marnela Vallée, France).
Particle size and zeta potential measurements. The particle size and zeta potential of SLPs, SLP/siRNA and TSLP/siRNA were determined on a Zetasizer Nano ZS 90 (Malvern Instruments, Ltd., Malvern, UK). The measurements were repeated three times and averaged. SLP/siRNA and TSLP/siRNA samples were stored at 4°C. To evaluate their stability at this temperature, particle sizes on days 0, 1, 2, 4, 6, 8, 10 and 12 after synthesis were determined.
Cell culture. HeLa and HepG2 cells were cultured in DMEM with 10% FBS and 1% antibiotics/antimycotics in a humidified atmosphere containing 5% CO2 at 37°C.
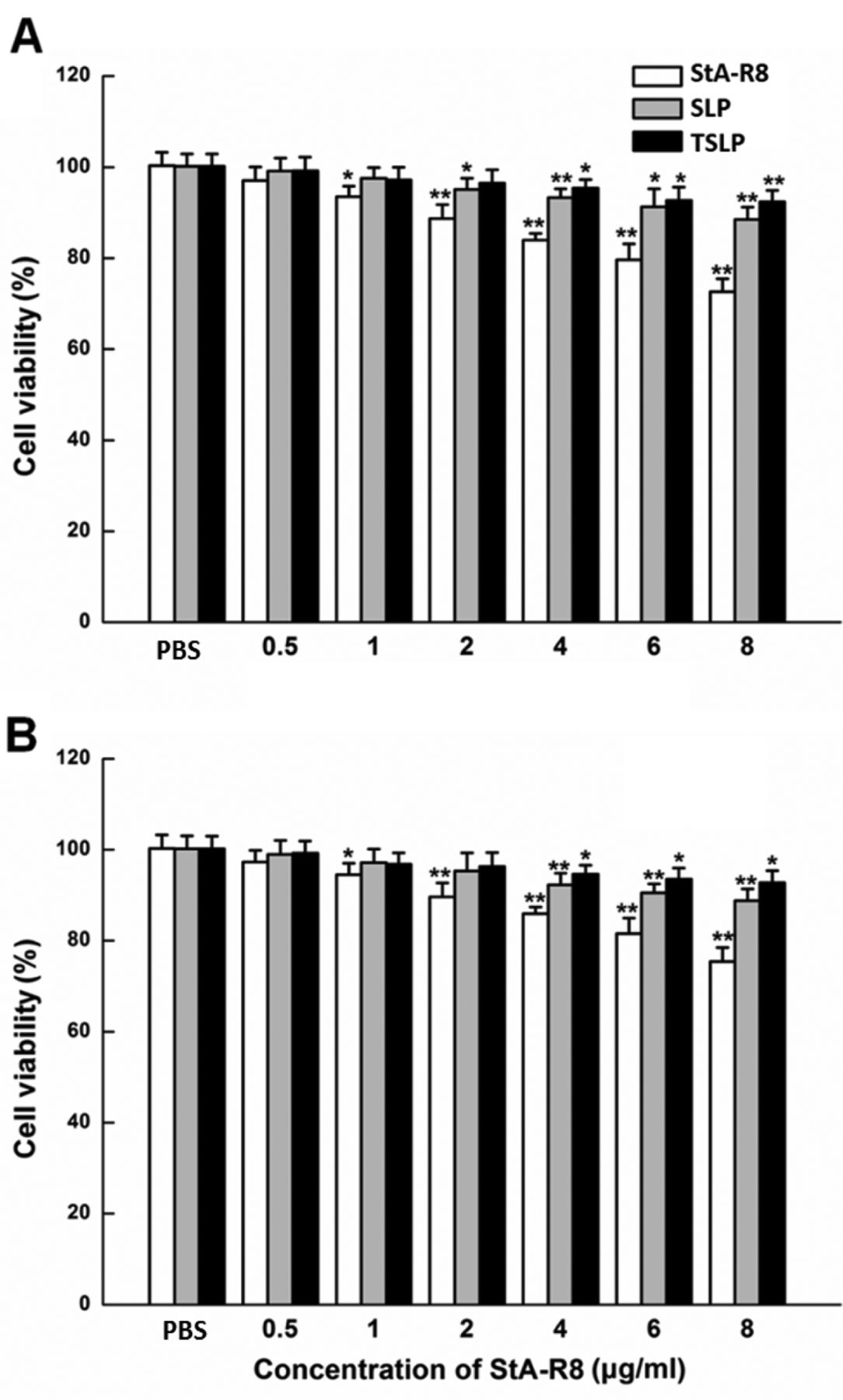
Cytotoxicity assay. Cell viability was measured by MTT assay. HeLa and HepG2 cells were seeded in a 96-well plate (1×104 cells/well) and cultured for 24 h. Subsequently, StA-R8, SLPs and TSLP at different concentrations were added to the plate and the cells were incubated for 4 h. Then, serum-free medium was replaced with fresh medium containing FBS. After incubating for another 20 h, 10 μl of MTT stock solution (5 mg/ml) was added to each well and the plate was incubated for 4 h at 37°C. Finally, 100 μl/well DMSO was added to dissolve the formazan crystals formed due to MTT reduction by viable cells. The absorbance was then measured at 490 nm on a Synergy 4 microplate reader from BioTek Instrument, Inc. (Winooski, VT, USA).
