


Abstract
Background: Epidermal growth factor receptor (EGFR) is a target of molecular therapeutics for colorectal cancer. However, mutation of Kirsten rat sarcoma viral oncogene homolog (KRAS) gene at codons 12 and 13 attenuates the therapeutic effect of anti-EGFR therapies. Therefore, the detection of KRAS gene mutation is important for therapeutic decision-making. Materials and Methods: KRAS gene mutation at codons 12 (c.34G>T, c.35G>C, c.35G>A) and 13 (c.38G>A) in six cancer cell lines were investigated. A loop-mediated isothermal amplification-based procedure was developed that employed peptide nucleic acid to suppress amplification of the wild-type allele. Results: This mutation-oriented gene-amplification procedure can amplify the DNA fragment of the KRAS gene with codon 12 and codon 13 mutation within 30 min. Moreover, boiled cells can work as template resources. Conclusion: This newly developed procedure can be useful for patient stratification for anti-EGFR therapies.
Epidermal growth factor (EGFR)-1 induces cell proliferation, inhibition of apoptosis, angiogenesis, and invasion via Kirsten rat sarcoma viral oncogene homolog (KRAS) activation in cancer cells (1). This makes EGFR1 a target molecule for cancer therapeutics against several types of carcinoma, including colorectal adenocarcinoma, head and neck squamous cell carcinoma, and non-small cell lung carcinoma (2-6). Recently, antibodies against EGFR, and EGFR tyrosine kinase inhibitors have been used in patients with these types of cancers (2-6). Cetuximab and panitumumab are monoclonal antibodies against the extracellular domain of EGFR1 (7, 8) which inhibit the activation of EGFR; these exhibit antitumor activity when used as monotherapy or in combination with chemotherapy or radiation (9). Clinical efficacy with these molecules has been reported for colonic cancer, head and neck cancer, and non-small cell lung carcinoma (10-12). The EGFR tyrosine kinase inhibitors gefitinib and erlotinib inhibit the auto-phosphorylation of EGFR and exhibit clinical benefit in the treatment of non-small cell lung carcinoma (13, 14).
The KRAS gene mutation, however, attenuates the treatment effect of both monoclonal antibodies against EGFR (15, 16) and EGFR tyrosine kinase inhibitors (17). This mutation is the most common abnormality of the dominant oncogenes in human tumors and is a frequent occurrence in the development and progression of adenocarcinomas of the pancreas (90%), colon (50%), thyroid (50%), bladder (50%), and lung (30%) (18).
Mutations of KRAS codons 12 or 13 were detected in 37.7% of colorectal adenoma and adenocarcinoma; of these, 80.8% of the specified mutations occurred in codon 12, with 78.1% of all specified mutations occurring at the second base of either codon (19). Various assays for detecting the KRAS mutation include polymerase chain reaction (PCR) restriction fragment length polymorphism (RFLP), PCR-single strand conformation polymorphism (SSCP), PCR sequencing, and mutant–allele-specific amplification (MASA); however, all these methods require DNA purification and involve multiple steps (20-24).
The loop-mediated isothermal amplification (LAMP) assay was developed to amplify copies of a particular DNA sequence (25). The LAMP assay is a rapid and highly specific method compared to PCR. LAMP employs a DNA polymerase with high strand-displacement activity and a set of four basic primers that recognize a total of six distinct sequences on the target DNA (26). Recently, the allele-specific LAMP assay was developed to detect Janus kinase 2 (p.V617F) mutation in chronic myeloproliferative neoplasms (27). The allele-specific LAMP assay employs four basic primers, one self-annealed primer and peptide nucleic acid (PNA), which have been shown to be effective for the enrichment of mutant alleles in PCR screening (28). The PNA could not serve as a primer for polymerization because it is not recognized by the polymerase as DNA and, thus, could not be a substrate for exonuclease activities of Taq polymerase (29).
Cell lines employed in this study.
Primer sequences used in this study.
In this study, we report a novel procedure, mutation-oriented LAMP assay, based on the principle of allele-specific LAMP assay. This assay used four basic primers with a back-loop primer (30) and a PNA-based lock primer. The mutation-oriented LAMP assay enabled a rapid and specific detection of KRAS mutation at codons 12 and 13.
Materials and Methods
Ethics statement. This study was approved by the Ethics Committee of the Nihon University School of Dentistry (2009-2). Written informed consent from the donor was obtained for the use of the oral mucosal cells in this research.