Cellular uptake assay. Cells were seeded in a 24-well plate at 1×105/well. SLP/siRNAFAM or TSLP/siRNAFAM diluted in 1 ml of serum-free medium was added to cells, which were then incubated for 4 h in the dark at 37°C. Then, the cells were washed three times with PBS, harvested and fixed in 4% para-formaldehyde. Samples were analyzed on a flow cytometer from BD Biosciences (San Jose, CA, USA).
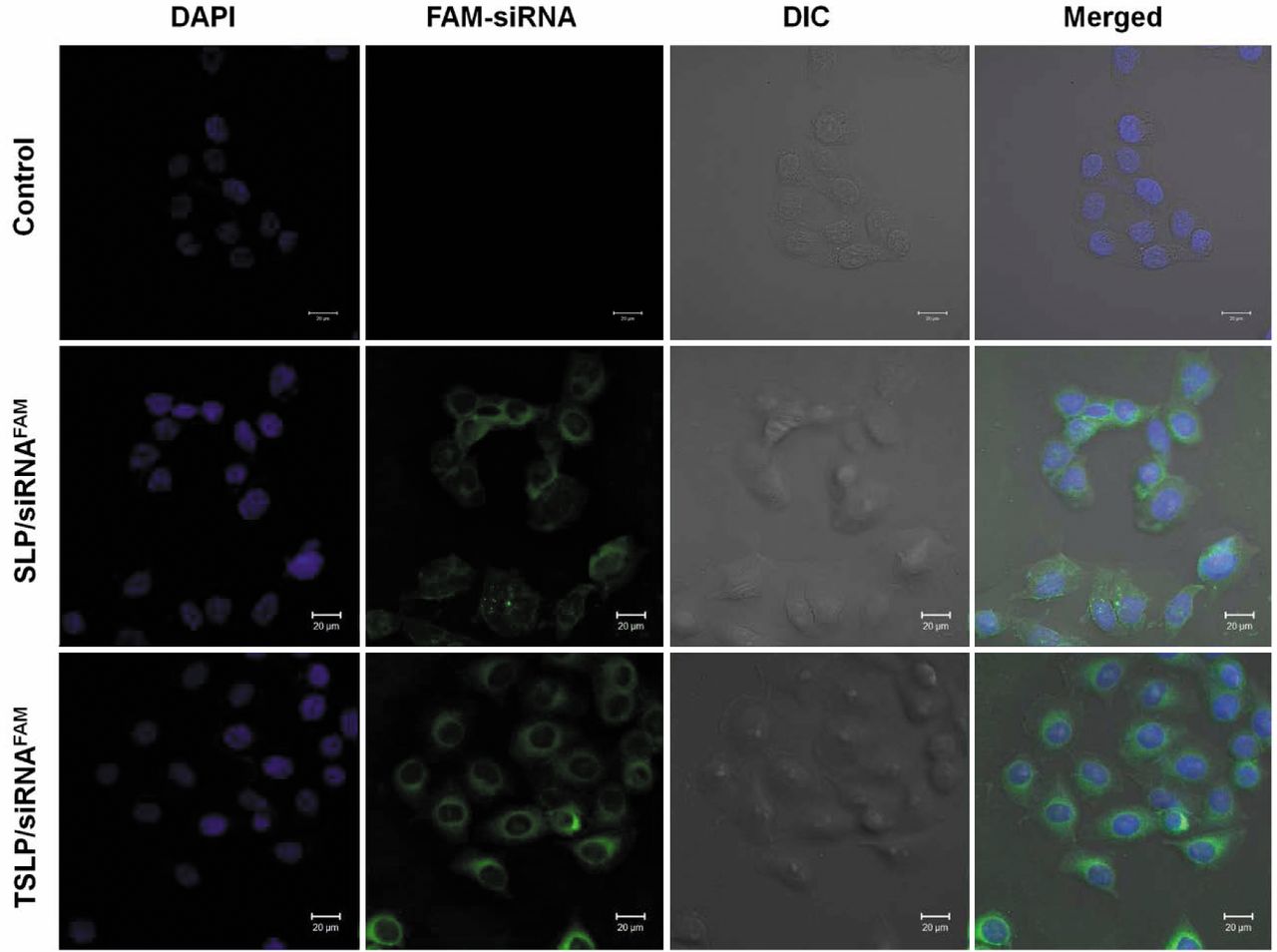
Confocal microscopy. Cells were seeded in a 35-mm glass bottom culture dish at 1.5×105/well. SLP/siRNAFAM or TSLP/siRNAFAM diluted in 1 ml of serum-free medium was added to the cells, which were incubated for 4 h in dark at 37°C. Then, the cells were washed three times with PBS and fixed with 4% paraformaldehyde/PBS for 20 min. Cellular nuclei were counterstained with DAPI (2 μg/ml) for 3 min at room temperature and washed with PBS. The cells were then observed on a Zeiss 710 LSMNLO Confocal Microscope from Carl Zeiss (Jena, Germany).