Sample preparation. The DNA samples for the LAMP assay were prepared from a pancreatic adenocarcinoma cell line (MIA PaCa-2: c.34G>T, p.G12C), a myeloma cell line (RPMI 8226: c.35G>C, p.G12A), a pulmonary adenocarcinoma cell line (NCI-H1573: c.35G>C, p.G12A), a gastric carcinoma cell line (SNU-1: c.35G>A, p.G12D), a colorectal adenocarcinoma cell line (T84: c.38G>A, p.G13D), and a breast adenocarcinoma cell line (DMA-MB-231: c.38G>A, p.G13D. These cell lines have point-mutations in KRAS codons 12 or 13. For KRAS wild-type samples, the melanoma cell line (SK-MEL-28), oral squamous cell carcinoma cell line (HSC-3), non–small cell lung carcinoma cell line (NCI-H1975), colorectal adenocarcinoma cell line (HT-29), and oral mucosal cells from healthy individuals were utilized (Table I). All cell lines were obtained from the American Type Culture Collection (Manassas, VA, USA), except for HSC-3 (Riken Bio Resource Center, Ibaraki Japan) and cultured in RPMI-1640 (Gibco, Carlsbad, CA, USA) supplemented with 10% fetal bovine serum (Gibco), and maintained under standard cell culture conditions at 37°C and 5% CO2 in a humid environment. Isolated DNA and boiled cells were used as the template for amplification. Genomic DNA from each cell line was isolated using the DNeasy mini kit (Qiagen, Hilden, Germany) according to the manufacturer's instructions. Both the quality and quantity of DNA were assessed by A260/A280 ratio on a NanoDrop-1000 (Thermo Fisher Scientific Inc., Waltham, MA, USA). Moreover, crude cell lysates were used as LAMP templates. Cells were washed and suspended in phosphate-buffered saline and boiled for 1 min to obtain the crude lysate.
Primer set. A set of five primers were designed for LAMP and mutation-oriented LAMP methods: forward and backward outer primers (F3, B3), forward and backward inner primers (FIP, BIP), and back-loop primer (loop B). These primers recognize an area of KRAS, including codons 12 and 13.

Schematic diagram of loop-mediated isothermal amplification (LAMP) and mutation-oriented LAMP method. The LAMP primer set was designed to bracket the Kirsten rat sarcoma viral oncogene homolog (KRAS) codon 12 and 13 region. A lock primer was designed for the wild-type allelic sequence and tightly bound to the sequence of wild-type allele, inhibiting the LAMP reaction, with resultant amplification of the wild-type allele. B3: Backward outer primer; BIP: backward inner primer; F3: forward outer primer; FIP: forward inner primer.
PNA with exact homology to the wild-type allele of KRAS, compatible with codons 12 and 13, was used as the lock primer. In addition to the non-recognition by DNA polymerase making PNA ineligible as a primer for polymerization, the melting temperature of a fully matched PNA–DNA duplex is higher than that of DNA–DNA of the same length, and a single mismatch destabilizes the PNA–DNA hybrids, causing a melting temperature shift of 10-18°C (Figure 1) (31). Thus, the amplification of wild-type allele of KRAS gene is inhibited in the mutation-oriented LAMP method. These primer sequences are summarized in Table II.
Mutation-oriented LAMP reaction. The LAMP reaction was performed with an Isothermal Master Mix (Nippon Gene Material Co., Ltd., Tokyo, Japan), which contains DNA polymerase, thermostable pyrophosphatase, reaction buffer, dNTPs, and a double-strand DNA-binding fluorescein dye. The reaction mixture was incubated at 61°C for 35 min. The time to fluorescence positive values (Tp) were monitored on a thermally controlled fluorimeter (Genie® II; Optigene Limited, Horsham, West Sussex, UK) and the Tp values of amplification were automatically recorded. The LAMP reaction was carried out in a 25-μl reaction mixture, containing 40 pmol of each FIP and BIP, 5 pmol of each F3 and B3, specified amount of lock primer, 15 μl of Isothermal Master Mix, distilled water, and target DNA. The lock primer was excluded from the conventional LAMP reaction.
For further confirmation, the amplicons were sequenced with the use of a Big Dye Terminator v3.1 Cycle Sequencing kit (Applied Biosystems, Foster city, CA, USA) and a 3130 XL Genetic analyser (Applied Biosystems) according to the manufacturer's instructions. Sequencing primers were designed to be within the LAMP amplicon.
Determination of optimum temperature. To determine the optimum conditions, the temperature and concentration of PNA were examined. The reaction mix was incubated at 60-65°C for 30 min and Tp was monitored. Alternatively, PNA was added to the reaction mix at different concentrations of 5, 10, 15, 16.75, 20, and 40 pmol, respectively.