Agarose gel analysis of liposome/siRNA complexes. siRNA complexes were prepared at different N/P ratios. Lane 1, naked siRNA; Lane 2-7: stearic acid-modified octa-arginine (StA-R8)-containing liposome/siRNA complexes at different N/P ratios, 0.5:1 (lane 2), 1:1 (lane 3), 2:1 (lane 4), 4:1 (lane 5), 6:1 (lane 6) and 8:1 (lane 7); 8, transferrin-conjugated liposomes containing StA-R8/siRNA complexes at an N/P ratio of 6:1.
Measurement of survivin protein expression by Western blot. HeLa cells were seeded in a 6-well plate at 2×105 cells/well. SLP/siRNA or TSLP/siRNA diluted in 1 ml of serum-free medium was added and the plate was incubated for 4 h at 37°C. The serum-free medium was then replaced with fresh medium containing 10% FBS. The cells were cultured for another 44 h, washed and harvested. Western blot was used to measure survivin protein expression.
Results
Formation of SLP/siRNA complexes. In order to evaluate the complexing activity of various liposomes to survivin siRNA, agarose gel electrophoresis was performed. Free survivin siRNA showed a bright band on the gel, which is absent when siRNA was complexed to liposomes. As shown in Figure 1, the first lane is naked siRNA. With increase of charge (N/P) ratio, the brightness of the band was reduced. When the N/P ratio was above 6:1, the band almost disappeared, indicating complete complexation of siRNA by the liposomes. The eighth lane in Figure 1 shows that TSLP/siRNA had a good complexation efficiency when the N/P ratio was 6:1 and that insertion of Tf had no significant effect on complexation efficiency.
Characterization of liposomes. Liposomes were prepared as described above. Particle size and ζ potential of SLPs, SLP/siRNA and TSLP/siRNA are shown in Table I. After complexing with siRNA, particle size of SLPs increased and the ζ-potential decreased. Compared to SLP/siRNA, particle size and ζ-potential for TSLP/siRNA changed, which was due to the presence of negatively charged Tf. TSLP had good colloidal stability at 4°C, and the particle size remained below 200 nm (Figure 2).

Stability of by particle size measurement.Transferrin-conjugated liposomes containing stearic acid-modified octa-arginine complexed with siRNA were synthesized as described in the Materials and Methods and stored at 4°C. Particle size was measured by dynamic light scattering at different time points.
Size and ζ-potential of stearic acid-modified octa-arginine (StA-R8)-containing liposomes (SLP), SLP/siRNA and transferrin-conjugated liposomes containing StA-R8 (TSLP)/siRNA at pH 7.4. Data are shown as the mean and standard deviation (n=3).
Assessment of cytotoxicity. The cytotoxicity of SLPs and TSLP was determined in HeLa and HepG2 cells. Figure 3 shows the effects of StA-R8, SLP and TSLP on cell viability. As shown, at all concentrations of liposomes studied, cell viability in both cell lines was over 85% compared with the control (0 μg/ml). Free StA-R8 showed higher cytotoxicity than the corresponding liposomes. Thus, SLP and TSLP displayed reduced cytotoxicity as a vector. There was no significant difference in cytotoxicity between SLP and TSLP.
Study of FAM-siRNA uptake by flow cytometry. Flow cytometry was used to quantify the uptake of SLP/siRNA and TSLP/siRNA by HeLa and HepG2 cells (Figure 4). The fluorescence intensity peaks of cells after treatment with FAM-siRNA liposomes migrated to the right (Figure 4 A and B). This showed that, compared to naked siRNA, the uptake of SLP/siRNA and TSLP/siRNA by HeLa and HepG2 cells were much higher. The mean fluorescence intensities of the treated cells are shown in Figure 4 C and D. In HeLa cells, the fluorescence intensity values for cells treated with TSLP/siRNA complexes were about 10 times those with naked siRNA. In HepG2 cells, TSLP/siRNA complexes also showed higher cellular uptake. Meanwhile, the uptake efficiency of TSLP/siRNA was higher than that of SLP/siRNA. These data indicate that Tf in TSLP mediated more efficient in vitro uptake of siRNA.

Cytotoxicity of different agents. HeLa (A) and HepG2 (B) cells were treated with PBS (vehicle control), StA-R8, SLP, or TSLP and assessed for viability by the MTT assay, as described in Materials and Methods. *p<0.05; **p<0.01; ***p<0.001 compared with untreated control.

Cellular fluorescence uptake in vitro by flow cytometry. HeLa (A) and HepG2 (B) cells were treated with 6-carboxyfluorescein-labeled siRNA complexed to liposomes and then analyzed by flow cytometry. Mean fluorescence values of HeLa (C) and HepG2 (D) cells are shown. The treatments were: a: Control, b: siRNA, c: stearic acid-modified octa-arginine-containing liposomes/siRNA, d: transferrin-conjugated liposomes containing stearic acid-modified octa-arginine/siRNA.
Cellular internalization analysis by confocal microscopy. The internalization of FAM-labeled siRNA mediated by SLP and TSLP was further evaluated in HeLa cells by confocal microscopy (Figure 5). DAPI was used for labelling the nucleus. Green fluorescence of FAM-siRNA was extensively distributed in the cytosol and the nuclei. In addition, the fluorescence intensity of HeLa cells treated with TSLP/siRNA complexes was higher than those treated with SLP/siRNA complexes. Confocal microscopy analysis indicated that the association of SLP/siRNA with HeLa cells was greater than that of TSLP/siRNA.
Determination of survivin protein. The expression levels of survivin protein in HeLa cells were determined by western blot. Densitometric analysis was used to calculate the relative level of survivin expression. The SLP/siRNA and SLP/siRNA complexes significantly inhibited the expression of survivin protein in HeLa cells, indicating that these liposomes were able to deliver survivin siRNA into cells and induce RNA interference. The results showed that TSLP/siRNA complexes more efficiently reduced the survivin protein level compared to SLP/siRNA complexes (Figure 6A). Treatment of cells with SLP/siRNA and TSLP/siRNA complexes reduced the expression of survivin protein to 44.1% and 20.4% of the control, respectively. The delivery efficiency of siRNA by TSLP was significant higher than SLP in the cell lines tested (Figure 6).
Discussion
In this study, we investigated liposomes containing StA-R8 and conjugated to Tf for delivery of survivin siRNA. Tf-PEG-DSPE was inserted into StA-R8- liposomes to further improve siRNA delivery into cancer cells.
Here, we prepared two types of liposomes, SLP and TSLP. After complexing with siRNA, TSLP had increased particle size and decreased ζ-potential compared to SLP (Table I). The colloidal stability of TSLP at 4°C was excellent. At the same time, incorporation of StA-R8 into liposomes apparently reduced their cytotoxicity (Figure 3). Tf-conjugated StA-R8 liposomes increased cellular delivery of survivin siRNA in vitro, as shown by increased siRNA uptake by tumor cells (Figures 4 and 5) and greater down-regulation of survivin (Figure 6). This is likely due to active targeting of Tf receptors.