Statistical analysis. To examine the difference of Tp values, the Kruskal–Wallis H-test was performed. All experiments were performed five times. Differences were considered statistically significant at p-values of less than 0.05.
Results
LAMP and mutation-oriented LAMP reaction. We compared the conventional LAMP and mutation-oriented LAMP reaction for the KRAS gene. In the LAMP reaction, all examined samples exhibited a positive reaction for KRAS. The amplification of the gene was observed in 10-15 min. There was no difference in the Tp value of the KRAS wild-type and mutation groups. In contrast, the mutation-oriented LAMP reaction delayed gene amplification in the KRAS wild-type group (NCI-H1975, HSC-3, HT-29, and healthy control). In the LAMP reaction, the Tp value was not significantly different for the KRAS mutation (15’56) and wild-type groups (14’56). However, in the mutation-oriented LAMP reaction, the Tp value significantly differed for the KRAS mutation (18’34) and wild-type groups (35’42) (p<0.0001; Figure 2).

Loop-mediated isothermal amplification (LAMP) and the mutation-oriented LAMP reaction in cancer cell lines. The mutation-oriented LAMP reaction was suppressed in the HSC-3 cell line (oral squamous cell carcinoma), NCI-H1975 cell line (lung adenocarcinoma), and healthy oral mucosal cells, which have the wild-type allele for Kirsten rat sarcoma viral oncogene homolog (KRAS) (solid line). The LAMP reaction amplifies all KRAS alleles (broken line).
We measured the difference between the Tp value of mutation-oriented LAMP reaction and the LAMP reaction. The differences were: 1’00 in T84, −0’12 in NCI-H1573, 2’54 in MIA PaCa-2, 1’00 in SNU-1, 14’51 in HSC-3, 10’14 in NCI-H1975, 12’08 in HT-29, and 13’33 in healthy controls (Table III).
The DNA sequences of each amplicon were evaluated and were found to be compatible with the estimated sequences. Optimization of reaction conditions. To explore optimal conditions, the reaction temperature and lock primer concentration were examined. The time at which the upstroke of fluorescence level elevation was recorded, and the difference of Tp value between the KRAS wild-type and mutation groups was examined. To compare the difference of reaction temperature, reaction mixtures were incubated at 60-65°C. Differences of Tp value in the two groups at different reaction temperatures are presented in Table IV. At 65°C, the Tp value was faster and the difference between the wild and mutant types was the greatest (Table IV). To compare the difference of the concentrations of lock primer, we conducted a mutation-oriented LAMP reaction with different PNA concentrations. Differences of Tp value between the KRAS wild-type (healthy control) and KRAS mutation type (T84) are presented in Table V. There was little difference beyond the concentration of 16.75 pmol. Therefore, we conducted both LAMP and mutation-oriented LAMP reaction at 61°C with 16.75 pmol PNA for further experiments (Table V).

Detection limit for Kirsten rat sarcoma viral oncogene homolog (KRAS) mutation by loop-mediated isothermal amplification (LAMP) reaction (A) and mutation-oriented LAMP reaction (B). Both LAMP and mutation-oriented LAMP reactions amplify the KRAS gene at concentrations over 10 pg; however, the time to positive fluorescence (Tp) value was delayed at concentrations of 10 and 100 pg in the mutation-oriented LAMP reaction.
Differences between loop-mediated isothermal amplification (LAMP) and mutation-oriented LAMP reaction.
To examine the detection limit of the mutation-oriented LAMP reaction, a template gDNA of T-84 cells was added into the reaction mix at concentrations of 100 fg to 15 ng. Both mutation-oriented LAMP and conventional LAMP reactions were conducted, and the results showed that both reaction procedures amplified the mutation allele of the KRAS gene over 10 pg of template gDNA; however, the Tp value was delayed at concentrations of 10 and 100 pg in the mutation-oriented LAMP reaction (Figure 3). Therefore, we infer that the mutation-oriented LAMP reaction requires a template gDNA of mutation allele of over 1 ng.
Furthermore, we performed the assay using genomic DNA from T-84 cell line gradually diluted (100, 10, 1, 0.1, 0.01, 0.001 and 0%) into wild-type DNA from healthy controls. The reproducible sensitivity of mutation-oriented LAMP was 10%, detected in all five repeated examinations (100%). The 1% dilution was detected in 60% (Table VI).
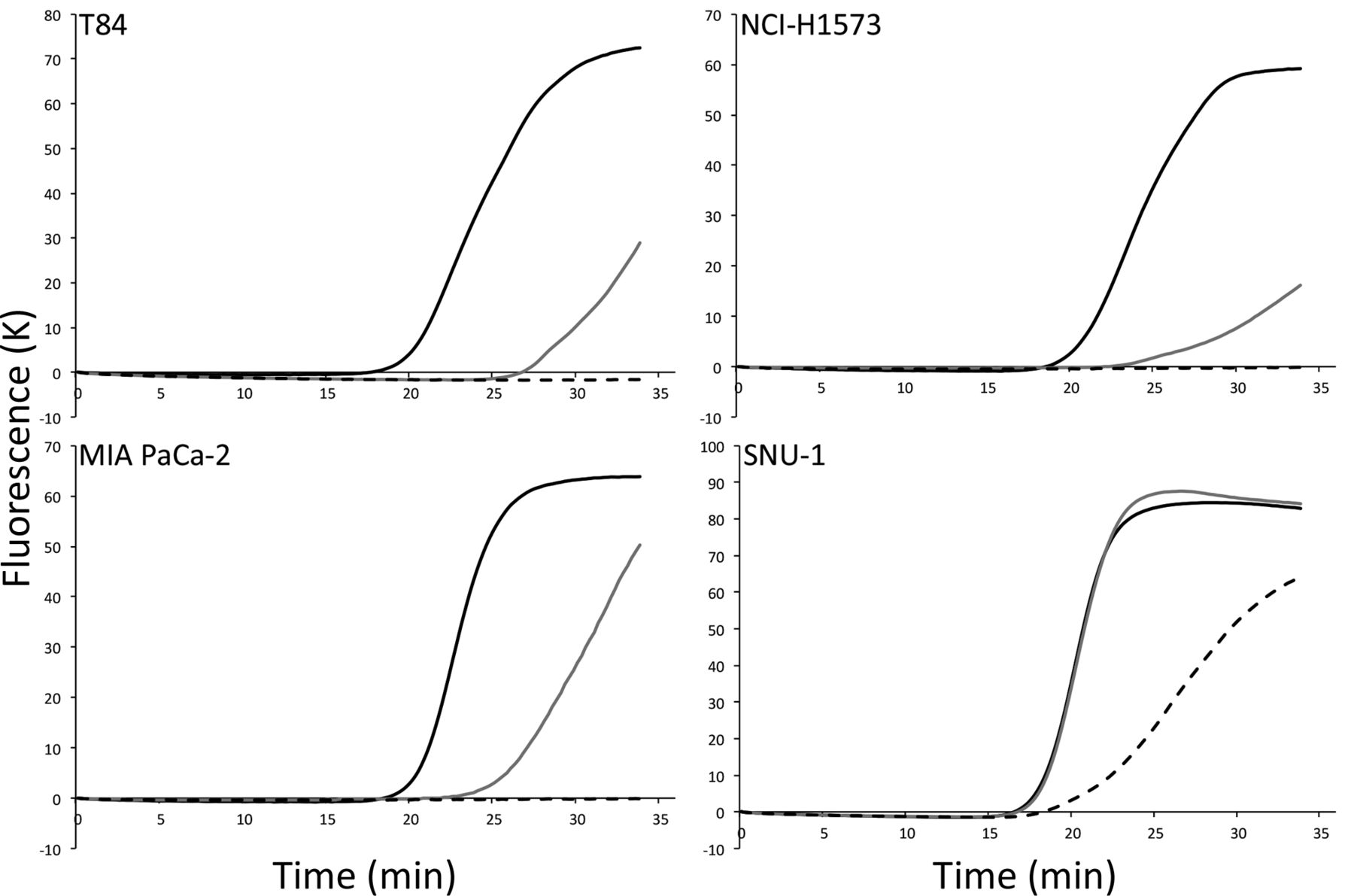
Mutation-oriented LAMP analysis directly from crude samples. We also investigated boiled cell lines as template. Each boiled cell works as the template for LAMP and mutation-oriented LAMP systems. We examined the detection limit of number of cells, and amplification was observed in 500 cells (T84: 16’34±0’11, NCI-H1573: 18’03±0’07, MIA PaCa-2: 17’48±0’19, SNU-1: 15’08±0’14) and 100 cells (T84: 25’07±0’16, NCI-H1573: 23’07±0’09, MIA PaCa-2: 22’39±0’18, SNU-1: 15’51±0’23); however, amplification could not be detected in samples of 10 cells, except for SNU-1 cells (16’48) (Figure 4).