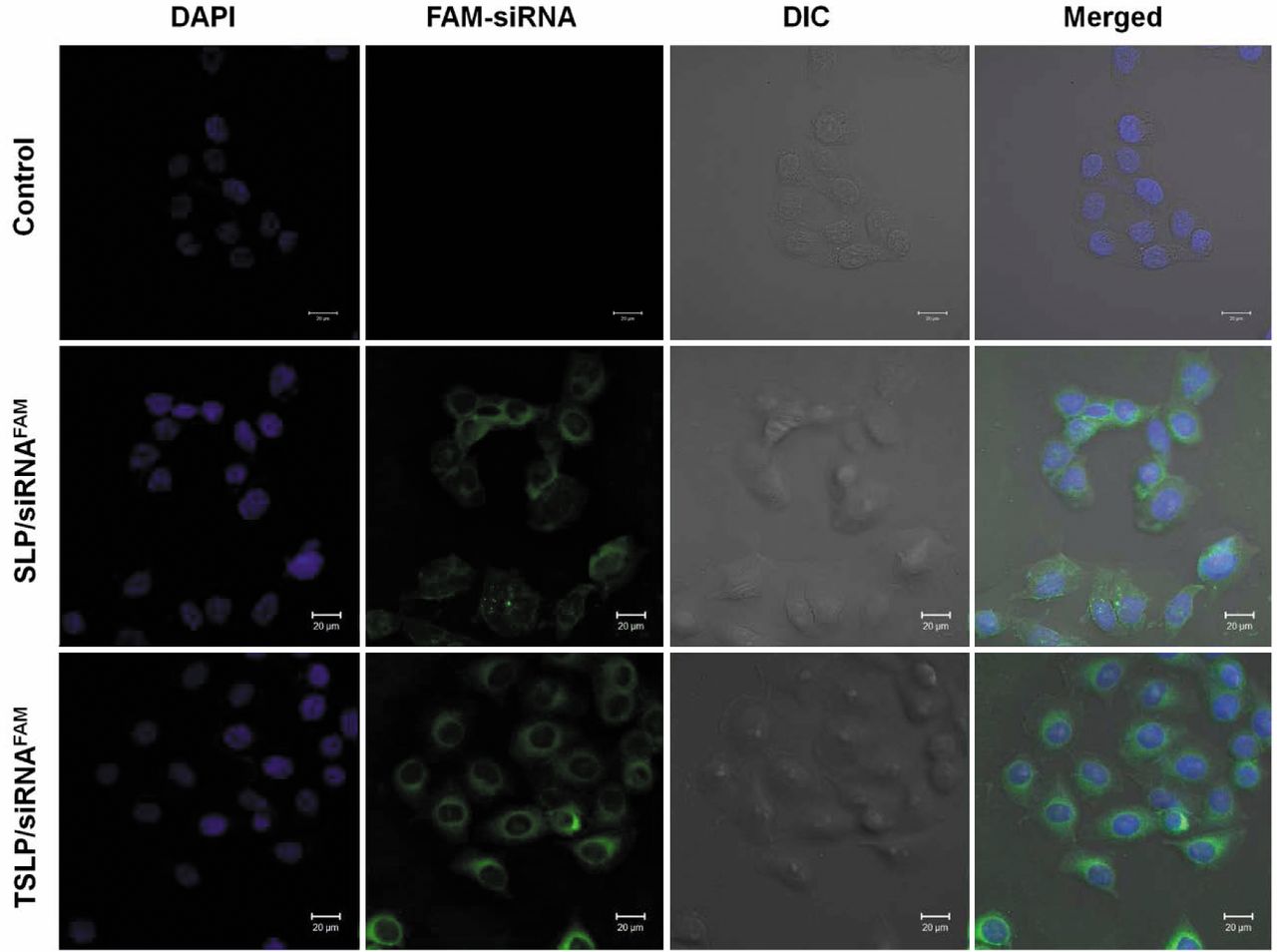
Intracellular localization of siRNA complexes by confocal microscopy. HeLa cells were treated with stearic acid-modified octa-arginine-containing liposomes (SLPs)/siRNA or transferrin-conjugated liposomes containing stearic acid-modified octa-arginine (TSLP)/siRNA complexes, stained and visualized, as described in the Materials and Methods section. Blue color: 4’,6-Diamidino-2-phenylindole (DAPI)-stained nuclei, green color: 6-carboxyfluorescein (FAM)-labeled siRNA. DIC: differential interference (phase) contrast bright-field image.

Effect on siRNA on survivin protein expression in HeLa cells. Survivin expression is shown by western blot (A) and by densitometric analysis (B). Lane 1, control; lane 2, siRNA; lane 3, stearic acid-modified octa-arginine (StA-R8)-containing liposome/siRNA complexes; lane 4, transferrin-conjugated liposomes containing StA-R8/siRNA complexes. Data are the mean and standard deviation of three experiments. **p<0.01, ***p<0.001 vs. control.
TSLP has reduced cytotoxicity and improved delivery efficiency for siRNA, and therefore warrants further evaluation as a siRNA delivery vehicle.
- Received February 17, 2017.
- Revision received March 7, 2017.
- Accepted March 13, 2017.
- Copyright© 2017, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.