Detection of Kirsten rat sarcoma viral oncogene homolog (KRAS) mutation using crude samples. Crude samples were obtained by washing and boiling several cultured cells in phosphate-buffered saline. Mutated KRAS codons 12 and 13 were detected from over 100 cells in the reaction mixture (500 cells; solid line, 100 cells; gray line). Moreover, mutant alleles could be detected from over 10 cells (broken line) in SNU-1 cell.
Discussion
The detection of the KRAS mutation at codons 12 and 13 is quite important for cancer therapeutics using antibodies against EGFR and EGFR tyrosine kinase inhibitor. We successfully developed a mutation-oriented LAMP assay using several cultured cell lines. This assay was able to amplify the DNA fragment of the mutated KRAS gene within 20 min. Lock primers made of PNA can inhibit gene amplification in KRAS wild-type codons 12 and 13. Thus, the assay is able to amplify all possible mutations in KRAS codons 12 and 13.
Various assays for KRAS mutation detection exist, including PCR-RFLP, PCR-SSCP, MASA, and other PCR-based technologies or DNA sequencing (20-24). PCR-RFLP requires DNA amplification, restriction enzyme digestion, and electrophoresis. PCR-SSCP requires DNA amplification, denaturation of amplicons, and polyacrylamide gel electrophoresis or capillary-gel electrophoresis. MASA requires DNA amplification with several PCR primer sets for each mutation as well as electrophoresis. All of these PCR-based methods are time consuming (shortest time, 3.5 h), require careful DNA extraction, and involve several steps (DNA extraction, PCR, electrophoresis, etc.) (32). DNA sequencing can detect unknown mutations; however, this is a time- and cost-ineffective method. In contrast, the mutation-oriented LAMP reaction enables identification of gene mutation within 20 minutes.
The mutation-oriented LAMP method enables targeting of all KRAS mutations in codons 12 and 13 at once. Jakovljevic et al. reported that 34.7% of patients with metastatic colorectal cancer have KRAS point mutations in codons 12 and 13. Among them, the frequencies of mutations in codon 12 were: p.G12D, 43.9%; p.G12V, 21.2%; p.G12A, 10.6%; p.G12C, 7.6%; p.G12S, 4.5%; p.G12R, 1.5%; the frequency of the only mutation in codon 13 was 10.6% for p.G13D (33). In the mutation-oriented LAMP method, the wild-type allele of KRAS was masked by lock primer, composed of PNA with a complementary sequence for KRAS codons 12 and 13. PNAs are DNA mimics, wherein the phosphoribose backbone is replaced by a peptide-like repeat of (2-aminoethyl)-glycine units (34). Due to this chemical difference, PNA differs from DNA molecules in several aspects. PNA–DNA hybrids have a higher thermal stability compared to the corresponding DNA–DNA hybrids. PNA–DNA hybrids are more easily destabilized by single base-pair mismatches than the corresponding DNA–DNA hybrids. Thus, PNAs cannot serve as primer molecules in PCR (31, 35). Therefore, if there are mutations in codons 12 and 13, the lock primer detaches from the template DNA, and the LAMP reaction is initiated.
Differences in mutation-oriented loop-mediated isothermal amplification (LAMP) reaction speed based on temperature.
Differences in mutation-oriented loop-mediated isothermal amplification (LAMP) reaction based on lock primer concentration.
Sensitivity of mutation-oriented loop-mediated isothermal amplification (LAMP) assay in mixed-cell populations.
Another advantage of this novel LAMP method was its detection ability in the crude sample. Conventional KRAS mutation detection methods require DNA extraction from samples. However, we were able to detect the KRAS mutations without performing DNA extraction. This was due to the high sensitivity of the LAMP method (26). In this study, we successfully detected the KRAS mutation from 100 boiled cells in T84 (c.38G>A), NCI-H-1573 (c.35G>C), and MIA PaCa-2 (c.34G>T) cell lines. Moreover, we detected mutations from as few as 10 cells in the SNU-1 (c.35G>A) cell line. These differences could be attributable to the differences of the substituted base and its position.
In summary, the mutation-oriented LAMP method detects KRAS gene mutation in codons 12 and 13 by using lock primers. As this analysis can be carried out without DNA extraction, this method could be applied for clinical examination in future applications.
Acknowledgements
This study was supported by Grant-in-Aid for Young Scientists (B) to Shotaro Mukae (No.24792011), and Grant-in-Aid for Scientific Research (C) to Naoyuki Matsumoto from the Ministry of Education, Cultures, Sports, Science and Technology, Japan.
- Received January 20, 2016.
- Revision received March 8, 2016.
- Accepted March 17, 2016.
- Copyright© 2016 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved





{kind=link}
{kind=link}
{kind=link}
{kind=link}